Cell motion as a stochastic process controlled by focal contacts dynamics
Texte intégral
Figure
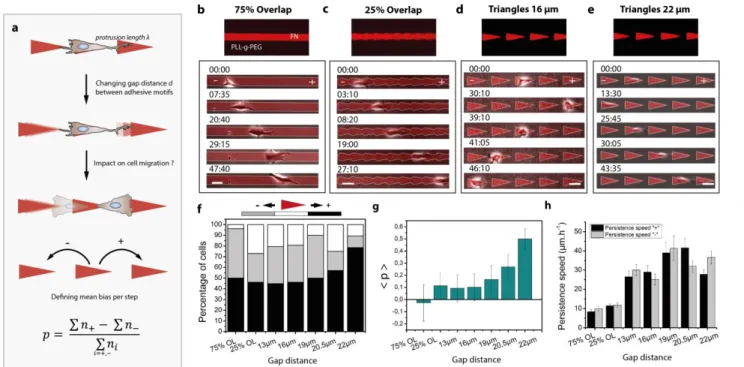
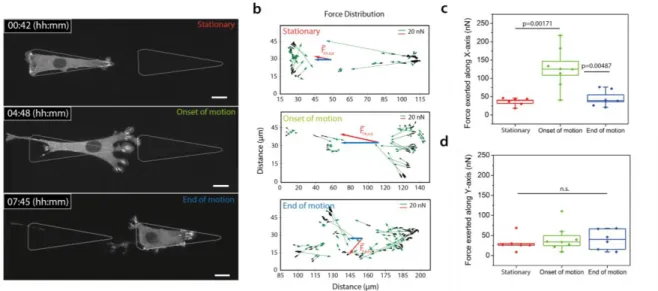
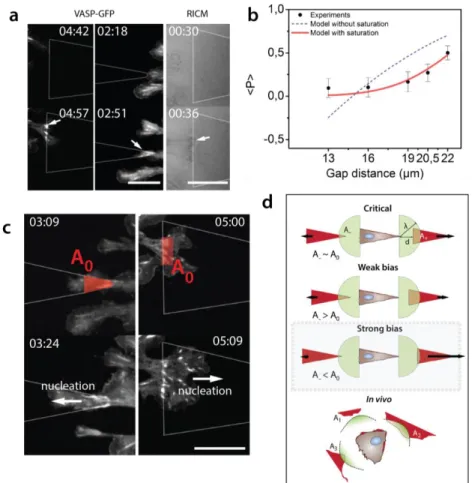
Documents relatifs
In vitro angiogenesis assays were thus performed using HMEC-1 cells treated with various concentrations of native MVL-PLA2 or with MVL-PLA2 catalytically inactivated by incubation
We first compared the amino acid sequence close to the Glu-374 or to the corresponding amino acid of various PKC isoforms (Fig. 2A) and detected putative structural motives very
À partir de combien de kilomètres parcourus le tarif proposé par l’agence EUROPAUTO est-il apparemment la plus avantageux. Dans la suite de l'activité, on note x le nombre
Du fait, un agent peut être considéré comme un objet doté de capacités supplémentaires : recherches de satisfactions (intentions, pulsions) d'une part, et d'autre part
This clearly demonstrated the lower capacity of paxillin ΔLIM2 and paxillin 1334 constructs to compete with the wild-type full-length paxillin for binding to endogenous β3 integrins
(c) Western blot of protein extracts from NIH-3T3 fibroblasts transiently co-expressing β3 WT _GFP and mCherry-tagged paxillin proteins, wt or deletion mutants.. The paxillin
Using kindlin-deficient cells, we show that kindlin membrane localization and diffusion are crucial for integrin activation, cell spreading and FAs formation.. Thus, kindlin uses
A: Western blot detection of HM-phosphorylated AKT2 (pAKT2 S474). Vinculin was used as loading control and Nb8 was detected through its V5-tag. AKT2 and pAKT2 S474 were detected
