Effect of agomelatine on memory deficits and hippocampal gene expression induced by chronic social defeat stress in mice
12
0
0
Texte intégral
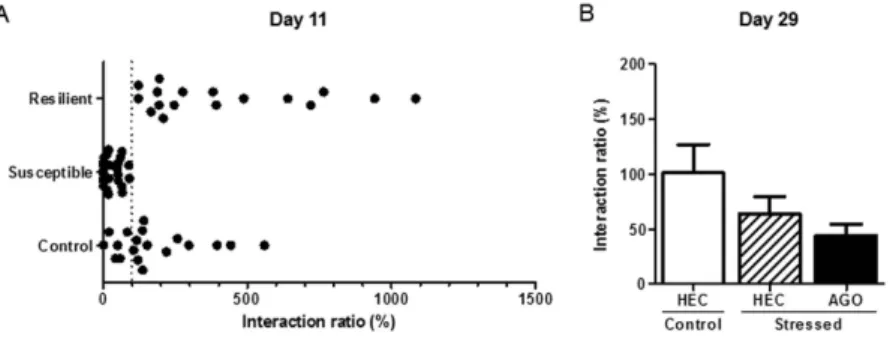
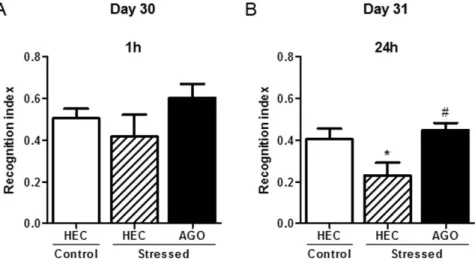
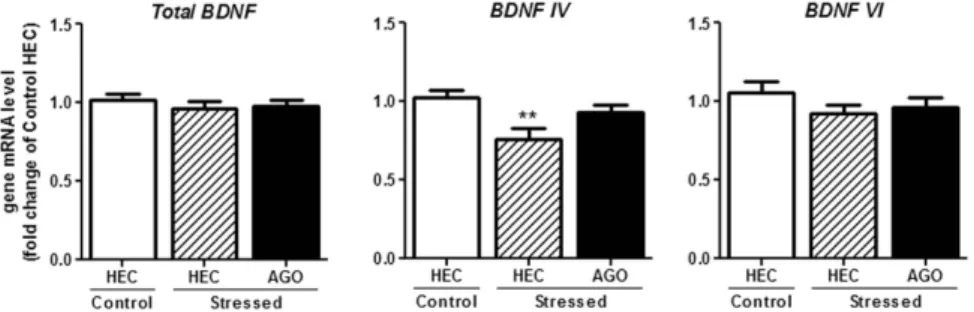
Figure

Documents relatifs
