X-ray microtomography (micro-CT): a reference technology for high-resolution quantification of xylem embolism in trees
Texte intégral
Figure
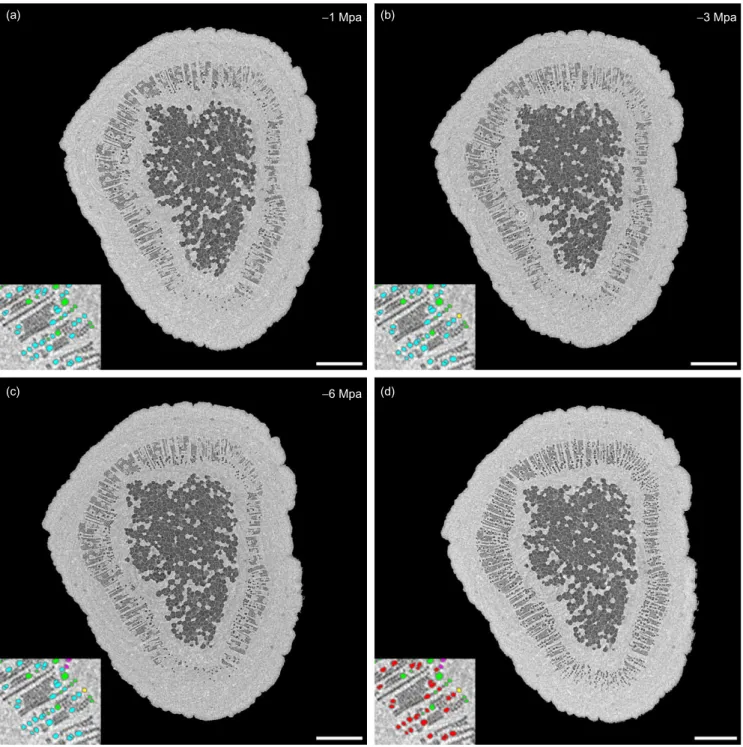
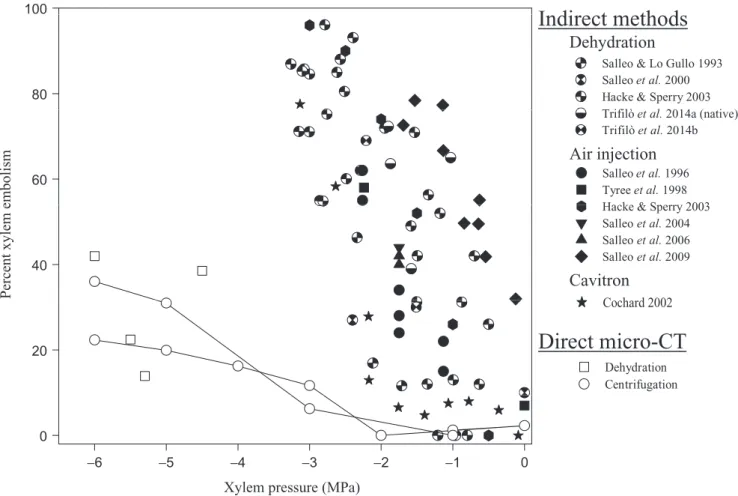
Documents relatifs
As we will see, Ife demonstrates a harmony policy that is thus for- mally identical to Finnish: non-contrastive vowels are fully transparent in medial positions, but when
This thesis presents the correction function method (CFM), a new family family of immersed methods designed to solve the incompressible Navier-Stokes equations,
(a) Total organic carbon content (wt % TOC), (b) total alkenone content ( mg per gram of sediment), (c) total alkenone content relative to TOC ( mg per gram of TOC), (d)
In this paper, we also give a characterization of the criticality eigenvalue associated to the energy- dependent diffusion model used in nuclear reactor theory [ 5 , 24 , 25 ]..
The combined impact of the second-hand market redistribution between regions and policy flows represents a small proportion of the remaining fleet size in 2009 in Nord-Pas
Selon les estimations des spécialistes, ce modèle agro-industriel tertiarisé (MAIT), caractérisé notamment par une consommation de masse de produits industrialisés
There are three main farms’ trajectories for organic farmers: the installation directly in organics, conversion of traditional low-intensity farms in the organics, and the
As parallel imaging moves forward to many-element coils with better spatial encoding abilities (e.g., (14)), future clinical protocol designs will have to fully
