Impact du longicorne noir, Monochamus scutellatus
scutellatus, sur l’épinette noire et le sapin baumier
à la suite de chablis en forêt boréale irrégulière
Mémoire
Mayerline Murillas Gómez
Maîtrise en sciences forestières
Maître ès sciences (M.Sc.)
Québec, Canada
iii
Résumé
Cette étude vise à déterminer quels sont les facteurs qui influencent le niveau d’attaque du longicorne noir (Monochamus scutellatus scutellatus) après un chablis en forêt boréale irrégulière, et quelle est la période de temps pendant laquelle les arbres affectés restent susceptibles aux attaques. Des mesures ont été prises dans un chablis naturel sur la Côte-Nord et dans un petit chablis artificiel réalisé à la Forêt Montmorency. Les espèces forestières à l’étude sont l'épinette noire (Picea mariana (Mill.) BSP) et le sapin baumier (Abies
balsamea (L.) P. Mill.). Les données récoltées indiquent que la sévérité du chablis et le nombre d’années
depuis la mort des arbres ont un effet significatif sur le nombre de trous d’entrée de larves des longicornes. De plus, l’essence forestière et l’état de l’arbre après le chablis influencent l’attractivité que les arbres peuvent exercer sur les longicornes noirs. Ces résultats pourraient aider les aménagistes forestiers à mieux préparer les opérations de récupération de bois de chablis en forêt boréale.
v
Abstract
This study aims to identify factors influencing the attack level by the whitespotted sawyer (Monochamus
scutellatus scutellatus) after windthrow in uneven-aged boreal forests, and the period of time during which
affected trees remain susceptible to attacks. Measures were taken in a natural windthrow on the Côte-Nord and in a small experimental windthrow generated at the Forêt Montmorency. Studied tree species are black spruce (Picea mariana (Mill.) BSP) and balsam fir (Abies balsamea (L.) P. Mill.). The collected data indicate that windthrow severity and the number of years since tree death significantly affect the number of entry holes of the whitespotted sawyer larvae. In addition, tree species and status after windthrow influence their attractiveness to the whitespotted sawyer. These results could help forest managers to improve salvage logging plans after windthrow in boreal forest.
vii
Table des matières
RÉSUMÉ ... III ABSTRACT ... V TABLE DES MATIÈRES ... VII LISTE DES TABLEAUX ... IX LISTE DES FIGURES ... XI LISTE DES FIGURES ... XI AVANT-PROPOS ... XIII REMERCIEMENTS ... XV INTRODUCTION GÉNÉRALE ... 1 RÉSUMÉ ... 7 ABSTRACT ... 8 1 INTRODUCTION ... 9
1.1 Facteurs influençant la présence de longicornes sur les arbres... 12
2 OBJECTIFS ET HYPOTHÈSES DE L’ÉTUDE ... 13
3 MATÉRIEL ET MÉTHODES ... 14
3.1 Chablis naturels sur la Côte-Nord ... 14
3.2 Chablis expérimental à la Forêt Montmorency ... 17
3.3 ANALYSES STATISTIQUES ... 18
viii
3.3.2 Forêt Montmorency ... 20
4 RÉSULTATS ... 20
4.1 Chablis naturel sur la Côte-Nord... 20
4.2 Chablis expérimental à la Forêt Montmorency ... 21
5 DISCUSSION ... 22
5.1 Côte-Nord ... 22
5.2 Forêt Montmorency ... 25
6 CONCLUSION... 26
ix
Liste des tableaux
TABLEAU 1.CARACTÉRISTIQUES DES SITES ÉCHANTILLONNÉS EN 2011 DANS LE CHABLIS NATUREL DE 2006 SUR LA
CÔTE-NORD.QUÉBEC,CANADA. ... 28 TABLEAU 2. RÉSUMÉ DES VARIABLES EXPLICATIVES OBTENUS DU MODÈLE DE RÉGRESSION DE POISSON
GÉNÉRALISÉE AVEC EXCÈS DE ZEROS AVEC LE PEUPLMENT COMME FACTEUR ALÉATOIRE, PRÉDISANT LA DENSITÉ DE TROUS D’ENTREÉE DE LONGICORNES DANS LES ARBRES D’AUN CHABLIS QUI S’EST PRODUIT EN
2006 SUR LA CÔTE-NORD AU QUÉBEC, EN FONCTION D’UN ENSEMBLE DE VARIABLES EXPLICATIVES À L’ÉCHELLE DE L’ARBRE AU DU PEUPLEMENT. ... 29 TABLEAU 3.RÉSUME DU MODÈLE DE RÉGRESSION DE POISSON GÉNÉRALISÉE PRÉDISANT LE NOMBRE D’ADULTES DE
M. S. SCUTELLATUSEN FONCTION D’UN ENSEMBLE DE DONNÉES EXPLICATIVES AU NIVEAU DE L’ARBRE DANS DES CHABLIS EXPÉRIMENTAUX RÉALISÉS À LA FORÊT MONTMORENCY. ... 30 TABLEAU 4. RÉSUMÉ DU MODÈLE DE RÉGRESSION MULTIPLE À DISTRIBUTION NÉGATIVE AVEC EXCÈS DE ZÉROS
PRÉDISANT LE NOMBRE DE LARVES DE M. S. SCUTELLATUSEN FONCTION D’UN ENSEMBLE DE DONNÉES EXPLICATIVES AU NIVEAU DE L’ARBRE DANS DES CHABLIS EXPÉRIMENTAUX RÉALISÉS À LA FORÊT
MONTMORENCY. ... 30 TABLEAU 5.POURCENTAGE D'HUMIDITÉ DES ARBRES EN 2011 SELON LEUR ÉTAT APRÈS LE CHABLIS NATUREL DE
2006 SUR LA CÔTE-NORD,QUÉBEC,CANADA. ... 30 TABLEAU 6. POURCENTAGE D'HUMIDITÉ DES ARBRES EN 2011 ET 2012 SELON LEUR ÉTAT APRÈS LES CHABLIS
xi
Liste des figures
FIGURE 1.RÉGION À L'ÉTUDE SUR LA CÔTE-NORD DU QUÉBEC,CANADA.CARTE ADAPTÉE DE CIMON-MORIN,2009.
... 32 FIGURE 2.RÉGION À L'ÉTUDE À LA FORÊT MONTMORENCY.QUÉBEC,CANADA.CARTE ADAPTE DE
WWW.FM.ULAVAL.CA ... 32 FIGURE 3.RELATION ENTRE LE POURCENTAGE DE PERTE DE VALEUR PAR BILLOT D'ÉPINETTE NOIRE ET SAPIN
BAUMIER ET LA DENSITÉ DE TROUS D'ENTRÉE DE LONGICORNES.ADAPTÉ DE SAFRANYIK ET RASKE (1970). .. 33 FIGURE 4.RELATION ENTRE LA SÉVÉRITÉ DU CHABLIS ET LE NOMBRE DE TROUS D'ENTRÉE DE LARVES DE
LONGICORNES DANS LA RÉGION DE LA CÔTE-NORD.QUÉBEC,CANADA. ... 34 FIGURE 5.DYNAMIQUE DE LA MORTALITÉ DES ARBRES APRÈS UN CHABLIS QUI S'EST PRODUIT EN 2006 DANS LA
RÉGION DE LA CÔTE-NORD.QUÉBEC-CANADA. ... 35 FIGURE 6.DYNAMIQUE DU NOMBRE DE TROUS D'ENTRÉE DE LARVES DE LONGICORNES EN FONCTION DE LA DATE DE
MORTALITÉ DES ARBRES AVANT ET APRÈS LE CHABLIS DANS LA RÉGION DE LA CÔTE-NORD.QUÉBEC-CANADA. ... 36 FIGURE 7. NOMBRE D'ADULTES DE M. S. SCUTELLATUS (MOYENNE ± ERREUR TYPE) SUR LES ARBRES SELON
L'ESSENCE FORESTIÈRE (A) ET SELON L'ÉTAT DES ARBRES (B) À LA SUITE DES CHABLIS SIMULÉS À LA FORÊT
MONTMORENCY EN 2011-2012. DES LETTRES DIFFÉRENTES INDIQUENT DES DIFFÉRENCES SIGNIFICATIVES SELON UN SEUIL Α=0.05. ... 37 FIGURE 8.NOMBRE LE LARVES DE M. S. SCUTELLATUS (MOYENNE ± ERREUR TYPE) SUR LES ARBRES SELON
L'ESSENCE FORESTIÈRE (A) ET SELON L'ÉTAT DES ARBRES (B) À LA SUITE D'UN CHABLIS SIMULÉ À LA FORÊT
MONTMORENCY EN 2011-2012.DES LETTRES DIFFÉRENTES INDIQUENT DES DIFFÉRENCES SIGNIFICATIVES SELON UN SEUIL Α=0.05. ... 38
xiii
Avant-propos
Ce projet de recherche a été conçu grâce à un partenariat entre la Chaire de recherche industrielle CRSNG - Université Laval en sylviculture et faune de l’Université Laval et Ressources naturelles Canada.
Ce mémoire de maîtrise comporte un chapitre principal qui contient la mise en contexte, des explications sur la façon dont les données ont été recueillies sur la Côte-Nord et à la Forêt Montmorency de l’Université Laval, l’analyse de ces données ainsi que leur interprétation. Ce projet n’aurait pu être possible sans la participation de mon directeur, Dr. David Pothier (Université Laval) et mon codirecteur, Dr. Christian Hébert (Ressources naturelles Canada).
xv
Remerciements
Je tiens à remercier plusieurs personnes liées à divers aspects de ce projet. Je tiens tout d’abord à remercier de tout cœur mon directeur de maîtrise, Dr. David Pothier, pour avoir eu confiance en moi, pour m’avoir donné l’opportunité de continuer ma formation académique et pour ses nombreux conseils et son amabilité. Je veux aussi remercier mon codirecteur, Dr. Christian Hébert, pour m’avoir poussé à relever des défis, pour sa disponibilité, pour son amabilité et ses conseils. Merci à M. Yves Dubuc du laboratoire d’écologie et de diversité des insectes forestiers, pour sa disponibilité, pour son aide au laboratoire et surtout pour son aide sur le terrain. Merci à Mme. Hélène Crépeau du Service de Consultation Statistique de l’Université Laval, pour sa précieuse aide avec les analyses statistiques de mon projet. Aussi j’aimerais remercier M. David Gervais du Centre de Foresterie de Laurentides pour sa précieuse aide avec les analyses dendrochronologiques.
Mille mercis à ma famille, en particulier à ma mère, mes tantes et mon grand-père, qui ont toujours été là pour moi, qui m’ont encouragé en tout temps et qui m’ont donné leur amour tout au long de mon cheminement. Je veux aussi remercier spécialement mon cousin Alejandro et son épouse Claudia, car sans eux, rien n'aurait été possible, merci pour avoir été là pour moi quand j’avais besoin de leurs mains. Merci aussi à Louis-Philippe pour son support et ses encouragements.
1
Introduction générale
Les perturbations naturelles sont des forces motrices générant des changements importants dans les écosystèmes (De Grandpré et al., 2000) et leur conférant une dynamique qui leur est propre (Bergeron, 2007). Elles modifient les attributs des peuplements produisant ainsi une variabilité dans les habitats (Vaillancourt, 2008). Toutes les perturbations naturelles peuvent être représentées à l’aide de trois descripteurs (Gauthier et
al., 2001; Vaillancourt, 2008), soit les intervalles, c'est-à-dire la fréquence et le temps de retour ou cycle; la
superficie, qui fait référence à la taille de l’événement et la sévérité, qui est la proportion des arbres affectés ou la spécificité de la perturbation (Gauthier et al., 2001; Vaillancourt, 2008).
Le chablis est une perturbation dite catastrophique qui exerce un rôle prépondérant dans la dynamique du paysage forestier de la forêt boréale du Québec (Doyon et Bouffard, 2008). Selon Helms (1998), un chablis correspond à un arbre ou à groupe d’arbres renversé ou cassé par le vent. Au Québec, il y a une fréquence plus élevée de chablis dans les domaines bioclimatiques dominés par les essences résineuses par rapport à ceux dominés par les feuillus (Vaillancourt, 2008). Les chablis sont des perturbations qui produisent des attributs différents de ceux des forêts aménagées et des forêts naturelles non perturbées. Ces attributs permettent à des assemblages d’espèces distincts de cohabiter (Vaillancourt, 2008) et de profiter du matériel disponible.
Les risques de chablis dépendent de plusieurs facteurs locaux et de l’interaction entre ces divers facteurs. La topographie, le climat, les propriétés du sol et les caractéristiques du peuplement sont les principaux facteurs environnementaux influençant la susceptibilité des forêts au chablis (Mitchell et al., 2001; Ruel et al., 2002, Ruel, 2010). Parmi ces facteurs, la topographie a beaucoup d’influence car elle est reliée à la pente, l’exposition du peuplement et aux variations locales qui modifient la direction des vents (Larouche, 2005). La vitesse et la direction des vents sont aussi des facteurs climatiques importants. Par exemple, les vents qui empruntent des directions inhabituelles causent plus de dommages, car les arbres y sont moins acclimatés (Moore, 1977; Robertson, 1987). De plus, la vitesse des vents détermine la force qui sera exercée sur les arbres (Ruel et Benoit, 1999).Il est difficile d’établir une vitesse moyenne à partir de laquelle le vent peut occasionner un chablis. Selon Ruel (2010), dans l’Ouest du Québec, une augmentation des dommages a été observée dans des peuplements d’épinette noire soumis à la coupe par bandes à la suite d’une tempête où les vents ont approché les 90 km/h.
2
Chaque espèce d’arbre a un niveau de résistance particulier, mais cette résistance spécifique demeure variable à cause de l’interaction fréquente avec d’autres facteurs, comme les conditions d’enracinement (Busby, 1965). Les caractéristiques du sol ont une influence sur la capacité d’enracinement des arbres
(Stokes et al., 1995), les sols profonds avec des textures fines (Coutts, 1983) et bien drainés (Hütte, 1968)
permettent aux arbres d’avoir un meilleur enracinement. La vulnérabilité aux chablis peut varier selon la composition du peuplement, car la susceptibilité augmente avec la hauteur de l’arbre (Cremer et al., 1982 ; Smith et al., 1987). De plus, la densité du peuplement (Gardiner et al., 1997; Bergeron, 2007) joue un rôle fondamental puisque les arbres qui ont crû isolés et exposés aux vents sont moins vulnérables que ceux qui ont crû à l’intérieur du peuplement et ayant peu été exposés aux vents (Cremer et al., 1982). De plus, selon les résultats de l’étude de Ruel et Benoît (1999) sur le chablis de 1994 dans les régions de la Gaspésie et de Charlevoix, le sapin baumier est une espèce plus sensible aux chablis que l’épinette noire à cause de sa plus grande susceptibilité aux caries de souche et de racines, qui affectent les arbres à partir de 50 ans (Whitney, 1989 ; Lavoie, 2011).La littérature nous permet de retracer plusieurs cas de chablis survenus au Québec. Dans le cadre d’une étude de Lehman et al. (1975) menée selon l’axe nord-sud entre la plaine du Saint-Laurent et la Baie James, 24 chablis d’une superficie moyenne de 115 ha et ayant été causés par des tornades ont été répertoriés sur une période de 10 ans. Aussi, d’autres types de tempête comme les orages violents peuvent causer des dommages aux forêts comme en 1984 lorsqu’un chablis a touché plusieurs stations en Outaouais pour une superficie totale de 1925 ha (MRN 1985). Ruel et Benoit (1999) mentionnent qu’en 1994, des chablis ont sévèrement affecté les régions de Charlevoix (2485 ha) et de la Gaspésie deux superficies (3100 ha). De plus, un chablis réparti sur environ 20000 ha de forêt a eu lieu entre la frontière de l’Ontario et la Mauricie en juillet 2006 (MRNF 2007).
Selon Bouget et Duelli (2004), les chablis font partie des perturbations naturelles qui jouent un rôle actif dans le cycle évolutif des forêts. Les chablis sont moins importants que les incendies forestiers en forêt boréale, mais le sont plus en forêt tempérée (Bouget et Duelli, 2004; De Grandpré et al., 2000; Vaillancourt, 2008). Par conséquent, aux endroits où la fréquence des feux est faible, le vent peut constituer la perturbation dominante et jouer un rôle majeur dans la dynamique forestière (DeGayner et al., 2005). Les chablis ont des impacts économiques importants dans les forêts aménagées (Gandhi et al., 2007). Par exemple, en 1995 dans l’état du Minnesota aux États-Unis, 20 % du bois d’un chablis n’a pu être récupéré (Minnesota Department of Natural Resources, 1996). On s’attend à ce que la taille des zones perturbées augmente avec les orages
3
violents et les tempêtes de vents de plus en plus fréquents associés aux changements climatiques. Ainsi, on anticipe des pertes économiques encore plus grandes (Lavoie, 2011). Toutefois, les perturbations de plus petite taille sont aussi très importantes, notamment pour les habitats qu’elles procurent à la faune, car elles contribuent substantiellement à maintenir une hétérogénéité élevée des habitats entre les perturbations de plus grande envergure (Gandhi et al., 2007).
Les chablis modifient considérablement l’environnement forestier. Les conditions post-chablis fournissent des attributs clés aux endroits affectés et jouent un rôle important dans la dynamique des populations animales et végétales (Vaillancourt, 2008). À la suite d’un chablis, les arbres peuvent être déracinés complètement ou partiellement (et donc appuyés sur d’autres arbres) ou leur tronc peut être cassé. Tous les arbres affectés deviennent des substrats attractifs pour les insectes consommateurs de matière ligneuse qui colonisent des arbres moribonds et stressés. Quand il y a une grande perturbation, les insectes ont la possibilité d’augmenter leur population pendant une certaine période de temps puis de coloniser des aires adjacentes, qu’elles aient été ou non affectées par la perturbation (Gandhi et al., 2007). Les attributs clés peuvent varier en fonction de la sévérité et de la récurrence des événements dans le même territoire et la biodiversité varie en fonction de la qualité de l’habitat et de la disponibilité des ressources (Vaillancourt, 2008).
Plusieurs groupes d’insectes peuvent être avantagés par les nouvelles conditions et niches écologiques qui sont engendrées par l’ouverture du couvert forestier ou par le déracinement des arbres à la suite d’un chablis (Vaillancourt, 2008). C’est le cas du longicorne noir (Monochamus scutellatus scutellatus (Say)). Cet insecte est originaire d’Amérique du Nord et est présent là où la forêt est composée de résineux (Peddle, 2000). Cet insecte cause des dommages économiques directs (Rose, 1957; Raske, 1972; Gardiner, 1975), car ses larves se retrouvent sur les arbres morts, moribonds ou affaiblis (Peddle, 2000) et se nourrissent de leurs tissus sous-corticaux (Hanks, 1999) en creusant des galeries profondes dans le bois. Ces galeries réduisent la valeur économique des arbres à un point tel qu’il ne vaut même plus la peine de les récolter (Gardiner, 1975). Cette problématique a incité la rédaction du présent document dans le but de trouver des réponses aux questions de l’industrie forestière en ce qui concerne les facteurs qui influencent les niveaux d’attaque par les longicornes noirs (Monochamus scutellatus scutellatus) après un chablis et la période de temps pendant laquelle les arbres affectés par un chablis sont plus susceptibles d’être attaqués par les longicornes noirs.
5
Chapitre principal
Impact du longicorne noir, Monochamus
scutellatus scutellatus, sur l’épinette noire et le
sapin baumier à la suite de chablis en forêt boréale
irrégulière
Mayerline Murillas Gómez
1, David Pothier
1Christian Hébert
2
1 Département des sciences du bois et de la forêt, Pavillon Abitibi-Price, 2405 rue de la Terrasse, Université Laval, Québec, Québec, G1V 0A6, Canada
2 Ressources naturelles Canada, Service canadien des forêts, Centre de foresterie des Laurentides, 1055 du P.E.P.S., P.O. Box 10380, Stn. Sainte-Foy, Québec, Québec, G1V 4C7, Canada
7
Résumé
Le longicorne noir, Monochamus scutellatus scutellatus, est un insecte qui profite des perturbations naturelles pour attaquer des arbres affaiblis ou récemment morts. On sait déjà que les galeries qu’il creuse dans le bois constituent un problème majeur pouvant limiter la rentabilité de la récupération post-feu. Les dommages qu’il cause après un chablis sont cependant peu connus. Ce travail de recherche vise à répondre à deux questions de l’industrie forestière : 1) quels sont les facteurs qui influencent les niveaux d’attaque par le longicorne noir après un chablis? et 2) quelle est la période de temps pendant laquelle les arbres affectés par un chablis sont plus susceptibles d’être attaqués par le longicorne noir en forêt boréale irrégulière? Pour répondre à ces questions, des mesures ont été effectuées sur la Côte-Nord et à la Forêt Montmorency au nord de Québec. Les espèces forestières à l’étude sont l'épinette noire (Picea mariana (Mill.) BSP) et le sapin baumier (Abies
balsamea (L.) P. Mill.). Le plan d’échantillonnage pour la Côte-Nord comporte 16 stations, dont 8 couvertes
par des peuplements purs et 8 par des peuplements mixtes. On a échantillonné des arbres dans un grand chablis ayant affecté 166 000 hectares sur la Côte-Nord en 2006 et mesuré le nombre de trous d’entrée de longicorne noir. Par ailleurs, le dispositif expérimental de la Forêt Montmorency a été réalisé en simulant un chablis à l’aide de machinerie forestière à l’automne 2011 et au printemps 2012. Ces opérations ont permis d'obtenir des arbres qui présentent des caractéristiques semblables à celles d’un chablis naturel: des arbres renversés (déracinés), cassés (coupés), moribonds (annelés) et vivants (témoins).
Les données récoltées sur la Côte-Nord indiquent que la probabilité de trouver des trous d’entrée de larves de longicorne augmente avec la sévérité du chablis et diminue avec le nombre d’années après la mort des arbres. Le type de peuplement n’a pas eu d’effet sur le nombre de trous d’entrée de larves de longicornes. En ce qui concerne les données récoltées à la Forêt Montmorency, elles indiquent que les femelles de M. s.
scutellatus préfèrent les épinettes noires aux sapins baumiers pour pondre leurs œufs. De plus, les adultes de M. s. scutellatus, préfèrent les arbres déracinés et coupés aux arbres annelés et témoins. Ces résultats
pourraient aider les aménagistes forestiers à mieux préparer les opérations de récupération de bois de chablis en forêt boréale.
8
Abstract
The whitespotted sawyer (Monochamus scutellatus scutellatus) is an insect that takes advantage of natural disturbances to attack weakened or recently dead trees. We already know that the galleries dug into the wood are a major issue that may limit the profitability of post-fire harvesting. The damage caused after a windthrow, however, are poorly known. This study aims to answer two questions of the forest industry: 1) what are the factors that influence the attack levels by the whitespotted sawyer after a windthrow? and 2) what is the period of time during which the trees affected by windthrow are more likely to be attacked by the whitespotted sawyer in irregular boreal forests? To answer these questions, measurements were taken on the Côte-Nord and at the Forêt Montmorency. The studied tree species are black spruce (Picea mariana (Mill.) BSP) and balsam fir (Abies balsamea (L.) P. Mill.). The sampling plan for the Côte-Nord includes 16 stations, of which 8 are covered by pure stands and 8 by mixed conifer stands. We sampled trees affected by a large windthrow affecting 166,000 hectares on the Côte-Nord in 2006 and measured the number of whitespotted sawyer entry holes. Furthermore, the experimental design of the Forêt Montmorency was conducted by simulating a windthrow with forest machinery in autumn 2011 and spring 2012. These operations resulted in trees that showed similar characteristics to those of a natural windthrow: fallen trees (uprooted), broken (cut), dying (ringed) and living (controls).
The data collected on the North Shore indicate that the probability of finding whitespotted sawyer larvae entry holes increases with the severity of the windthrow and decreases with the number of years after tree death. The forest stand composition has no effect on the number of whitespotted sawyer larvae entry holes. Regarding the data collected at the Forêt Montmorency, the results indicate that M. s. scutellatus females prefer black spruce to balsam fir to lay their eggs. In addition, M. s. scutellatus adults prefer uprooted and cut trees to ringed and control trees. Those results could help forest managers to improve salvage logging plans after windthrow in boreal forest.
9
1 Introduction
La forêt boréale représente 77 % du couvert forestier au Canada, ce qui en fait le biome forestier le plus important de ce pays (Cadieux, 2010). La forêt boréale du Québec comporte deux domaines bioclimatiques qui sont la sapinière à bouleau blanc et la pessière à mousses (Lavoie, 2011). Le gradient longitudinal de précipitation permet de distinguer les sous-domaines de l’ouest et de l’est (Saucier et al., 1998). Dans la pessière à mousses de l’ouest du Québec, la matrice forestière naturelle est surtout composée de jeunes forêts de structure équienne issues de feux.Dans les sous-domaines bioclimatiques de l’est, les précipitations sont influencées par le climat maritime (Saucier et al., 1998), ce qui allonge le cycle de feu et donne davantage d’importance aux perturbations secondaires comme les épidémies d’insectes et les chablis pour moduler la dynamique forestière (Bergeron et al., 2001). Les chablis sont des perturbations naturelles occasionnées par des vents violents et prédominent dans les forêts nordiques tempérées de l’Amérique du Nord (Doyon et Bouffard, 2009). Ils constituent des perturbations forestières qui produisent des changements d’habitat, notamment en introduisant une dynamique par trouées lorsqu’ils n’affectent que quelques arbres à la fois dans les forêts. Comme toutes les perturbations, on peut caractériser les chablis selon leur fréquence, leur taille et leur sévérité (Vaillancourt, 2008; Gauthier et al., 2001).
Les chablis entrainent la mortalité des arbres et leur impact sur les écosystèmes forestiers varie selon l’échelle spatiale considérée (Bergeron, 2007). Les chablis peuvent être classés en chablis catastrophiques ou endémiques. Les chablis catastrophiques sont ceux qui sont occasionnés par une forte tempête tandis que les chablis endémiques sont le résultat de plusieurs chablis dans le même territoire au fil des années (Rollinson, 1987; Ruel, 2010). Dans la forêt vierge, les petites perturbations causées par le chablis endémique sont très nombreuses (Larouche, 2005). Les chablis sont des perturbations qui occasionnent des dégâts tant environnementaux qu’économiques en Amérique du Nord et ailleurs dans le monde (Savill 1983 ; Lohmander et Helles, 1987; Mitchell, 1995a). Par exemple, Rollinson (1987) mentionne que le volume de bois endommagé à cause d’un chablis catastrophique dans une plantation commerciale de conifères en Grande-Bretagne a été évalué à environ 6 % de la récolte totale sur une période de 30 ans. Par ailleurs, au Minnesota, les pertes causées par le chablis endémique ont été évaluées à environ 20 % de la récolte totale au cours de la même période (Minnesota Department of Natural Resources 1996).
10
Les risques de chablis sont régis par l’interaction simultanée de plusieurs facteurs, comme la topographie, le climat, les caractéristiques du sol et du peuplement ainsi que l’aménagement (Ruel, 2000; Mitchell et al., 2001; Ruel et al., 2002). Toutefois, parmi ces facteurs, la topographie est celui qui a le plus d’influence sur la susceptibilité d’un peuplement aux chablis (Larouche, 2005). La vitesse du vent est importante car elle détermine la force à laquelle les arbres sont soumis (Ruel et Benoit, 1999). La direction des vents est aussi importante puisque les arbres auront tendance à être plus susceptibles au renversement lorsque les vents empruntent des voies inhabituelles. En effet, l’enracinement des arbres témoigne généralement de leur acclimatation aux vents dominants (Larouche, 2005). La susceptibilité au chablis dépend aussi des caractéristiques de l’espèce d’arbre, comme la profondeur de l’enracinement ou la rigidité de la tige, ainsi que des caractéristiques du peuplement comme la densité, la hauteur et l’âge (Fraser, 1962; Coutts, 1986). Ainsi, le sapin baumier est plus vulnérable aux chablis à cause de sa plus grande susceptibilité aux caries de souche et de racines, qui affectent les arbres surtout à partir de 50 ans (Whitney, 1989; Lavoie, 2011). Les pratiques sylvicoles telles que les coupes partielles et les coupes totales (Rollinson, 1987) peuvent aussi influencer les pertes par le chablis en modifiant les caractéristiques du peuplement aménagé, ce qui peut augmenter l’exposition des arbres aux vents (Ruel 1995; Ruel et al., 2003). Quel que soit le principal élément responsable du chablis, cette perturbation modifie substantiellement l’environnement forestier (Vaillancourt, 2008). Les conséquences d’un chablis peuvent varier selon sa sévérité, sa taille et sa récurrence sur la même station (Vaillancourt, 2008). À la suite d’un chablis, la mortalité de la plupart des arbres est causée par le bris des tiges et le déracinement des arbres qui peut être partiel ou complet (Vaillancourt, 2008). Donc, les arbres ne meurent pas toujours immédiatement, ce qui ajoute une dimension temporelle aux effets du chablis.
Les chablis jouent un rôle très important en augmentant la biodiversité des écosystèmes forestiers (Ulanova, 2000). Ils n’ont pas seulement une influence sur la végétation, mais aussi sur les populations animales, notamment les insectes. Les arbres renversés constituent des habitats hautement favorables au développement des insectes saproxyliques qui sont très nombreux en forêt boréale (Wikars et al., 2005). Ces insectes dépendent du bois mort ou moribond pendant une partie de leur vie ou pendant l’ensemble de leur cycle biologique (Speight, 1989). Parmi ces espèces saproxyliques, plusieurs sont xylophages, c'est-à-dire qu’elles se nourrissent des tissus ligneux (Saint Germain et al., 2007) comme dans le cas des longicornes (Hanks, 1999). Ces derniers peuvent devenir un problème pour l’approvisionnement en bois des usines de sciage. En effet, ils peuvent endommager la matière ligneuse à un point tel qu’il n’est souvent plus rentable de récolter le bois après une perturbation naturelle comme le feu (Bélanger et al., 2013). Toutefois, en ce qui concerne le chablis, on manque présentement d’information sur la dynamique des insectes xylophages
11 (Bouget et Duelli, 2004 ; Gandhi et al., 2007) et sur la récupération des arbres (Foster et Orwig, 2006 ; Doyon et Bouffard, 2008) pour tirer le même type de conclusion.
De nombreuses espèces saproxyliques montrent une spécificité pour un genre d’arbres ou pour l’état de décomposition du bois (Toivanen et Kotiaho, 2010). Dans la forêt boréale québécoise, le longicorne noir (Monochamus scutellatus scutellatus) est l’insecte xylophage qui a le plus grand impact économique après une perturbation naturelle (Morley 1939 ; Peddle, 2000). Ses larves creusent des galeries longues et profondes qui peuvent atteindre 9 cm dans le bois d’arbres morts ou moribonds (Gardiner, 1975 ; Peddle, 2000). Cette espèce peut occasionner des pertes allant jusqu’à 30 % de la valeur du bois (Safranyik et Raske, 1970). Les larves de M. s. scutellatus se nourrissent des tissus sous-corticaux pendant les premières semaines de leur développement, puis elles entrent dans l’aubier pour finalement revenir vers la zone cambiale pour former une case pupale (Hanks, 1999). De plus, la formation de galeries dans le bois peut occasionner l’entrée de champignons et la colonisation par d’autres insectes xylophages qui accélèrent la décomposition du bois (Peddle, 2000). Les adultes se nourrissent sur les rameaux, mais ils ne font pas de dégâts importants sur le bois (Peddle, 2000). Le cycle biologique de cet insecte prend environ 23 mois pour s’accomplir (Rose, 1957). Les adultes sont très actifs pendants les journées ensoleillées, mais les femelles préfèrent les journées plus fraîches. La période d’oviposition a lieu pendant les mois de juin et de juillet. Cet insecte se maintien au stade d’œufs pendant une période de 9 à 14 jours. Après l’éclosion, les larves commencent à creuser des tunnels à travers le phloème jusqu’au cambium. Cette étape prend environ 2 ou 3 semaines (jusqu’à la mi-août). Après, les larves passent à leur deuxième stade pendant lequel elles se nourrissent du cambium en approfondissant les tunnels. Cette étape peut prendre, elle aussi, entre 2 ou 3 semaines. Le troisième stade a lieu au début du mois de septembre. À ce stade, elles creusent des tunnels très profonds dans le bois. Les larves de troisième stade restent dans le bois pendant tout l’hiver et au printemps, elles commencent à passer à leur quatrième stade jusqu’à mi-août. À la mi-septembre, les larves commencent leur pré-nymphose jusqu’au prochain printemps. Par la suite, pendant deux semaines, le stade de nymphose a lieu. Entre la mi-mai et le début du mois de juin de la deuxième année, les adultes commencent à émerger (Rose1957; Raske, 1972).
Cette espèce peut se développer sur différentes espèces de conifère, comme le pin gris (Pinus banksiana Lamber),, le pin rouge (Pinus resinosa Aitom) et le pin blanc (Pinus strobus L.) (Saint-Germain et al., 2004). De plus, certaines observations indiquent que le longicorne noir attaquerait plus sévèrement l’épinette noire (Picea mariana (Mill.) BSP) que le sapin baumier (Abies balsamea (L.) P. Mill.) après un chablis (Boucher et
12
Hébert, 2010). Pour accomplir son cycle vital, cet insecte a besoin d’arbres fortement stressés car sa reproduction est seulement possible sur les arbres qui viennent d’être blessés et qui sont en train de mourir, possiblement parce que le bois est encore humide et les nutriments sont convenables (Raske, 1972; Vallentgoed, 1991; Saint-Germain et al,. 2004).
À la suite d’un chablis, on doit souvent adopter des mesures spéciales qui incluent des plans de récolte ponctuelle visant à limiter les pertes de bois dues aux attaques d'insectes xylophages (Dajoz, 1998). Les chablis ont des impacts négatifs sur l’aménagement forestier car ils désorganisent la planification des réseaux routiers et de la récolte. De plus, la récupération des arbres renversés n’est possible que durant une courte période de temps à cause des dommages des insectes xylophages (Mitchell 1995a, 1995b) et cette pratique devient plus coûteuse à cause de la valeur moindre des tiges affectées (Savill 1983; Mitchell 1995b; Quine 1995) et de la baisse de productivité des opérateurs de machinerie.
1.1 Facteurs influençant la présence de longicornes sur les arbres
Selon Bouget et Duelli (2004), l’espèce végétale, le stade de décomposition du bois, le diamètre de la tige, la position (debout ou couché), l’humidité des tissus ligneux et l’association à des micro-habitats influencent la présence d’insectes saproxyliques dont font partie les longicornes. Après une perturbation comme le chablis, le bois des arbres affectés subit des changements chimiques et physiques (p. ex : humidité) qui le rendent de moins en moins convenable pour un insecte comme le longicorne qui s’alimente d’abord de phloème (Hanks, 1999). Pour les longicornes, la qualité nutritionnelle du bois diminue rapidement suivant la mort de l’arbre (Hanks, 1999), ce qui réduit considérablement la fenêtre de temps durant laquelle cette ressource leur est disponible (Vaillancourt, 2008).De plus, les espèces composant le peuplement affecté par le chablis ont certaines caractéristiques (comme l’épaisseur de l’écorce, la quantité de résine, les mécanismes de défense, la densité du bois, etc.) qui influencent l’abondance d’insectes xylophages (Gardiner, 1975). Quand les arbres sont blessés à la suite d’un chablis ou d’une autre perturbation, ils libèrent des composés chimiques comme la térébenthine (Kula et Zabecki, 2006). Cette dernière est un mélange de monoterpènes que l’on retrouve dans la famille des Pinaceae (Dyer et Seabrook, 1978). D’ailleurs, on a déjà observé que plus un chablis est sévère, plus il y a de
13 possibilités d’observer une augmentation de population de Monochamus sp. (Kula et Zabecki, 2006), car les peuplements les plus affectés par un chablis produisent une plus grande quantité de composés volatils qui attirent les insectes xylophages (Austarå et al., 1986; Lindelöw et Risberg, 1992; Franklin et al., 2000). Les adultes de M. s. scutellatus sont attirés par des composés secondaires comme la térébenthine, notamment les femelles qui l’utilisent pour trouver les arbres hôtes propices à la survie de leur progéniture (Gardiner, 1975; Peddle, 2000). Les essences forestières qui ont des niveaux plus élevés d’alpha-pinène sont aussi les plus attirantes pour les longicornes noirs (Peddle, 2000).
Le diamètre des arbres renversés a aussi une influence sur la préférence des longicornes car plus le diamètre est grand, plus il y a de ressources pour se nourrir (Kula et Zabecki, 2006). Selon Ulyshen et Hanula, (2009) l’état des arbres après un chablis peut aussi avoir une influence sur la colonisation des insectes qui est influencée par la dynamique de dégradation des arbres (Vaillancourt, 2008). Les arbres qui sont sur le sol se dégradent plus rapidement que ceux qui sont appuyés sur d’autres arbres et ne touchent pas le sol directement (Ulyshen et Hanula, 2009). Ces arbres couchés sur le sol attirent des cérambycidés comme les
Asemum (Speight, 1989). Par ailleurs, la partie exposée au soleil d’arbres partiellement déracinés peut attirer
des insectes, mais les arbres qui ont des parties cachés du soleil ou complètement à l’ombre sont plus convenables pour certains autres insectes comme des cérambycidés, particulièrement les Monochamus (Speight, 1989).
2 Objectifs et hypothèses de l’étude
Le but principal de ce projet est de répondre à deux questions importantes pour l’industrie forestière 1) quels sont les facteurs qui influencent les niveaux d’attaque par les longicornes noirs (Monochamus scutellatus
scutellatus) après un chablis ? et 2) quelle est la période de temps pendent laquelle les arbres affectés par un
chablis sont plus susceptibles d’être attaqués par les longicornes noirs? Trois hypothèses de recherche ont servi de guide à l’étude :
H1. Les taux d’attaque du longicorne noir vont varier en fonction des caractéristiques des arbres renversés et des peuplements affectés.
14
P. 1 Le taux d’attaque du longicorne augmentera avec la sévérité du chablis. P. 2 Le taux d’attaque du longicorne sera plus élevé sur les gros arbres.
P. 3 Le taux d’attaque du longicorne sera plus élevé sur l’épinette noire que sur le sapin baumier.
P3.1 Parce que la fréquence des visites sur l’épinette noire sera plus élevée que sur le sapin baumier. P3.2 Parce que le taux d’acceptation de l’épinette noire comme hôte sera plus élevé que pour le sapin
baumier.
H2. Le taux d’attaque du longicorne noir devrait varier selon l’état des arbres et être plus élevé sur ceux demeurant convenables longtemps qui sont, par ordre décroissant, les arbres appuyés, les arbres déracinés, les arbres cassés et les arbres sur pied non affectés.
H3. Les arbres sont plus attaqués par le longicorne noir pendant les premières années qui suivent le chablis.
3 Matériel et méthodes
3.1 Chablis naturels sur la Côte-Nord
La première partie de l’étude comportait un plan d’échantillonnage sur le terrain afin d’évaluer la gravité des dommages causés par le longicorne noir à la suite des grands chablis qui ont affecté la Côte-Nord à la fin de l’été 2006. Cette région, située dans le nord-est du Québec, fait partie du sous-domaine bioclimatique de la pessière à mousses de l’est. Ce sous-domaine bioclimatique est principalement couvert de peuplements de conifères dominés par l’épinette noire (Picea mariana (Mill.) B.S.P.), mais on y retrouve aussi du sapin baumier(Abies balsamea (L.) Mill.), du bouleau à papier (Betula papyrifera Marsh.) et du peuplier faux-tremble (Populus tremuloides Michx.) (Bergeron, 1996; Saucier et al., 1998; Bergeron, 2007; De Grandpré et al., 2000). Dans ce sous-domaine, les peuplements à structure irrégulière représentent environ 70 % du territoire forestier (Gauthier et De Grandpré, 2003). Le climat y est froid et maritime avec une température moyenne
15 annuelle variant entre -2,5 et 0,0 ºC et des précipitations annuelles variant entre 1100 et 1300 mm, dont 35 % sous forme de neige. Le cycle de retour des feux de forêt est relativement long avec un intervalle moyen entre deux feux de plus de 350 ans (Bouchard et al., 2008). La topographie varie de vallonnée à accidentée, où l’altitude peut varier entre 450et 650 m (Légaré, 2010).
Deux secteurs situés au nord de la ville de Baie-Comeau ont été échantillonnés. Le premier secteur se trouvait dans la région du camp Saint-Pierre où opère la compagnie Produits forestiers Résolu (longitude : -68 º 23' 41"; latitude : 50º 9' 32") et le deuxième secteur était situé dans la région du camp Brooch où opère la compagnie Produits forestiers Arbec (50º 43' 5" N, 67º 48' 22" O) (Figure 1). Puisque plusieurs études soutiennent que l’état des arbres après une perturbation a une influence sur l’abondance des insectes xylophages (Jonsell et Weslien, 2003, Gibb et al., 2006, McGeoch et al., 2007; Ulyshen et Hanula, 2009), il a été jugé pertinent de comparer quatre catégories d’arbres qui varient en fonction de leur état à la suite d’un chablis : APP (arbres appuyés sur d’autres arbres, vivants et moribonds), DER (arbres déracinés, vivants et moribonds), CAS (arbres cassés, morts), TEM (arbres vivants sur pied ou témoin). L’échantillonnage a été effectué dans 16 stations distantes d’au moins 10 km au cours de l’été 2011. Nous avons comparé des peuplements purs (4 à dominance de sapin baumier et 4 à dominance d’épinette noire) et des peuplements mixtes (8 peuplements composés de sapin baumier et d’épinette noire). Une parcelle de 20 x 20 m a été établie dans chaque station et nous avons identifié tous les arbres de dimensions commerciales (DHP > 9,0 cm) et catégorisé leur état (mort, moribond, vivant) ainsi que leur position (cassé, déraciné, penché, droit). La surface terrière (ST) des différents types d’arbres a été compilée pour évaluer la sévérité du chablis qui a été déterminée par le rapport entre la ST des arbres affectés (états cassé, déraciné et penché) sur la ST totale (arbres affectés et non affectés) (Tableau 1).
Dans chaque station, on a choisi 10 arbres (5 sapins et 5 épinettes dans les peuplements mélangés) dont le DHP était d’au moins de 16 cm. Pour chacun des arbres, nous avons noté l’espèce, le statut (mort, moribond ou vivant), l’état (déraciné, appuyé, cassé ou droit) et le DHP. On a ensuite abattu chacun des arbres sélectionnés à l’aide d’une scie mécanique pour y prélever une section de 1,50 m de long à partir des premiers 50 cm du sol. Deux rondelles d’environ 3 cm d’épaisseur ont été prélevées, la première à environ 50 cm du sol et l’autre juste au-dessus de la bille de 1,50 m. Les rondelles ont été placées immédiatement dans des sacs de plastique à fermeture étanche et déposées dans une glacière. La première rondelle (50 cm du sol) a servi à l’analyse dendrochronologique pour déterminer l’année pendant laquelle l’arbre est mort alors que la deuxième rondelle a servi à mesurer l’humidité du bois. Cette mesure a été déterminée en pesant les
16
rondelles encore humides avant de les faire sécher dans un four à une température de 70 °C pendant quatre jours. Après cette période, elles ont été pesées à nouveau puis on a divisé le poids sec par le poids humide pour obtenir le pourcentage d’humidité de chaque rondelle (masse humide-masse anhydre/masse humide *100). En ce qui concerne la section de 1,50 m, elle a d’abord été écorcée en forêt à l’aide d’une plane à écorcer droite pour y dénombrer les trous d’entrée et de sortie des longicornes noirs. On a considéré que les trous ovales d’environ 8 mm de longueur et orientés de façon non perpendiculaire dans le bois étaient des trous d’entrée de larves de longicornes (Wilson, 1962). L’impact économique des dommages de longicornes a été évalué en utilisant la relation mise au point par Safranyik et Raske (1970) entre le pourcentage de perte de valeur par billot de pin tordu latifolié (Pinus contorta Dougl. Ex Loud.) et la densité de trous d’entrée de larves de Monochamus. Ils ont estimé des pertes légères (5 %) à partir de 5,4 trous/m², des pertes moyennes (11,5 à 18 %) lorsqu’il y a entre 10,8 et 16,1 trous/m² et des pertes élevées (>29 %) s’il y a plus de 32,3 trous/m².
L’année de mortalité des arbres échantillons a été déterminée à l’aide d’une analyse dendrochronologique. Les rondelles ont d’abord été séchées à une température ambiante de 23 °C pendant environ trois semaines dans une chambre de conditionnement au pavillon Gene-H. Kruger de l’Université Laval. Par la suite, chaque rondelle a été sablée trois fois : le premier sablage a été fait avec un papier de grade 80, le second avec un papier de grade 120 et le dernier avec un papier de grade 320. Les cernes de croissance ont été dénombrés sur chaque rondelle le long de trois rayons représentatifs à l’aide d’un binoculaire (Wild M4-63700, Heerbrugg, Switzerland) (8x-20x) au laboratoire de dendrochronologie de l’Université Laval. Les cernes ont été comptés à partir de l’extérieur de la rondelle, le premier cerne étant considéré comme provenant de l’année 2010 pour les rondelles prises avant la mi-juillet, parce que le bois final pour l’année 2011 n’avait pas encore eu le temps de se former. Par contre, pour les rondelles prises après la mi-juillet, le premier cerne a été considéré comme provenant de l’année 2011.
Les cernes de toutes les rondelles ont été numérisés, dénombrés de nouveau et mesurés à l’aide des logiciels CooRecorder (Version 7.4, mars 2011) et CDendro (Version 7.5, mars 2011). Afin de faciliter la datation, on a aussi identifié les cernes diagnostiques (cernes pâles, cernes de gel et cernes discontinus) pour chacune des rondelles des arbres témoins (TEM ; arbres vivants droits). Cette procédure a permis d’identifier trois cernes diagnostiques (1956, 1978, 1979) qui correspondent à ceux rapportés par Morin (1998). Ces cernes diagnostiques correspondent aux diminutions de croissance des arbres qui ont été observées lors des deux dernières épidémies de la tordeuse des bourgeons de l’épinette (Choristoneura fumiferana [Clem.]) (Morin 1998). Une fois que les cernes ont été dénombrés et mesurés, on a procédé à l’interdatation à l’aide du
17 logiciel CDendro qui fournit une courbe de référence conçue à partir de la largeur des cernes des arbres vivants, ce qui inclut les trois cernes diagnostiques. Ensuite, on a tracé les courbes pour chacun des arbres morts. En faisant correspondre les cernes diagnostiques à ceux des arbres témoins, on a pu estimer l’année de la mort de chacun des arbres.
3.2 Chablis expérimental à la Forêt Montmorency
La deuxième partie de l’étude comportait un volet expérimental visant à valider les résultats obtenus sur la Côte-Nord tout en ajoutant un volet temporel qui n’avait pas été couvert. Cette partie a été réalisée dans le sous-domaine bioclimatique de la sapinière à bouleau blanc de l’Est (Grondin et al., 1996, Bélanger, 2001) sur le terrain de la Forêt Montmorency de l’Université Laval (47º 19' 18" N, 71º 8' 49" O) (Figure 2). Ce sous-domaine est localisé au sud de la forêt boréale et couvre une bande de 150 km de large qui traverse le Québec entre les 47º et 50º de latitude nord (Ministère des Ressources naturelles (MRN) 2000). Ce sous-domaine couvre une superficie de 55 970 km² constituée principalement de peuplements résineux et mélangés et son altitude varie entre 600 et 1000 mètres (Larouche, 2005). Le bouleau blanc (Betula papyrifera Marsh.), l’épinette blanche (Picea glauca (Moench) Voss), l’épinette noire (Picea mariana (Mill.) B.S.P.) et le peuplier faux-tremble (Populus tremuloides Michx.) sont les espèces d’arbres les plus fréquemment associées au sapin baumier (Larouche, 2005). Le climat est caractérisé par une température annuelle moyenne avoisinant 0 ˚C ainsi que par d’abondantes précipitations totalisant environ 1400 mm par année. Les feux y sont peu fréquents en raison de ces fortes précipitations. La tordeuse des bourgeons de l’épinette y est le principal facteur de perturbation, mais les chablis endémiques sont aussi nombreux (Larouche, 2005). La Forêt Montmorency est représentative des grandes forêts résineuses de l’est du Québec dominées par le sapin baumier (Département des sciences du bois et de la forêt de l’Université Laval, 2004).
Le dispositif mis en place avait pour but de répondre d’une manière plus précise aux objectifs de l’étude en contrôlant la date du chablis et l’état des arbres. Un dispositif factoriel (2 x 4 x 2) complètement aléatoire a été établi avec comme facteurs fixes l’espèce végétale (épinette noire et sapin baumier), la catégorie d’arbres (déracinés, coupés, annelés, vivants sur pied) et la période d’application des traitements (automne 2011 et printemps 2012). Chaque catégorie d’arbre correspond à ce que l'on retrouve en forêt après un chablis naturel: des arbres renversés (déracinés), des arbres brisés (coupés) et des arbres moribonds (annelés), en plus d’arbres témoins (vivants sur pied). Vingt arbres de chacune des deux espèces ont été identifiés pour chaque catégorie d’arbre et pour chaque période de chablis pour un grand total de 320 arbres échantillons
18
dans ce dispositif. Pour être sélectionnés, les arbres devaient avoir un DHP supérieur à 16 cm. Nous avons utilisé une débusqueuse pour renverser des arbres et une scie à chaine pour en abattre d’autres. L’annelage a été réalisé manuellement en retirant l’écorce sur une bande de 5 cm de largeur autour de l’arbre à une hauteur de 1,3 m. Les traitements ont été réalisés le 27 octobre 2011 (chablis d’automne) et le 15 juin 2012 (chablis de printemps).
La fréquence des visites du longicorne noir sur les différents types d’arbres a été mesurée à l’aide de pièges à impact fixés au tronc des arbres échantillons. Des pièges ont été installés (10 pièges/traitement/essence) le 19 juin 2012 et sont restés sur place jusqu’au 29 août 2012, afin de vérifier les taux de visite des adultes du longicorne noir pendant la première saison d’activité après la perturbation. Une solution pure d’éthylène glycol a été utilisée dans les pots collecteurs pour tuer et préserver les spécimens. Les pièges ont été visités à toutes les semaines pour récolter les échantillons et remplacer la solution d’éthylène glycol. Les échantillons ont été triés au laboratoire et les spécimens de longicornes ont été identifiés à l’espèce. La teneur en eau du tronc des arbres a été mesurée à partir de deux carottes de bois (séchées et pesées de la même façon que pour les rondelles) prélevées en fin de saison (29 août 2012). À la fin du mois de septembre, 5 arbres par traitement et par essence sur lesquels nous n’avions pas installé de pièges ont été abattus et des bûches de 1 m de longueur (centrée à 1,3 m) ont été transportées à l’insectarium du Centre de Foresterie des Laurentides (CFL). Les bûches ont été écorcées avec un ciseau à bois, ce qui nous a permis de dénombrer les trous d’entrée de larves de longicornes et de récolter les larves présentes.
3.3 Analyses statistiques
Dans cette étude, des modèles de régression ont été construits en tenant compte de la présence des trous d’entrée de larves de longicornes noirs (Côte-Nord), des adultes dans les pièges et des larves de
Monochamus scutellatus scutellatus sous l’écorce d’épinettes noires et de sapins baumiers (Forêt
19
3.3.1 Chablis naturel sur la Côte-Nord
Les données analysées sont celles obtenues par le décompte des trous d’entrée de larves de longicorne noir trouvés sous l’écorce des billots d’épinette noire et de sapin baumier qui ont été écorcés sur place. Un modèle de régression de Poisson généralisé avec excès de zéros et facteur aléatoire (où le peuplement était le facteur aléatoire), a été ajusté aux données afin de comparer le niveau d’attaque de M. s. scutellatus en fonction de l’espèce d’arbre, du DHP, de l’état de l’arbre (appuyé, déraciné, cassé ou debout) et de l’année de mortalité de l’arbre. Ce type de régression a la particularité de prendre en compte le nombre élevé de zéros (absence de trou d’entrée sur plusieurs échantillons) et les nombres entiers qui caractérisent le décompte de trous de larve dans le jeu de données. La procédure GLIMMIX de SAS3 a été utilisée pour les analyses. Les comparaisons deux à deux ont été effectuées avec la méthode LSD protégée de Fisher. La force de la relation entre la sévérité du chablis et le nombre de trous d’entrée de longicornes a été calculée en utilisant la méthode de Nakagawa et Schielzeth (2013) (Équation 1).
Équation 1. Méthode pour obtenir R2 à partir des modèles linéaires généralisés avec effets. Rg2 = 1-(L0/Lβ)2/n ,
où, L0 représente le « likelihood » du modèle nul, Lβ représente le « likelihood » du modèle complet et n représente le nombre d’observations.
De façon à déterminer s’il y a un lien entre le nombre d’années après la mortalité des arbres et le nombre de trous d’entrée de longicornes, un modèle de régression segmentée a été ajusté aux données dans le but de trouver un point de jonction indiquant un seuil à partir duquel les arbres étaient moins affectés par l’insecte. Ce modèle de régression segmenté était composé de deux modèles linéaires simples ajustés à l’aide de la procédure NLIN de SAS.
20
3.3.2 Forêt Montmorency
Une première analyse a été faite sur les adultes de M. s. scutellatus capturés à l’aide des pièges à impact fixés au tronc. Un modèle de régression de Poisson généralisé a été ajusté aux données afin de comparer le nombre de visites pour l’ensemble de la saison en fonction de l’essence forestière, du DHP, de la position des arbres (déracinés, coupés, annelés et témoins), de l’humidité des arbres et de la période du chablis. Des « Least Square Means (LS-means) » ont été calculés pour déterminer si les différences entre les moyennes étaient significativement différentes (p<0,05) lorsque l’ANOVA indiquait des différences significatives. Les analyses ont été faites en utilisant la procédure GLIMMIX de SAS. En ce qui concerne l’analyse des larves de
M. s. scutellatus, on a utilisé un modèle de régression multiple à distribution binomiale négative avec excès de
zéros pour déterminer la préférence des femelles de M. s. scutellatus en fonction de l’essence forestière, du DHP, de la position des arbres (déracinés, coupés, annelés et témoins), de l’humidité des arbres et de la période du chablis. Les « Least Squares Means (LS-means) » ont été calculés pour déterminer si les différences entre les moyennes étaient significativement différentes (p<0,05) lorsque l’ANOVA indiquait des différences significatives. Les données ont été transformées (Log x+1) pour satisfaire les principes de normalité et d’homogénéité des variances. Les analyses ont été faites en utilisant la procédure GENMOD de SAS parce que la convergence des itérations n’était pas atteinte avec la procédure GLIMMIX. Les comparaisons deux à deux ont été effectuées avec la méthode LSD protégée de Fisher.
4 Résultats
4.1 Chablis naturel sur la Côte-Nord
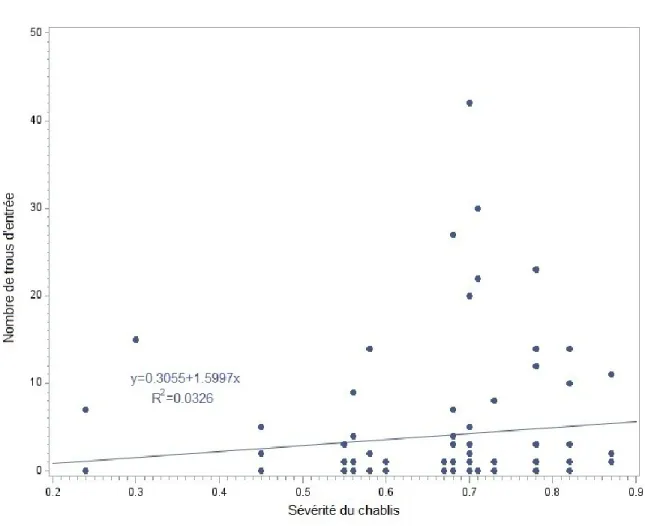
On a dénombré un total de 354 trous d’entrée de larves de longicornes sur les 160 billots écorcés pour une moyenne (± écart type) de 4,3 ± 8,3 trous d’entrée/m2. Le nombre moyen de trous d’entrée de larves de longicorne a varié de 0 à 41,7 trous/m² (Tableau 2). Sur 14 % des arbres étudiés, plus de 11 trous d’entrée par mètre carré ont été dénombrés, entraînant ainsi des pertes en valeur de plus de 15 % si on se réfère à la relation établie par Safranyik et Raske (1970) (Fig. 3). Chez le sapin, on a trouvé une densité 3,5 trous/m² alors qu’elle était de 5 trous/m² chez l’épinette noire. Selon le modèle de régression utilisé, seuls la sévérité du chablis (F=5,89, p = 0,0168) et le nombre d’années après la mort des arbres (F = 3,77, p = 0,0546) ont montré un effet significatif sur le nombre de trous d’entrée de larves de longicornes. Le modèle n’a montré aucun effet significatif du DHP, de l’état de l’arbre, de l’humidité et de l’essence forestière sur le nombre de trous d’entrée
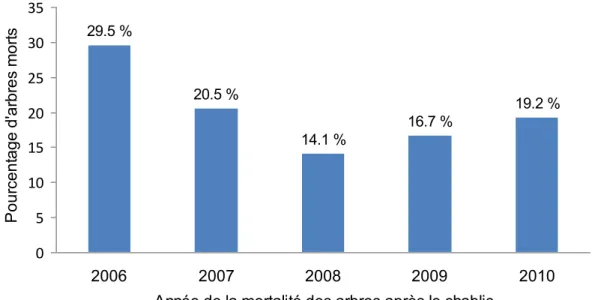
21 des larves. La relation significative et positive entre la sévérité du chablis et le nombre de trous d’entrée de larves de longicornes (Fig. 4) a cependant un R2 modeste de 0,0326, obtenu en utilisant la méthode de Nakagawa et Schielzeth (2013). Le chablis qui a affecté les arbres évalués a eu lieu à la fin de l’été de 2006, mais la mortalité s’est poursuivie au cours des années suivantes (Fig. 5). De cette façon, l’abondance et la distribution des ressources pour les insectes xylophages à la suite de cette perturbation sont réparties dans le temps.
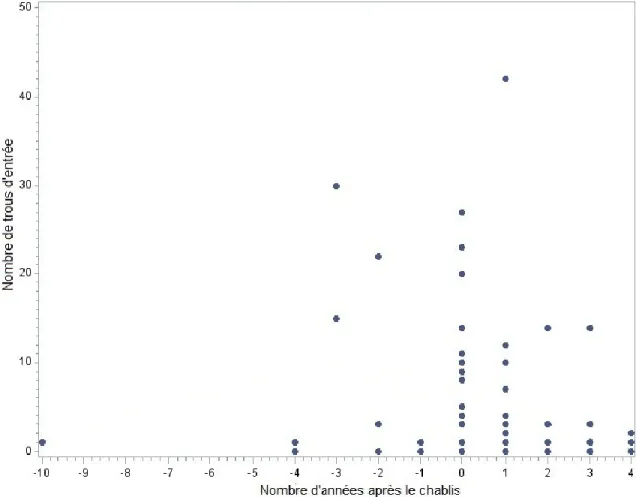
Le modèle de régression segmentée utilisé pour relier le nombre de trous d’entrée de longicorne à l’année de mortalité des arbres n’a pas convergé à cause du peu de données disponibles pour les arbres morts avant le chablis, particulièrement entre 10 et 3 ans avant le chablis (Fig. 6). Ces résultats tendent toutefois à indiquer que les arbres morts depuis plus de trois ans avant le chablis ont été très peu utilisés par le longicorne noir. De plus, à la suite du chablis, le nombre de trous de longicorne tend à diminuer avec le temps (Fig. 6).
4.2 Chablis expérimental à la Forêt Montmorency
Un total de 4214 insectes a été capturé dans les pièges à impact fixés au tronc des arbres, dont 4194 coléoptères et 20 siricides. Parmi ces captures, 378 étaient des longicornes dont 353 ont été identifiés comme
Monochamus scutellatus scutellatus. (260 mâles et 93 femelles). Les 25 autres spécimens de longicornes ont
été identifiés comme Acmaeops proteus proteus (10 specimens), Judolia montivagans (8 spécimens),
Neoclytus leucozonus (3 spécimens), Pygoleptura nigrella (2 spécimens), Xylotrechus undulatus (1 spécimen)
et Monochamus mutator (1 spécimen). À la fin de l’été, nous avons dénombré 611 larves de M. s. scutellatus après l’écorçage des billots.
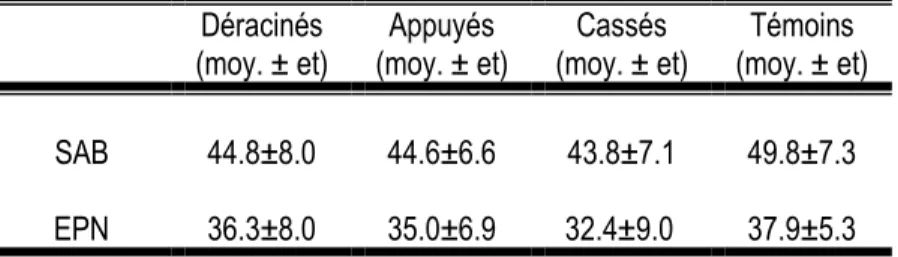
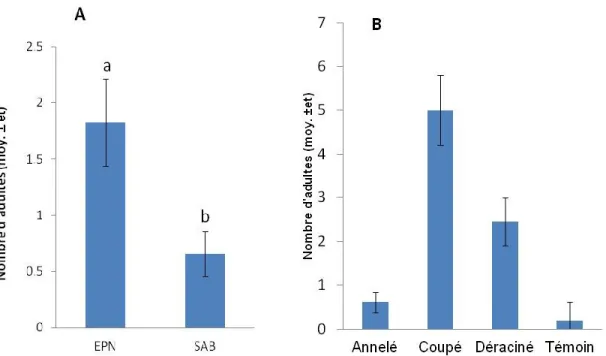
Le modèle statistique montre que les variables présentant des effets significatifs sur la fréquence des visites de M. s. scutellatus sont l’essence forestière (F = 0,81, p = 0,3690) et l’état de l’arbre (F = 11,04, p < 0,0001) (Tableau 3, Figure 7). L’épinette noire a reçu environ trois fois plus de visites d’adultes de M. s. scutellatus. Les arbres coupés ont été les plus visités, soit deux fois plus que les arbres déracinés et dix fois plus que les arbres annelés, ces derniers n’étant pas significativement différents des arbres témoins.
22
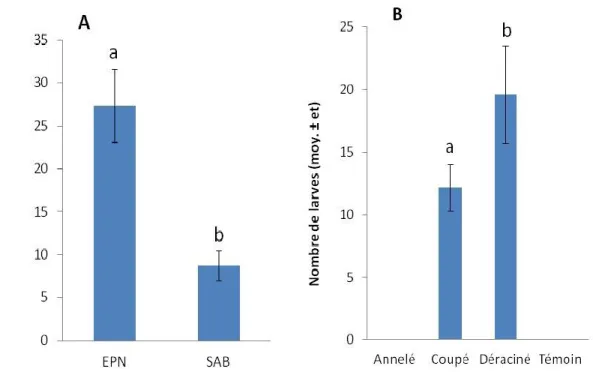
De façon analogue, les variables qui ont influencé significativement le nombre de larves de longicorne noir sont l’essence forestière (F = 14,59, p = 0,0001) et l’état de l’arbre (F = 3,08, p = 0,0791) (Tableau 4, Figure 8). L’épinette noire a été trois fois plus colonisée par des larves de M. s. scutellatus que le sapin baumier. Les arbres déracinés ont été les plus colonisés, environ 1,5 fois plus que les arbres coupés. Cependant, aucune colonisation par M. s. scutellatus n’a été observée sur les arbres annelés et témoins.
5 Discussion
5.1 Côte-Nord
Sur la Côte-Nord, l’abondance de trous de longicornes dans le bois des arbres renversés par le vent a été influencée par la sévérité du chablis et le nombre d’années après la mort des arbres. Les mécanismes de défense d’arbres récemment tombés à cause d’un chablis sont fortement diminués et ces arbres sont donc plus faciles à coloniser (Grégoire et al., 1997). Quand ils sont renversés par le vent, les arbres émettent des substances volatiles que les arbres sur pied n’émettent pas (Austarå et al., 1986 ; Grégoire et al., 1997). Ces substances volatiles sont perçues par les insectes, ce qui leur permet de trouver les arbres qui leur sont convenables (Duduman et Vasian, 2012). Ainsi, Raske (1972) indique que les adultes de M. s. scutellatus peuvent voler sur une distance d’environ 10 km, tandis que Ross (1966) soutient que les adultes de M. s.
scutellatus sont capables de voler jusqu’à 96 km au cours d’une période de cinq semaines. Dans le cas d’un
chablis important, les substances volatiles des conifères sont libérées en grande quantité, ce qui permet aux longicornes de repérer facilement ces arbres (Groot et Nott, 2004). Monochamus scutellatus semble attiré par l’odeur de la résine des arbres blessés (Clemence, 1916), plus particulièrement par la térébenthine, qui est un mélange de monoterpènes produits par les conifères (Gardiner, 1975 ; Dyer et Seabrook, 1978). Aussi, lorsque les arbres commencent à se décomposer, ils libèrent une grande quantité d’éthanol associé au processus de fermentation et ces vapeurs d’éthanol ont le même effet sur les insectes xylophages que les substances volatiles primaires (Duduman et Vasian, 2012). Pour ajouter à ces stimuli, M. s. scutellatus réagit aux kairomones (des substances chimiques produites et libérées par un organisme vivant et qui bénéficient à l’organisme qui recoit ces substances, mais pas à l’organisme les ayant émises (Bashford, 2008)) qui sont libérées par d’autres espèces d’insectes xylophages comme les scolytes et cette information peut contribuer à la colonisation de l’habitat (St-Germain et al., 2004 ; Erbilgin et Raffa, 2000). Finalement, si le nombre d’arbres renversés est important, les populations d’insectes xylophages vont rapidement augmenter (Grégoire et al., 1997), ce qui contribuera à augmenter les dommages de longicornes dans le bois des arbres renversés.
23 Le chablis est une perturbation naturelle qui a souvent une dynamique progressive de sorte que quand il touche un peuplement, ce ne sont pas tous les arbres qui meurent en même temps, mais seulement une partie, ce qui étale la mortalité dans le temps (Ruel et., al 2010; Bouget et Duelli, 2004). Cette dynamique permet de fournir un substrat intéressant aux insectes xylophages pendant une période de temps relativement longue. Cette progression du chablis est semblable à une série de petites perturbations qui, à plus long terme, ont la même importance qu’une grande perturbation, mais qui contribuent significativement à l’hétérogénéité de l’habitat (Gandhi et al., 2007). Le feu n’a pas une dynamique progressive comme le chablis, mais la sévérité du brûlage a une influence sur la susceptibilité des arbres aux attaques des insectes (McCullough et
al., 1998; Saint-Germain et al., 2004).
Dans notre étude, la sévérité du chablis a montré un effet significatif sur le niveau d’attaque des longicornes, ce qui s’apparente aux feux de forêt où la sévérité du brûlage est le facteur le plus important pour prédire l’abondance des insectes xylophages après une perturbation (Boulanger et al., 2010). Après une perturbation comme le chablis ou le feu, les arbres affectés commencent à libérer des composés volatils qui les rendent attractifs aux insectes xylophages (Kula et Zabecki, 2006). La quantité de composés attractifs libérés est fonction de la sévérité et des dimensions de la perturbation.
Nos résultats indiquent que le niveau d’attaque par les longicornes tend à diminuer en fonction du nombre d’années après la mort des arbres à cause d’un chablis. Il semble que les arbres morts depuis plus de trois ans après la perturbation subissent moins d’attaques que ceux morts plus récemment, mais nous ne disposons que de très peu de données pour soutenir cette hypothèse. Ainsi, après les trois premières années suivant leur mort, la présence de longicornes commence à diminuer possiblement à cause de la dégradation du bois qui en fait un milieu moins favorable au développement de l’insecte qui tendrait alors à migrer vers des arbres plus convenables. Cette hypothèse est appuyée par Graham et Knight (1965) qui mentionnent que les arbres morts récemment sont plus attirants pour les insectes xylophages, ce qui pourrait expliquer la présence plus élevée de l’insecte pendant la première année après un chablis (Ruel et, al., 2010). Un chablis sévère qui a lieu entre les mois de juin et août semble particulièrement attrayant pour la reproduction des insectes xylophages (Gandhi et al., 2007). Nos résultats montrent aussi que des arbres morts au cours des trois années précédant le chablis de 2006 ont été attaqués par les longicornes (Fig. 6). Les conditions du bois de
24
ces arbres étaient donc suffisamment bonnes pour que les longicornes puissent y déposer leurs œufs et s’y développer ou encore les arbres avaient été colonisés avant le chablis.
Les pertes économiques de la valeur du bois peuvent être évaluées par rapport à la densité de trous d’entrée de larves de longicornes. Safranyik et Raske (1970) ont caractérisé la relation entre le pourcentage de perte de valeur par billot de pin tordu latifolié (Pinus contorta Dougl. Ex Loud.) et la densité de trous d’entrée de larves de Monochamus. Ils ont estimé des pertes légères (5 %) à partir de 5,4 trous/m². des pertes moyennes (11,5 à 18 %) lorsqu’il y a entre 10,8 et 16,1 trous/m² et des pertes élevées (>29 %) s’il y a plus de 32,3 trous/m². Dans notre étude, la densité des trous d’entrée chez le sapin baumier était de 3,5 trous/m² comparativement à 5 trous/m² chez l’épinette noire. En se basant sur la relation établie par Safranyik et Raske (1970), on estime donc les dommages comme légers et les pertes à environ 5 % de la valeur des bois. Par ailleurs, 14 % des arbres affectés dans notre étude comptaient plus de 11 trous/m² pour des pertes de valeur économique >10 % selon la relation établie par Safranyik et Raske (1970). Or, l’étude de Boucher et Hébert (2010), réalisée en 2008 dans le même chablis, soit deux années après la perturbation, rapportait que 60 % des arbres affectés montraient plus de 15 trous/m², ce qui indique que le niveau d’attaque par le longicorne est élevé immédiatement après la perturbation, mais qu’il tend à diminuer avec le temps sur les arbres résiduels qui s’ajoutent au fil des ans. En fait, comparativement à Boucher et Hébert (2010), notre échantillonnage intégrait des arbres morts après le chablis initial et ces arbres avaient subi moins d’attaques par le longicorne noir, expliquant ainsi la sévérité moins élevée de nos estimés.
Nos résultats ne montrent pas d‘effet significatif de variables comme l’humidité, le DHP et l’essence forestière. Dyer et Seabrook (1978) rapportent que les femelles de Monochamus scutellatus (Say) et M. notatus (Drury) laissent peu ou pas d’œufs sur les arbres qui ont moins que 10 % ou plus que 60 % de teneur en eau. Or, nos données montrent que le pourcentage moyen d’humidité des arbres renversés par le vent est supérieur à 45 % pour le sapin baumier et supérieur à 35 % pour l‘épinette noire (Tableau 5). De plus, la faible variation du DHP des arbres étudiés (Tableau 1) n’a possiblement pu permettre l’expression de l’importance de cette variable qui est reconnue comme importante après un feu (Saint-Germain et al., 2004). Ces auteurs indiquent que la faible densité des larves par rapport au DHP des arbres peut être expliquée par le faible taux de survie de l’insecte entre le stade de l’œuf et le troisième stade larvaire, celui où l’insecte entre dans le xylème.