Étude de la régulation génique par les microARN dans les cellules germinales
Texte intégral
Figure
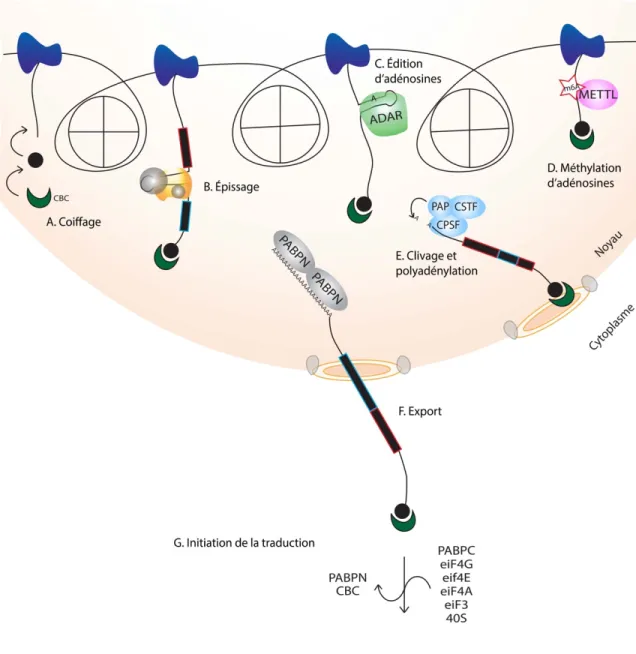
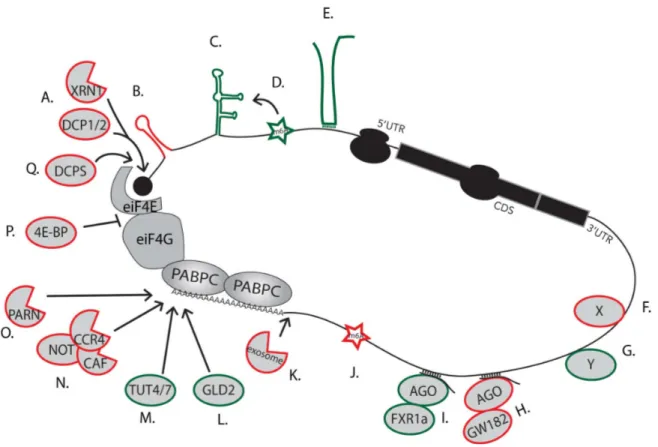
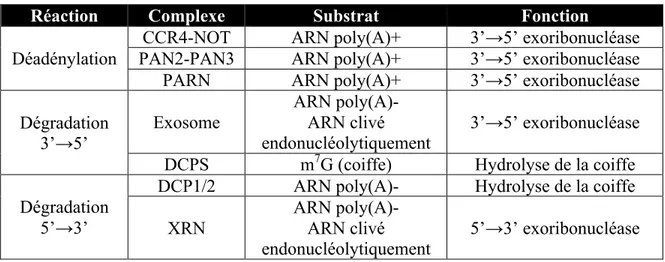
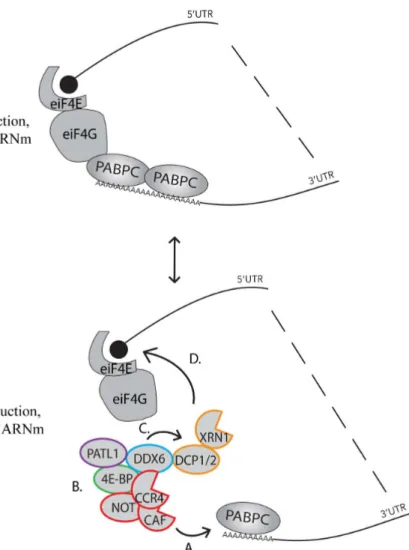
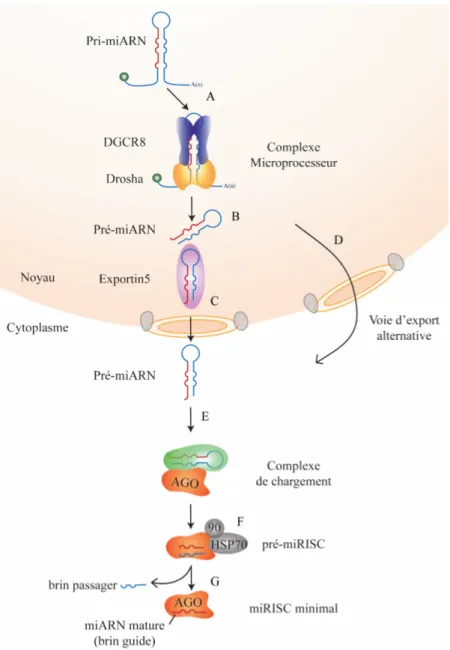

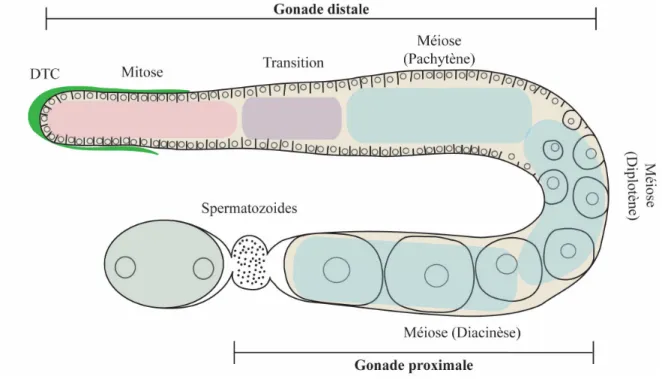



Documents relatifs
- Régulation de l’expression des enzymes de conversion de la D-Met, D- et L-HMTBA Muscles : turnover protéique/croissance. - Régulation du turnover protéique en fonction des apports
Figure 38: (A) Abondance du nombre de pics autour du TSS (en abscisse) détectés par Quest, SISSR et SICER pour la protéine Chd8 sur ses propres gènes cibles.. A partir de liste de
Nous avons également démontré que la dilatation dépendante de l’endothélium à la BK (agoniste des récepteur B2) et la dilatation indépendante de l’endothéÏium au SNP
MAPK/ERK dans la régulation du clivage de la PrP c médiée par les récepteurs muscariniques : 1/ La thréonine 735 est localisée au niveau de la queue cytoplasmique d’ADAM17 dans
Enfin, comme les transfections transitoires avec différentes quantités de NR0B2 n’ont pas permis d’observer de répression de l’activité du promoteur Insl3 dans les cellules
CRMPR – Centre Régional de Médecine Physique et de Réadaptation... La chromatine est fine et un nucléole est
Le travail présenté dans le cadre de ce mémoire concerne l’utilisation de la théorie de la fonctionnelle de la densité dépendante et indépendante du temps
Nous avons par ailleurs réalisé une étude théorique par des calculs de chimie théoriques en utilisant la théorie fonctionnelle de la densité dépendante (DFT) et
