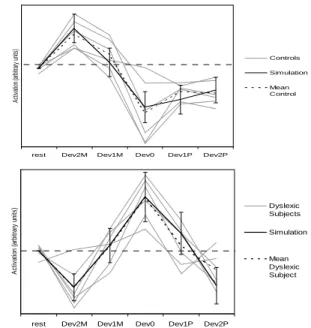
Dynamic Bayesian modeling of the cerebral activity
7
0
0
Texte intégral
Figure
![Figure 1: Large-scale network involved in phoneme monitor- monitor-ing, according to results from [Démonet, et al., 1994]](https://thumb-eu.123doks.com/thumbv2/123doknet/2355123.37400/2.918.520.792.675.859/figure-network-involved-phoneme-monitor-monitor-according-demonet.webp)

Documents relatifs