UNIVERSITE PAUL SABATIER TOULOUSE III - Sciences
THESE
En vue de l’obtention du titre de
DOCTEUR DE L’UNIVERSITE PAUL SABATIER
Spécialité : Physiopathologie moléculaire, cellulaire et intégrée
Présentée par
Marie DANCE
NOUVEAUX MECANISMES D’ACTIVATION DES VOIES Ras/MAPK ET
PI3K PAR GAB1 EN AVAL DES RECEPTEURS À ACTIVITE TYROSINE
KINASE :
EXEMPLE DES RECEPTEURS DE L’EGF ET DU VEGF
Thèse dirigée par le Dr. Patrick RAYNAL
Soutenue le 21 septembre 2007 devant le jury composé de :
Dr. Jean GIUDICELLI INSERM U145, Nice Rapporteur
Dr. Patrick MAYEUX INSERM U567, Paris Rapporteur
Dr. Béatrice PARFAIT Université René DESCARTES, Paris V Examinateur
Pr. Bertrand PERRET INSERM U563, Toulouse Président
Dr. Patrick RAYNAL INSERM U563, Toulouse Examinateur
A Olivier,
A mes parents, A mon frère,
Remerciements
Je tenais tout d’abord à remercier les personnalités qui ont siégé à ce jury de thèse :
Monsieur le Professeur Bertrand Perret, je vous remercie de m’avoir fait l’honneur de présider ce jury de thèse. Merci également de m’avoir accueillie au sein de votre département, ainsi que pour votre écoute, votre disponibilité et pour l’intérêt que vous avez porté à mes travaux durant ces quatre années. Vous avez largement contribué à la valorisation de cette thèse.
Monsieur le Professeur Jean Giudicelli, je vous remercie d’avoir accepté la lourde tache de rapporteur avec autant de gentillesse et de disponibilité. Je tiens aussi à souligner que vous avez énormément contribué à la richesse de la discussion scientifique lors de la soutenance.
Monsieur le Docteur Patrick Mayeux, je vous remercie d’avoir vous aussi accepté cette
charge de rapporteur, et d’avoir porté sur ce manuscrit et lors de cette journée, un regard pertinent sur l’ensemble de mes travaux. Les réflexions scientifiques n’en auront été que plus profondes.
Madame le Docteur Béatrice Parfait, je vous remercie tout d’abord d’avoir accépté de
siéger à ce jury de thèse et d’avoir représenté aussi bien la gente féminine. Merci pour votre ouverture d’esprit sur le monde du signal, votre gentillesse et pour vos interventions pertinentes lors de cette journée.
Monsieur le Professeur Jean-Pierre Salles, je vous remercie d’avoir accepté de siéger à ce jury de thèse. Merci également pour votre soutien, la confiance que vous m’avez accordée et vos conseils tout au long de cette thèse.
Monsieur le Docteur Patrick Raynal pour avoir encadré ce travail tant durant le DEA que pendant ces 4 années de thèse. Merci d’avoir cru en moi et de m’avoir fait confiance depuis le début et ce jusqu’à la fin de ce doctorat. Merci aussi pour tout ce que tu as apporté à ces travaux, tant grâce à ton recul scientifique et tes précieux conseils, que pour m’avoir appris la rigueur de la progression et de l’analyse scientifique. Merci de m’avoir accompagnée dans le monde de la recherche. Je sais que la voie que je choisis n’est pas forcément celle à laquelle tu t’attendais ou du moins celle que tu me conseillerais, mais merci de ne pas m’avoir jugée et de continuer à me soutenir. En attendant que nos chemins se recroisent peut être un jour, je te souhaite le meilleur, tant d’un point de vue professionnel que personnel. Encore merci pour tout…
Remerciements
J’entame maintenant l’une des parties les plus agréables lors de la rédaction d’une thèse, mais pourtant la partie la plus délicate et personnelle. Je préfère m’excuser dès à présent auprès des personnes m’ayant côtoyé et dont le nom n’apparaît pas ci-après, mais je vous assure que ce n’est pas volontaire et que l’exercice est plus difficile qu’il n’y paraît. En tout cas, n’y voyez aucune impolitesse ou ingratitude de ma part, mais plutôt une certaine émotion et un manque de temps qui font que les idées ne sont parfois pas aussi claires qu’on ne le voudrait.
Je vous invite maintenant à me suivre dans cette visite guidée de notre fameux bâtiment C qui trouve la force de rester planté bien droit malgré les nombreux stress subis ces derniers mois.
Au risque de manquer d’originalité, je vais bien entendu commencer par l’endroit que je connais le mieux du bâtiment : le labo 107 !!! Mes premiers mots seront tout d’abord pour Armelle sans qui cette thèse ne serait pas vraiment ce qu’elle est… Merci tout d’abord à toi aussi d’avoir cru en moi pour le DEA, je pense que la décision de Patrick de me soutenir cette année-là ne t’est pas étrangère. On s’est peu vu à l’époque, mais ton retour cette dernière année m’a permis de te (re)découvrir. Merci donc pour ta disponibilité, ton recul scientifique, tes conseils et ton investissement dans ce manuscrit et lors de la préparation de ma soutenance. Merci aussi pour nos nombreuses discussions, parfois scientifiques, parfois moins, ainsi que tous ces bons moments passés au 107. Je sais que cette relation continuera au delà du laboratoire, il me tarde déjà de voir ton futur petit nid douillet… En tout cas, je te souhaite le meilleur pour la suite, tant d’un point de vue professionnel que personnel. Merci ensuite à Carla, pour ta disponibilité et ta motivation. Je tenais aussi à te dire que même si nos rapports ont parfois été délicats, j’ai apprécié de travailler sur ce papier avec toi et je suis heureuse que notre duo ait porté ses fruits, je ne l’aurais pas fait sans ton aide. Bonne chance pour la soutenance de ta thèse et ton futur post-doc ainsi que pour réaliser tous tes projets personnels. Je passe maintenant à notre pédiatre préféré, Thomas. Je ne remercierais jamais assez « les chefs » de nous avoir amené quelqu’un comme toi, dans ce monde de filles. J’admire ton courage pour avoir entamé cette thèse en continuant à côté tes activités cliniques et la « gestion » de toute ta petite famille. Merci pour ta simplicité, ta disponibilité, ta gentillesse, ta motivation, ta bonne humeur et ton ouverture d’esprit. Surtout ne changes rien, et je suis sûre que tout ira pour le mieux par la suite… Récemment, le 107 s’est vu ajouter un autre représentant de la gente masculine : Jean-Philippe. Je pense que tu as maintenant bien pris tes marques dans ce monde de biologie cellulaire et je te souhaite bonne chance pour ta carrière et profites bien de tes petits si craquants !!! Mais attention les garçons, accrochez vous, 2007 marque l’arrivage de nouvelles filles au 107 (il faut bien prendre la relève…) : Marie-Anaïs (excellent ton gâteau au chocolat…), à qui je souhaite d’obtenir sa bourse de DEA et de concrétiser ses objectifs, Audrey qui, je suis sûre, va dignement prendre le relais des thésards du 107 et Sophie que je n’ai pas encore rencontré mais à qui je souhaite bonne chance pour la suite. Merci aussi aux stagiaires et collaborateurs qui ont parcouru ces murs ces dernières années (Claudia notre roumaine, Marek notre polonais,…) et tout particulièrement Delphine qui nous a tant touché par sa « naïveté » et sa spontanéité. Mais le 107 ne serait pas vraiment ce qu’il est sans Alex, avec qui j’ai partagé la majeure partie de cette thèse. Merci pour toutes ces années d’amitié, pour nos discussions qui ont si
Remerciements
bien su nous rapprocher. Merci pour ton dévouement sans faille quand il s’agissait d’aider les autres, merci pour nos nombreux fous rires, pour ton aide tant d’un point de vue scientifique que personnel. Ma thèse n’aurait pas été la même sans toi, je serais toujours là si tu as besoin. Conserves tes valeurs qui t’honorent et que tu sais si bien défendre et apprends juste à avoir un peu plus confiance en toi et aux autres. J’espère que cette amitié durera encore longtemps et que nos chemins se recroiseront bientôt. Je suis sûre que tu vas brillamment réussir ta carrière et je ne te souhaite que du bonheur avec Alain et tes proches.
Je sors maintenant du laboratoire pour aller vers le « 104-105 » (il est difficile de dissocier les 2…). Merci tout d’abord à Monique, pour ses qualités humaines indéniables, sa gentillesse, et sa disponibilité dès qu’il s’agissait d’obtenir un conseil scientifique. Je te souhaite pleins de bonnes choses pour la suite, c’est certes une page qui se tourne mais je suis sûre que la suite ne sera que bonheur et réussite, notamment grâce à tout ce que tu es capable d’apporter à ce qui sont dans le besoin. Merci pour toutes ces leçons d’humanité. Merci aussi à Muriel, tout d’abord pour tout ce qu’elle a apporté de positif au laboratoire par son dynamisme, ses conseils, ses qualités scientifiques et son aide précieuse (souvent) apportées aux étudiants. Merci pour ton caractère optimiste, ta franchise qui m’a parfois aidé à ouvrir les yeux et à considérer les choses sous un autre angle, ta disponibilité et les valeurs que tu défends si bien. Je ne vais pas loin, et j’espère qu’on se recroisera un de ces jours autour d’une tasse de café. Tu as maintenant pris sous ton aile notre petite Anne, et c’est une excellente chose. Je ne sais pas si tu t’en rends compte, Anne, mais tu es une vraie perle rare, tant pour ce qui est des relations professionnelles que tu entretiens que d’un point de vue personnel. On avait eu une bonne intuition avec les filles quand on t’avait vu arriver pour ton DESU et ça s’est confirmé par la suite. J’espère que notre amitié naissante va continuer au-delà de cette thèse. En tout cas, ne changes rien, et je te souhaite le meilleur pour ce doctorat (je te fais entièrement confiance, tu vas briller) ainsi que pour tous tes projets personnels. Le duo Muriel/Anne fonctionne maintenant parfaitement bien avec le duo Fabienne/Hélène. Fabienne, je ne te remercierais jamais assez pour ton soutien lors de ma thèse, notamment dans les moments de doute, mais aussi lors de mon DEA. Je sais que tu n’aimes pas trop qu’on te le rappelle, mais même si tu as l’impression de ne pas avoir fait grand-chose, ton soutien lors de cette période a beaucoup compté pour moi. Merci aussi pour tes nombreux fous rires, notamment avec Muriel, de même que pour tes qualités scientifiques, humaines, et pour ton grand cœur. J’espère te recroiser toi aussi un de ces jours, histoire d’aller prendre un café en papotant. Je ne peux pas évoqué Fabienne lors de mon DEA sans penser à Geneviève. Merci pour cette année partagée, pour nos fous rires, pour tes qualités humaines uniques et pour les leçons de maturité que tu nous faisais souvent avec Flo lorsqu’il le fallait. Bonne continuation avec tout ton petit monde qui ne cesse de s’agrandir… C’est maintenant au tour d’Hélène de prendre le relais et elle le fait avec brio. Dommage que je ne reste
Remerciements
Dotée d’un caractère aussi fort que son cœur est grand, je ne te remercierais jamais assez pour tout ce que tu as fait pour nous (moi) au quotidien… Merci pour ta franchise, ta disponibilité et ta générosité. Le thème B n’aurait pas la même saveur sans toi… Tu travailles maintenant avec
Safouane, que j’ai connu entouré de ses drôles de dames à l’époque du 106. Merci à toi pour toutes
nos discussions, scientifiques et autres, et pour ton sourire et ta bonne humeur qui ne te quittent que rarement. Bonne chance pour la suite, je suis sûre que tout va se passer pour le mieux. Enfin, la dernière et non des moindres, Nicole. Merci pour ta disponibilité, ton calme qui ne semble jamais te quitter et pour ta gentillesse naturelle… Tu as droit aussi à un merci tout spécial pour m’avoir laissé ta carte d’accès qui m’a évité bien des ampoules aux pieds et un agacement supplémentaire en cette fin de thèse. En tout cas, je te souhaite pleins de bonnes choses avec ta petite famille et notamment tes jolies miss et ton petit chien… Je terminerais le 104-105 par quelqu’un qui m’a énormément apporté : Stef. Je ne sais par où commencer mais merci pour tout : pour tes qualités humaines uniques, pour ton sourire, ta gentillesse, ta franchise, ta bonne humeur. Merci d’avoir su me donner un coup de pied au derrière quand j’en avais besoin, mais aussi pour m’avoir soutenue dans les moments de doutes et dans les moments de délires « karaokesques ». Je sais qu’on continuera à se donner des nouvelles malgré la distance et qu’on se retrouvera très vite sur Toulouse. En attendant, je te souhaite bonne chance avec tes petites souris et pleins de bonheur dans ta vie personnelle. Le thème B s’est aussi vu grandir avec l’arrivée du CRC. Ah, quel bonheur de vous avoir avec nous maintenant… Merci Françoise pour ta gentillesse, ton naturel, ta spontanéité et pour tous ces moments passés en ta compagnie. J’espère qu’il y en aura pleins d’autres, au bord de ta piscine ou ailleurs, mais en tout cas, je ne vais pas loin donc il n’y a aucune raison de ne plus se voir. Promis, je te graverais « fais la poule » pour que tu conserves ta bonne humeur et ta générosité encore longtemps… Maintenant, à ton tour Marianne. Là aussi j’aurais tant à dire… Merci tout d’abord pour ton soutien et ton écoute lors de tous mes moments un peu « bas », mais merci aussi de me « ressembler » ou me « soutenir » autant (je sais pas trop comment dire), sur tout ce qui concerne certains de nos goûts artistiques qui laissent les autres (à part peut-être Françoise), souvent perplexes. Grâce à toi je me sens moins seule et j’espère que cette amitié se prolongera encore longtemps, histoire de papoter encore bien des heures… Merci aussi à Sara, que je n’ai appris à connaître que tard, à mon grand regret. Je pense qu’elle est tout à fait dans « l’esprit CRC » avec sa gentillesse et son calme naturels. Bonne continuation à toi, tant dans cette aventure qu’est la thèse qu’avec ta petite famille. Enfin, je sais bien qu’il est parti vers de nouvelles aventures, mais un grand merci à notre aveyronnais préféré : Arnaud. Merci pour ton caractère si joyeux qui a souvent ensoleillé l’ambiance au 107. Je ne sais pas si je dois te remercier pour le choix des stations de radio, mais ça nous aura valu de belles séances karaoké qui enchantaient tant Alex. En tout cas, bonne chance à toi pour la suite et à très vite j’espère.
Je termine maintenant ce tour du 1er étage par le labo 106 qui a abrité de nombreux locataires. A l’époque de mon DEA, il y avait tout d’abord Michel, notre expert du monde des lipides. Merci pour votre disponibilité, vos nombreux conseils concernant ce monde qui ne nous est pas très familier. Merci aussi pour votre bonne humeur et votre simplicité, j’espère que tout se passe
Remerciements
bien à l’ICR et je vous souhaite bonne continuation pour la suite. Petite Caro, notre « maman thésarde », merci à toi aussi pour ta simplicité, ta gentillesse et pour nos nombreux bavardages (j’ai enfin trouvé quelqu’un qui parle au moins autant que moi, si, si, je vous promets…). Je te souhaite bonne chance pour ta thèse, mais surtout plein de bonheur avec tes petites et ton homme. C’est promis, je t’appelle bientôt pour aller partager un petit café et papoter, ça commence à me manquer… Merci aussi à Julien et Christophe, pour nous avoir souvent fait rire avec Caro et pour tous ces bons moments partagés lors de vos stages. Les murs du 106 abritent maintenant de nouvelles expertes en lipidomique : Justine et Véronique. Merci à toutes les 2 de m’avoir accueillie dans votre bureau lors de cette fin de thèse. Merci pour votre disponibilité et votre bonne humeur. Un merci tout particulier à Véronique pour ses histoires d’araignées qui font froid dans le dos, et à Justine pour ce super pointeur laser le jour de la soutenance, celui-là, on s’en souviendra… Bonne continuation à toutes les 2 dans vos vies professionnelles et personnelles.
Passons maintenant au 2ème étage qui lui aussi abrite des personnalités hautes en couleur !!! ça ne vous étonnera pas si je commence donc par notre petite Camille. Là aussi, j’aurais tant à dire. J’ai su que t’irais loin dès le premier jour où je t’ai vu dans le labo (bien sur, juste après que j’ai compris que t’étais pas la petite sœur de Seb…). Non, plus sérieusement, tu es quelqu’un d’unique et adorable. Merci pour ton côté « atypique » si attachant et qui te va si bien. Merci pour ton caractère enjoué, pour ton soutien de tous les instants (et il y en a eu), pour ta connaissance parfaite des imprimantes et photocopieuses du labo qui a permis que ce manuscrit soit prêt à temps et pour tous ces moments partagés. Je souhaite qu’il y en ait bien d’autres et que notre amitié ne s’arrête pas là (de toute façon, au moins pendant 1 an, tu vas encore devoir me supporter à la fac…). Dernière chose, ne laisses personne te faire douter de toi, tu as tout pour réussir et tu vas le faire, c’est une certitude… Je ne peux pas parler de Camille sans penser à Florence. Sans toi, l’année de DEA aurait été bien différente. C’est ton soutien et ta présence qui m’ont fait tenir à cette époque et je garde en mémoire tous ces excellents moments, de travail mais aussi de fêtes et de complicité passés avec toi. La suite nous a un peu éloigné, tu pars maintenant pour Amsterdam et je suis sûre que tu vas tout réussir là bas pour nous revenir vite. On se retrouvera alors sur Toulouse, autour d’un bon repas avec un bon petit vin pour fêter ça… Concernant les autres thésards du second, je pense que la relève est assurée… Merci à toi Pierre pour ton côté un peu décalé qui a souvent égayé les repas du midi avec l’aide de Muriel et Fabienne. Gardes cette passion de la science qui t’anime et bon courage pour la suite. Bonne chance aussi à Aurélie et Claudia. Merci pour votre simplicité et votre gentillesse. Je vous souhaite tout le meilleur pour la suite. Bienvenu aussi aux nouveaux du thème A, Gérald et Jayati, que je n’ai que peu côtoyés mais à qui je souhaite une excellente réussite professionnelle dans nos murs. Re-bienvenu à Guillaume, qui a finalement décidé de ne pas encore nous quitter. Merci pour ton sourire et ta gentillesse et bonne continuation.
Remerciements
avoir longtemps veillé au bon fonctionnement du laboratoire et sur nous tous. Merci pour ta disponibilité et pour ton caractère qui représente un doux équilibre entre beaucoup de rigueur, d’exigence et un cœur gros comme ça. Un grand merci aussi à Michel, pour sa disponibilité, son calme qui semble résister à toutes les épreuves. Merci pour ta gentillesse et pour avoir bien pris soin de nos papilles gustatives en nous dénichant quelques mets délicieux qui nous ont ravi. On ne peut pas parler de Michel sans évoquer sa compère Christine. Merci à toi pour nos longues discussions et nos fous rires lorsque je me réfugiais dans la réserve… Merci pour ton franc parlé et ta bonne humeur, ainsi que pour tous nos mails échangés… Je te souhaite le meilleur pour la suite. Ça y est, avec le retour de Véro le trio s’est reformé. Tu m’as connu à mes débuts et tu reviens pour mon départ, c’est dommage, mais on aura sûrement d’autres occasions de se croiser. En tout cas, merci pour tes encouragements avant la soutenance, pour ta gentillesse et pour ta soirée de thèse que nous avions tant apprécié avec Flo. Courage pour la suite et je suis sûre que le meilleur reste à venir… Bonne chance à Marie-France pour sa soutenance de thèse et bonne retraite à Marie-Claude qui s’est montrée si disponible lorsqu’il fallait s’occuper des formations ou de la prise en charge des stagiaires. Je souhaite aussi une excellente réussite à Laurent et Stéphane, merci pour votre dynamisme qui anime si bien la vie scientifique du bâtiment C. Merci à Eric pour sa disponibilité et sa gentillesse et pour nous avoir si bien sensibilisés au monde de l’immunologie qui nous paraissait alors si lointain. Je finirais ce tour du thème A par notre fameux trio de l’époque : Xavier, Ronald et François. Merci d’avoir si bien animé la vie du bâtiment C, d’avoir accepté de signer un fameux poster qui restera longtemps dans les annales. Bonne chance à Ronald pour sa nouvelle aventure et merci d’avoir amélioré notre quotidien avec tes compétences informatiques. Merci François pour avoir aidé les petits lutins lors de quelques repas de Noël et d’avoir fait en sorte que tout se passe pour le mieux. Enfin, merci à Xavier pour ta disponibilité et ton implication lors des différents séminaires dès qu’il s’agissait de discuter et de faire avancer les différents sujets. Merci aussi pour ton humour, parfois grinçant, mais qui fait souvent sourire et qui te va si bien.
Ça y est, je descends enfin au rez-de chaussée. Un immense merci tout d’abord à Yvette, notre maman à tous. Merci pour ta disponibilité de tous les moments, y compris lorsqu’on te dérange au milieu de ton repas. Merci pour tout ce que tu fais au quotidien, de si bien veiller sur nous. Le labo ne tournerait pas si bien sans toi, et merci aussi pour toutes tes qualités humaines et ta gentillesse si naturelle. Un merci tout particulier pour avoir si bien reçu ma petite famille et pour les cachotteries que vous avez organisé dans mon dos… Bonne continuation à toi et à toute ta petite famille. Merci à Joël, pour continuer à venir prendre ses repas en notre compagnie et pour souvent endosser le rôle de seul représentant masculin au milieu de toutes ces filles. Mais je te rassure, tu le faisais parfaitement bien… Enfin, un merci tout particulier à Jacques et Nordine, sans qui ce bâtiment ou nos équipements ne tiendraient pas aussi bien le coup. Merci d’accourir si rapidement à notre secours, ainsi que pour la gentillesse avec laquelle vous le faites. Désolée de vous avoir si souvent dérangé… Je finirais ce tour du bâtiment en évoquant des personnes qui ont quitté ces lieux mais qui restent pour moi associés à cette thèse. Merci à Paul pour sa gentillesse et son soutien. Bonne chance à toi et à Carol pour la suite et j’espère qu’on se retrouvera très vite. Un bisou
Remerciements
particulier à Karine. Merci pour ta spontanéité et pour nos nombreux fous rires. Tu es vraiment quelqu’un de bien et j’espère que dès ton retour en France on se retrouvera rapidement autour d’une tasse de café pour tout se raconter. Merci aussi à Seb, Laurence et notre québécois Martin pour ces quelques soirées ou journées au ski passées en votre compagnie. Merci pour votre soutien et votre bonne humeur. Je vous souhaite une bonne continuation et j’espère vous revoir très vite.
Je ne peux pas penser à ma thèse sans parler de l’équipe Payrastre avec laquelle j’ai longuement co-habité à mes débuts… Merci Bernard pour ton recul scientifique et tes conseils avisés. Merci pour ta franchise et ton caractère qui peut parfois impressionner mais qui fais aussi partie de ton personnage. Merci aux autres membres de l’équipe, Fred, Hélène, Claire, Monique,
Marie-Pierre... Merci à toutes pour votre gentillesse, votre disponibilité et vos nombreux conseils.
Bonne chance pour la suite. Merci aussi à Audrey, Sophie et Fabienne pour leur écoute, leur soutien et les bons moments passés ensemble. Merci à Loïc et Damien pour leur bonne humeur et pour égayer régulièrement l’ambiance dans les labos. Bonne continuation à vous 2. Une dédicace toute particulière à Gaëtan et Cédric. Je ne vous remercierais jamais à tous les 2 de m’avoir permis de rencontrer quelqu’un qui représente maintenant un des piliers de ma vie. Je ne l’oublierais pas, promis… Cédric, merci pour ton caractère qui peut parfois déplaire mais qui pour moi est un exemple de franchise. J’espère qu’on continuera à se donner des nouvelles régulièrement et à se voir de temps en temps et quoi qu’il arrive, je suis sûre que tu vas tout déchirer lors de ta thèse.
Gaët, j’aurais là aussi pleins de choses à dire. Merci pour tous ces moments passés ensemble, pour
avoir souvent été mon confident, pour cette franchise qui t’honore et pour ta gentillesse et ta disponibilité de tout instant. Je sais qu’on s’est moins vu ces derniers temps, mais j’espère bien que notre amitié durera encore un moment. Plein de bonheur pour la suite… Dans la même catégorie des gens « à part », je mettrais aussi Sonia et Kro. Merci à toutes les 2 pour votre amitié sans faille. J’ai eu de la chance de vous avoir rencontré… So, je t’admire pour cette décision que tu as prise de partir. Promis, je viens passer un week-end chez toi très vite. En attendant, je pense fort à toi et je suis sûre que tu vas briller pour la suite et concrétiser tout ce dont tu rêves. Tu as toutes les qualités scientifiques et humaines pour ça. Ne restes jamais trop loin car j’ai besoin de ton amitié et ton soutien. Merci pour tout… Kro, merci à toi aussi pour ton soutien, pour toutes nos longues discussions et confidences, pour toutes ces soirées et ces bons moments. Tu es quelqu’un d’exceptionnel et j’espère que notre amitié va encore durer longtemps. On s’est aussi éloigné quelques peu ces derniers temps, mais je compte bien remédier à ça. Que tous tes projets se concrétisent et je te souhaite plein de bonheur avec Romain. Vivement que je vois votre petit nid douillet… Enfin, cette aventure du DEA et de la thèse n’aurait pas eu lieu sans ceux qui m’ont donné envie de continuer dans cette voie. Merci à mon équipe d’accueil de l’ENVT lors de mon stage de maîtrise. Merci à Michèle, notamment pour être venu pour ma thèse, ainsi que pour sa
Remerciements
apporter aux autres. Je ne serais pas arrivée là sans toi. J’espère qu’on restera longtemps en contact, que je puisse voir grandir ton p’tit bout de chou si craquant. Plein de bonheur pour la suite… Merci aussi de m’avoir fait connaître notre petite association qui m’a elle aussi tant apporté et m’a permis de rencontrer des personnalités si exceptionnelles. Un gros bisou particulier à mes « chouchous » :
Christian, Marie-Laure, Marie la brune, Emmanuel, Guy et Jeff. Je sais que je vous ai dit au
revoir il y a 2 ans maintenant, mais je vous promets que ce n’était pas un adieu et que j’espère vous retrouver très vite. Merci à chacun d’entre vous pour vos caractères si différents mais qui m’ont tant apporté, chacun à leur façon…
Je sais, vous allez finir par dire que mes remerciements sont à la hauteur de ce manuscrit, c'est-à-dire très longs… Ne vous inquiétez pas, il ne me reste maintenant que les « non initiés » à la vie de labo, mais qui pourtant comptent tant pour moi. Je commencerais par mes ami(e)s. Tout d’abord, les filles du Gers, eh oui, déjà 11 ans d’amitié et 11 ans qu’on se supporte depuis ces folles années d’internat… Merci à Audrey, Flo qui a maintenant son Nico, Nanou, Amé et Chéri-Chéri,
Caille et Aurélie. Merci à chacune pour votre amitié fidèle. Vous avez toutes contribué à cette
réussite, en m’apportant soutien, écoute et compréhension quand j’en avais besoin et pleins de fous rires et d’excellents moments quand il fallait que je décompresse. Que cette amitié dure encore longtemps… Il y a aussi 2 amies d’enfance chères à mon cœur que je voulais remercier ici : Paps, toi ma plus « vieille » amie et Delf, ma petite sœur d’adoption. Vous voilà toutes 2 maintenant mariées et je vous souhaite pleins de bonheur, en espérant partager encore bien longtemps des moments privilégiés avec vous. Après les filles, je passe logiquement aux mecs. Merci au trio infernal : Fred, Mika et Juju, ainsi qu’à vos moitiés respectives : Nath, Brigitte et Mag. Merci à tous pour tous ces moments partagés, toutes ces fêtes et ces voyages remplis de tant de souvenirs (ah, l’Italie !!!). Je vous souhaite pleins de bonheur et à très bientôt pour fêter ça !!! Spécial grand merci aux footballeurs fous : La Boosk, Tchitche, Jéjé et Le Maz. Merci à tous les 4 pour votre amitié et pour me voler mon homme presque 1 samedi soir sur 2… J’espère passer encore pleins de bons moments avec vous... Je ne peux pas évoquer le foot sans penser à Schies et à Ced. Merci à tous les 2 de m’avoir si rapidement adoptée et pour cette si belle amitié qui vous lie à Olivier. Je sais qu’on s’est peu vu ces derniers temps, mais promis, on va remédier à ça, et l’avenir nous promet encore de bons moments… Merci à Sandy, je suis heureuse qu’on se soit rapprochée ces dernières années, et vu qu’on est bientôt voisine, j’espère que cette amitié durera encore longtemps. En parlant de voisins, merci aussi à Caro et Lolo, qui seront bientôt 3 et à qui je souhaite le meilleur pour la suite. Vous savez qu’on est là pour le baby sitting quand vous voulez, le package Bébé et Titi est valable pour une longue durée… Pour finir avec ce groupe, un bisou tout particulier à Bouddha et Yana que j’ai plaisir à retrouver à chaque fois malgré la distance et à Coco pour sa gentillesse et son caractère si attachant. Promis, on viendra te voir à Sète, et tu passes quand tu veux à la maison, la porte est ouverte et les bières sont au frais… Enfin, c’est un peu particulier, mais merci aux « vieux », les amis de mes parents mais pourtant si proches de moi et toujours prêts à me soutenir : les Buffomène, avec bien entendu Magalie, Eric et Eugénie et Carole, David et Axel, les Faget, les Brossard, les Larroche, les Lampin avec une pensée particulière pour
Jean-Remerciements Jacques qui nous manque, mais qui je suis sûre a tout vu de là-haut et est associé à part entière à ce
bonheur et cette réussite, les Comet et les Dupin.
J’en ai fini avec les amis et il me reste maintenant la famille. Merci à mon Papa, ma
Maman, mon petit frère Juju et sa moitié Anne-Cé. Merci à tous pour ce que vous êtes, pour
toujours croire en moi et pour tout l’amour que vous me donnez. Cette réussite est aussi la vôtre, je ne serais pas allée bien loin sans vous… Merci d’avoir fait de moi ce que je suis maintenant… Merci à mes grands-mères, Marcelle ma super Mamie et Jeanine qui me gâte tant. Merci à toutes les 2 pour votre amour et vos petites attentions. Papi Pierre, Papi Georges et Jean-Michel auraient sûrement été fiers, et cette thèse est aussi pour eux… Merci à ma Tatie et mon Tonton qui ont toujours été là pour moi. Merci pour votre soutien et votre écoute. Ne restez jamais trop loin de moi, vous faites partie de mon équilibre… Un énorme bisou et un énorme merci à ma cousine,
Marianne. Tu l’as dit un jour, on est unie par le sang, mais aussi par le cœur. On ne peut pas lutter
contre ça, et merci à toi pour ce que tu es. Sans toi, je ne serais pas non plus allée bien loin. La prochaine étape, c’est le mariage, il me tarde déjà de te voir toute en blanc… Je te souhaite en tout cas de longues années de bonheur avec Nico. Merci aussi à Alice, Jeanne, Annie, Jean-Louis et les
Cordelier d’être toujours là pour moi, tant pour me soutenir que pour passer de bons moments
ensemble.
Je passe maintenant à ma deuxième famille. Merci à Nicole, Marc et Manu. Merci pour tout cet amour que vous m’apportez, pour votre soutien, pour m’avoir fait confiance, et pour avoir fait de cette thèse une réussite. Si, si, je vous assure, c’est en partie grâce à vous. Il nous reste encore pleins de bons moments à vivre ensemble et il me tarde déjà… Un grand merci aussi aux
familles Maître et Noguer au sens large, c'est-à-dire les grands parents et les générations qui
suivent : tous les oncles, tantes et cousin(e)s. Merci de m’avoir adoptée si rapidement, pour vos encouragements de tout instant, ainsi que pour votre gentillesse, vos valeurs et vos qualités humaines à tous. Je suis heureuse de vous avoir rencontré et je vous dédie aussi cette thèse. Un merci tout particulier aux « vieux » cousins : Fabienne et Laurent, Cyril et Céline et Stéphane. Merci pour toutes nos discussions, ces moments passés ensemble, ces soirées et ces fous rires. J’espère qu’il y en aura bien d’autres…
Enfin, et ceci n’étonnera personne, mais mes derniers mots seront pour Olivier. Tu es l’une des plus belles choses qui me soient arrivées. Je ne te remercierais jamais assez pour ta persévérance à nos débuts qui a permis que nous ayons déjà parcouru tout ce chemin ensemble. Tu sais tout ce que tu représentes pour moi, tu es à la fois mon pilier, mon équilibre et ma moitié. Merci pour ton soutien sans faille, ton amour, ta patience, ta gentillesse. Merci de me redonner confiance dans les moments de doutes. Ne t’éloignes jamais trop de moi. Que ces moments passés avec toi ne cessent jamais, et je sais qu’il nous reste encore beaucoup de projets à concrétiser. Cr…’
Sommaire
ABREVIATIONS...4
RESUME ...6
ABSTRACT ...7
LISTE DES FIGURES ET TABLEAUX ...8
INTRODUCTION GENERALE ...10
PARTIE I : INTRODUCTION BIBLIOGRAPHIQUE ... 13
CHAPITRE 1 : LES RTK, STRUCTURE ET MECANISMES D’ACTIVATION : EXEMPLES DE L’EGFR ET DU VEGFR ...14
I- Généralités sur les RTK ... 14
I-1. Introduction ... 14
I-2. Fonctions biologiques des RTK ... 15
I-3. Pouvoir oncogène des RTK... 16
I-4. Structure des RTK... 16
I-5. Dimérisation et activation des RTK ... 18
II- EGFR et famille des récepteurs ErbB ... 19
II-1. Introduction... 19
II-2. Les récepteurs ErbB et leurs ligands ... 19
II-3. Fonctions biologiques des récepteurs ErbB ... 21
II-4. Structure, dimérisation et activation des récepteurs ErbB... 22
II-5. Activation des récepteurs ErbB indépendante du ligand... 25
III- VEGFR ... 27
III-1. Introduction ... 27
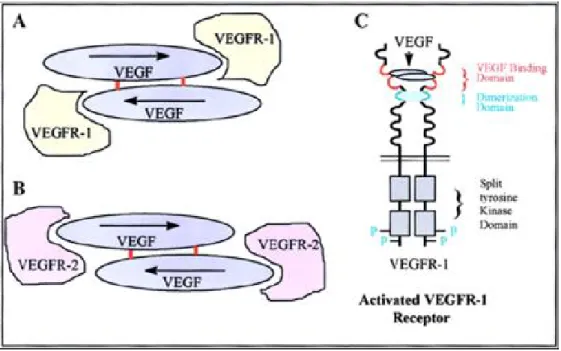
III-2. Présentation des VEGFR et de leurs ligands ... 27
III-2-1. Présentation des ligands de la famille du VEGF ... 28
III-2-2. Présentation des VEGFR ... 30
III-3. Fonctions biologiques des VEGFR... 32
III-4. Structure, dimérisation et activation des VEGFR... 33
III-5. Activation des VEGFR indépendante du ligand ... 35
IV- Régulation des RTK... 36
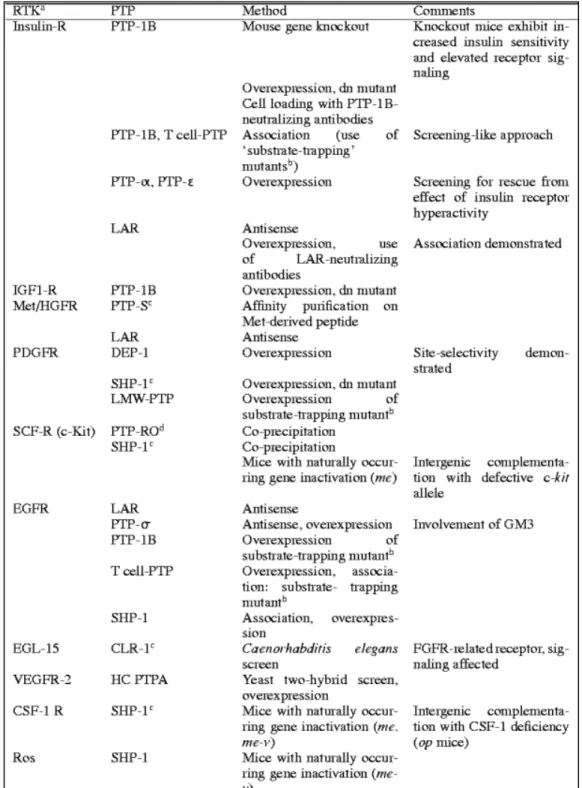
IV-1. Régulation par le récepteur lui-même... 37
IV-2. Régulation par des protéines tyrosines phosphatases (PTP)... 37
IV-3. Régulation par endocytose ... 39
V- Conclusion... 42
CHAPITRE 2 : MECANISMES D’INITIATION ET D’ACTIVATION DES VOIES RAS/MAPK ET PI3K PAR L’EGFR ET LE VEGFR ...43
I- Mécanismes d’initiation et d’assemblage des voies de signalisation par les RTK : exemples de l’EGFR et du VEGFR ... 43
I-1. Initiation des voies de signalisation par recrutement des protéines à domaines SH2 et PTB... 43
I-2. Propagation du signal par assemblage des complexes de signalisation grâce à divers domaines protéiques d’interaction ... 44
I-3. Les protéines adaptatrices : protéines indispensables à la formation des complexes de signalisation ... 46
Sommaire
I-3-2. Les protéines Gab (Grb2-associated binder) ... 49
I-4. Voies de signalisation activées par l’EGFR et les récepteurs ErbB ... 52
I-5. Signalisation induite par les VEGFR ... 54
I-5-1. Signalisation induite par le VEGFR-1... 54
I-5-2. Signalisation induite par le VEGFR-2... 56
I-5-3. Signalisation induite par le VEGFR-3... 58
II- La voie des MAPK en aval de l’EGFR et du VEGFR... 59
II-1. La voie canonique des Ras/MAPK ... 59
II-1-1. Ras : protéine centrale dans l’activation des MAPK... 59
a) la superfamille des Ras... 59
b) présentation de Ras et de ses mécanismes d’activation... 61
c) inhibiteurs de Ras ... 62
d) effecteurs de Ras ... 63
II-1-2. Les MAPK ... 64
a) Raf ... 64
b) le module MEK1-2/Erk1-2 ... 66
c) les substrats des Erk1-2 ... 66
d) mise en place et régulation de la voie des MAPK... 67
II-2. Gab1 et SHP-2 : deux nouveaux acteurs de la voie des MAPK en aval de l’EGFR ... 70
II-3. Cas du VEGFR-2 : présentation d’un modèle original d’activation des MAPK... 70
II-3-1. Mécanisme d’activation des MAPK en aval du VEGFR-2... 70
II-3-2. Fonctions biologiques des MAPK dans les cellules endothéliales... 72
II-4. Conclusion ... 72
III- La voie des PI3K en aval de l’EGFR et du VEGFR ... 72
III-1. La famille des PI3K ... 73
III-2. Mécanismes d’activation des PI3K... 75
III-3. Les 3-phosphoinositides ... 77
III-3-1. Le phosphatidylinositol 3-phosphate (PI3P)... 78
III-3-2. Le phosphatidylinositol 3,4-bisphosphate (PI3,4P2)... 78
III-3-3. Le phosphatidylinositol 3,5-bisphosphate (PI3,5P2)... 79
III-3-4. Le phosphatidylinositol 3,4,5-trisphosphate (PIP3) ... 79
III-4. Effecteurs de PI3K... 79
III-4-1. La sérine/thréonine kinase Akt/PKB (protéine kinase B) ... 80
a) Présentation de la famille des kinases Akt et de leurs mécanismes d’activation :... 80
b) Fonctions cellulaires régulées par PI3K par l’intermédiaire d’Akt :... 81
III-4-2. Les GEF des petites protéines G... 83
III-4-3. Autres ... 83
III-5. Régulation de PI3K ... 84
III-5-1. Les phosphatases PTEN (phosphatase and tensin homolog deleted on chromosom 10) et SHIP (SH2-containing inositol phosphatase): ... 84
III-5-2. Par phosphorylation de p85 : ... 85
III-5-3. Par régulation de Akt :... 85
III-6. SHP-2 et Gab1 dans l’activation de PI3K en réponse à l’EGFR ... 86
III-7. La voie PI3K en aval du VEGFR-2 ... 86
III-7-1. Mécanismes d’activation de PI3K en aval du VEGFR-2 ... 86
III-7-2. Fonctions biologiques de la voie PI3K dans les cellules endothéliales ... 87
III-8. Conclusion ... 88
IV- Cross talk entre les voies Ras/MAPK et PI3K... 88
IV-1. Régulation de la voie PI3K par la voie Ras/MAPK ... 88
IV-2. Régulation de la voie Ras/MAPK par la voie PI3K ... 89
IV-3. Conclusion... 90
Sommaire
I-1-3. Thérapies anti-cancéreuses ciblant MEK ... 94
I-2. Altérations de la voie PI3K et cancer ... 96
II- ErbB et cancers... 99
II-1. Les ErbB dans les tumeurs solides... 99
II-2. Inhibiteurs des ErbB en thérapie anti-cancéreuse ... 100
II-2-1. Les anticorps bloquants... 101
II-2-2. Les inhibiteurs de l’activité kinase (TKI ou Tyrosine Kinase Inhibitor) ... 103
II-2-3. Autres inhibiteurs et échecs thérapeutiques ... 104
III- le VEGFR ... 105
III-1. Les VEGFR et leurs ligands dans l’angiogenèse ... 105
III-2. Inhibiteurs du VEGFR en thérapie ... 107
III-2-1. Les anticorps bloquants ... 107
III-2-2. Les TKI... 108
IV- Intérêt des combinaison anti-EGFR et anti-VEGFR en thérapie anti-cancéreuse. ... 109
IV-1. Liens entre EGFR et VEGFR et efficacité de la double inhibition... 109
IV-2. Mécanismes de la double inhibition ... 110
IV-3. Conclusion... 111
PARTIE II : RESULTATS EXPERIMENTAUX ET PERSPECTIVES ... 112
RESULTATS EXPERIMENTAUX ...113
I- Article 1 : Rôle de la protéine adaptatrice Gab1 dans l’activation de PI3K en aval du VEGFR-2 ... 113
I-1. Introduction ... 113
I-2. Résultats : article 1 ... 113
I-3. Conclusion... 125
II- Article 2 : La dépendance de la voie Ras/MAPK vis-à-vis de PI3K est conditionnée par le mode de recrutement du module Gab1/SHP-2... 127
II-1. Introduction... 127
II-2. Résultats : article 2... 128
II-3. Conclusion ... 166
PERSPECTIVES ...168
I- Identification du rôle précis de Gab1 en aval du VEGFR-2 ... 168
I-1. Détermination du site d’interaction de Gab1 sur le récepteur et de la kinase responsable de sa phosphorylation ... 168
I-2. Détermination des cibles du module Gab1/PI3K impliquées dans la migration des cellules endothéliales et/ou dans le processus d’angiogenèse... 169
I-3. Détermination de l’importance de Gab1 dans la migration des cellules endothéliales et le processus d’angiogenèse ... 169
I-4. Gab1 dans l’activation des MAPK en aval du VEGFR-2 ?... 170
II- Participation du mode de recrutement du module Gab1/SHP-2 dans la résistance aux inhibiteurs de l’EGFR ... 171
III- Conclusion ... 172
PARTIE III : BIBLIOGRAPHIE ... 173
Abréviations
ABREVIATIONS
3 - PI 3 - Phosphoinositide
ADN Acide Désoxyribonucléique
ADP Adenosine Diphosphate
AMP Adenosine Monophosphate
ARN Acide ribonucléique
ATP Adenosine Triphosphate
DAG Diacylglycérol
Dos Daughter of Sevenless
EGF Epidermal Growth Factor
EGFR EGF Receptor
ERK 1 - 2 Extracellular-regulated kinase 1 - 2
FAK Focal Adhesion Kinase
FGF fibroblast Growth Factor
Flk Fetal Liver Kinase
Flt Fms-Like Tyrosine kinase
FTI Farnesyl Transferase Inhibitor
Gab1 Grb2-associated Binder 1
GAP GTPase Activating Protein
GDI Guanosine nucleotide Dissociation Inhibitor
GDP Guanosine Diphosphate
GEF Guanosine Exchange Factor
GTP Guanosine Triphosphate
HB-EGF Heparin Binding EGF-like factor
HGF Hepatocyte Growth Factor
hsp Heat Shock Protein
HUVEC Human umbilical Vein endothelial Cells
Ig Immunoglobulin
InsR ou IR Insulin Receptor
IP3 Inositol 1,4,5-trisphosphate
IRS Insulin Receptor Substrate
JNK c-Jun Terminal Kinase
KDR Kinase insert Domain Receptor
MAPK Mitogen-Activated Protein Kinase
MAPKAPK ou MPK MAPK-activated Protein Kinase
MBD Met Binding Domain
MEK MAPK Erk Kinase
MEKK MEK Kinase
MKP MAPK Phosphatase
Abréviations
NOS Nitric Oxyde Synthase
NRG Neuréguline
PDGF Platelet-Derived Growth Factor
PDK Phosphoinositide Dependent Kinase
PH Pleckstrin Homology
PI3,4P2 Phosphatidylinositol 3,4-Bisphosphate
PI3,5P2 Phosphatidylinositol 3,5-Bisphosphate
PI3K Phosphoinositide 3-Kinase
PI Phosphoinositide
PI3P Phosphatidylinositol 3-Phosphate
PI4,5P2 Phosphatidylinositol 4,5-Bisphosphate
PI4K Phosphoinositol 4-Kinase
PI4P Phosphatidylinositol 4-Phosphate
PI5K Phosphoinositol 5-Kinase
PI5P Phosphatidylinositol 5-Phosphate
PIP3 Phosphatidylinositol 3,4,5-Trisphosphate
PKA, B, C, D Proteine Kinase A, B, C, D
PLC Phospholipase C
PLD Phospholipase D
PlGF Placenta Growth Factor
PRD Prolin Rich Domain
PTB Phosphotyrosine Binding
PTEN Phosphatase and Tensin Homolog Deleted on chromosome 10
PTK Protein Tyrosine Kinase
PTP Protein Tyrosine Phosphatase
RBD Ras Binding Domain
RCPG Récepteur couplés aux Protéines G
RTK Récepteur à activité Tyrosine Kinase
SH2 Src Homology 2
SH3 Src Homology 3
SHIP SH2-containing Inositol Phosphatase
SHP SH2 domain -containing tyrosine phosphatase
sos Son Of Sevenless
STAT Signal Transducers and Activators of Transcription
TGF Transforming Growth Factor
TKI Tyrosine Kinase Inhibitor
Résumé
RESUME
Nos travaux se sont centrés sur l’étude des voies de signalisation mises en place par la cellule en aval de deux récepteurs à activité tyrosine kinase intrinsèque (RTK), le récepteur de l’EGF (Epidermal Growth Factor) et le récepteur du VEGF (Vascular Endothelial Growth Factor). Deux voies de signalisations semblent prépondérantes dans l’activité biologique du VEGFR et de l’EGFR : la voie PI3K (phosphoinositide 3-kinase) et la voie Ras/MAPK (Mitogen-Activated Protein Kinase). Non seulement ces récepteurs et les signalisations associées contrôlent de nombreux processus cellulaires fondamentaux essentiels à l’homéostasie tissulaire mais se retrouvent aussi respectivement impliqués dans un grand nombre de tumeurs solides et dans le processus d’angiogenèse tumorale. Ainsi, comprendre la signalisation induite par ces 2 récepteurs permettrait sûrement d’identifier de nouvelles cibles pouvant être utilisée en thérapie anti-cancéreuse.
Dans un premier temps, nous avons montré que la protéine adaptatrice Gab1 était phosphorylée en réponse au VEGF, s’associait alors à PI3K et jouait un rôle majeur dans son activation. Nous avons donc permis l’identification de Gab1 comme étant un relais essentiel entre le VEGFR et l’activation de PI3K, lien qui était jusqu’à présent mal connu. Nos résultats suggèrent de plus que ce module VEGFR/Gab1/PI3K pourrait jouer un rôle majeur dans la migration des cellules endothéliales, processus généralement associé à l’angiogenèse tumorale. Dans un deuxième temps, nous nous sommes intéressés au rôle de PI3K dans l’activation de la voie Ras/MAPK en réponse à l’EGF. En effet, il a été montré que PI3K pouvait intervenir ou pas dans l’activation de Ras/MAPK, sans que d’explications à ces différences aient été fournies. Nous avons montré que la dépendance de la voie Ras/MAPK vis-à-vis de PI3K en réponse à l’EGF observée dans divers modèle cellulaire était en fait conditionnée par le mode de recrutement de Gab1 et de la phosphatase SHP-2 (SH2 domain-containing tyrosine phosphatase), indispensables à l’activation des MAPK en aval de la plupart des RTK. Nos résultats montrent que, lorsque beaucoup de molécules d’EGFR sont mobilisées, le module Gab1/SHP2 est recruté auprès du récepteur par l’intermédiaire de la protéine adaptatrice Grb2. Cependant, lorsque peu de molécules d’EGFR sont engagées, le module Gab1/SHP2 est recruté grâce au domaine PH de Gab1 ayant une affinité particulière pour les produits de PI3K, permettant ainsi le recrutement de molécules additionnelles de Gab1 et donc une activation efficace des MAPK. De plus, nous avons remarqué que ce mécanisme de recrutement de Gab1/SHP-2 pouvait être mis en place dans des cellules tumorales traitées par des inhibiteurs de l’EGFR, notamment, des lignées de glioblastomes, permettant donc une activation de la voie Ras/MAPK malgré l’inhibition de l’EGFR. Ainsi, la mise en jeu de ce mécanisme compensatoire de recrutement de Gab1 pourrait expliquer le phénomène de résistance aux traitements anti-EGFR souvent observé dans ces tumeurs.
Abstract
ABSTRACT
This thesis work has been devoted to the study of signaling pathways mobilized by the cell downstream of two receptors tyrosine kinase (RTK), namely the EGF (Epidermal Growth Factor) receptor and the VEGF (Vascular Endothelial Growth Factor) receptor. Two paramount signaling pathways mediate the biological activity of VEGFR and EGFR: the PI3K (phosphoinositide 3-kinase) and the Ras/MAPK (Mitogen-Activated Protein Kinase) pathways. Not only are these receptors and associated signalings controlling many fundamental cellular processes that are necessary to maintain cellular homeostasy, but they are respectively involved in solid tumor development and tumor angiogenesis. Thus, understanding the signaling pathways induced by these two receptors would certainly provide new insight into the identification targets for anticancer therapies.
Firstly, we have demonstrated that the adaptor protein Gab1 is phosphorylated in response to VEGF, binds to PI3K and plays then a key role in PI3K activation. This study has thus identified Gab1 as an essential relay between VEGFR and PI3K activation, link that was yet poorly understood. Moreover, our results suggest that this VEGFR/Gab1/PI3K module could critically participate in endothelial cells migration, a process that is in general associated with tumor angiogenesis. Secondly, we have been interested in the role of PI3K in the activation of the Ras/MAPK pathway in response to EGF. Indeed, it has been shown that Ras/MAPK activation can rely, or not, on PI3K activity, but the molecular explanation for these differences have not been described so far. We have demonstrated that the dependency of the Ras/MAPK pathway towards PI3K was in fact conditioned by the mechanism of recruitment of Gab1 and SHP-2 (SH2 domain-containing tyrosine phosphatase), two key actors of the MAPK pathway downstream of a wide range of RTK. Our results show that, when a lot of EGFR molecules are mobilized, the Gab1/SHP2 module is recruited through Grb2. Conversely, when a few EGFR are engaged, the Gab1/SHP2 module is recruited through the Gab1 PH domain, which displays high affinity for PI3K lipid products, thereby allowing the mobilization of additional Gab1 molecules and thus promoting efficient MAPK activation. Moreover, we have observed that this recruitment of Gab1/SHP2 could be implemented in tumor cells treated with EGFR inhibitors, leading to full MAPK activation despite EGFR inhibition. Thus, this compensatory mechanism for Gab1 recruitment could explain the resistance to anti-EGFR treatment, which is often observed in those tumors.
Altogether, these results have identified Gab1 as a key protein in the activation of signaling pathways induced by EGFR and VEGFR. Given its involvement in EGFR-dependent tumor cells proliferation, as well as VEGF-dependent tumor-associated angiogenesis, we assume that Gab1 might be a potential target for directed anticancer therapies.
Liste des figures et tableaux
LISTE DES FIGURES ET TABLEAUX
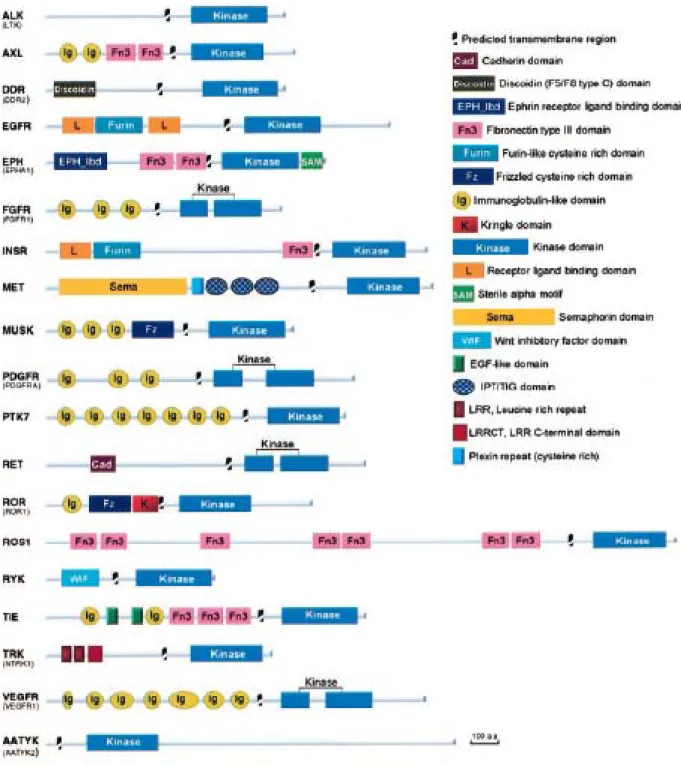
Figures :- Figure 1 : Structure des principales familles de RTK p17
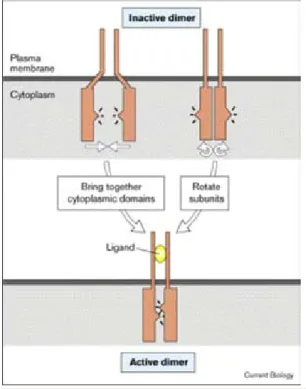
- Figure 2 : Modèle d’activation des dimères de RTK par fixation du ligand p18
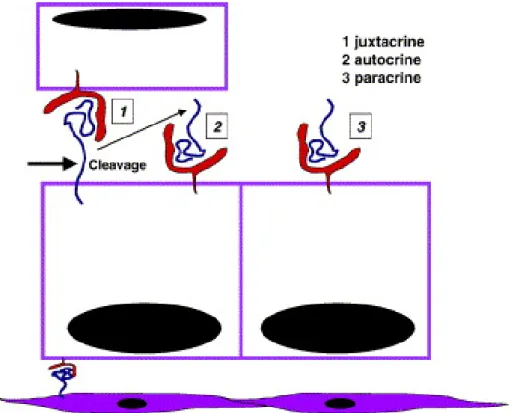
- Figure 3 : Modes d’activation juxtacrine, autocrine et paracrine des récepteurs
ErbB p21
- Figure 4 : Représentation schématique de l’EGFR p23
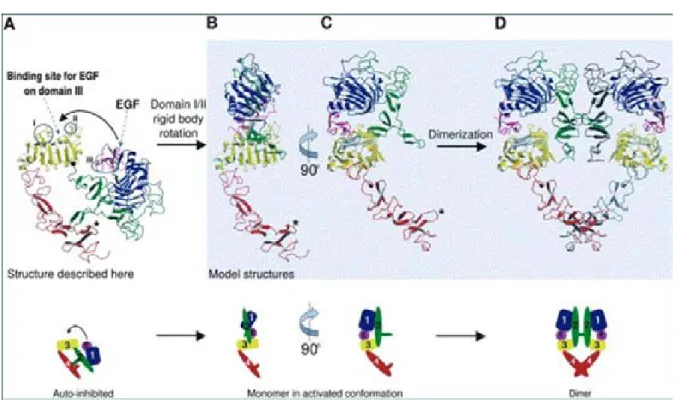
- Figure 5 : Modèle de changement de conformation de la partie
extracellulaire de l’EGFR après fixation du ligand p24
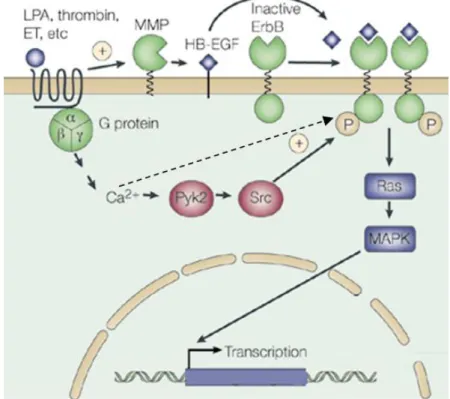
- Figure 6 : Mécanismes de transactivation de l’EGFR par les RCPG p26
- Figure 7 : Structure des principaux VEGF p28
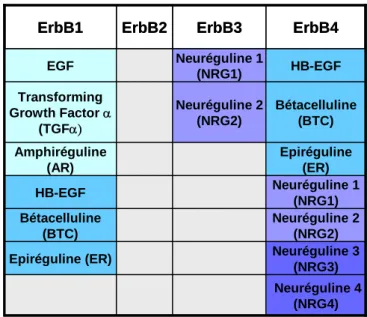
- Figure 8 : Résumé des différentes affinités entre les VEGFR et leurs ligands
et rappel de leurs principales localisations p31
- Figure 9 : Modèle illustrant la fixation du VEGF sur ses récepteurs p34
- Figure 10 : Modèle d’endocytose régulée des RTK p41
- Figure 11 : Mécanismes de régulation des protéines adaptatrices p48
- Figure 12 : Structure des protéines Gab p50
- Figure 13 : Profil de phosphorylation des ErbB p53
- Figure 14 : Profil de phosphorylation du VEGFR-1 p55
- Figure 15 : Profil de phosphorylation du VEGFR-2 p56
- Figure 16 : Profil de phosphorylation du VEGFR-3 p58
- Figure 17 : Mécanismes d’activation des GTPase de la famille des petites
protéines G p60
- Figure 18 : Mécanismes d’activation des effecteurs de Ras p63
- Figure 19 : Schéma général de la voie des MAPK p65
- Figure 20 : Schéma général de l’assemblage de la voie des MAPK p68
- Figure 21 : Résumé des principales protéines d’ancrage et d’assemblage
de la voie des MAPK et de ses inhibiteurs en aval des RTK p69
- Figure 22 : Activation des MAPK en aval du VEGFR-2 p71
- Figure 23 : Les différentes classes de PI3K p74
- Figure 24 : Réactions catalysées par les PI3K in vitro p75
- Figure 25A : Structure des différents phosphoinositides p77
- Figure 25B : mécanismes de synthèse des 3-phosphoinositides p78
- Figure 26 : Principales voies de signalisation activées par Akt et
impliquées dans la prolifération et la survie p82
- Figure 27 : résumé des différents effecteurs de PI3K p84
- Figure 28 : Schéma général de la voie des PI3K p85
- Figure 29 : Gab1 et SHP-2 dans l’activation des voies MAPK et PI3K
en réponse à l’EGF p91
- Figure 30 : Caractéristiques des différentes catégories d’anticorps utilisés
en thérapie p101
- Figure 31 : Effet du VEGF et de ses récepteurs sur le processus d’angiogenèse p107 - Figure 32 : Schéma illustrant l’intérêt de la double thérapie anti-EGFR
Liste des figures et tableaux Tableaux :
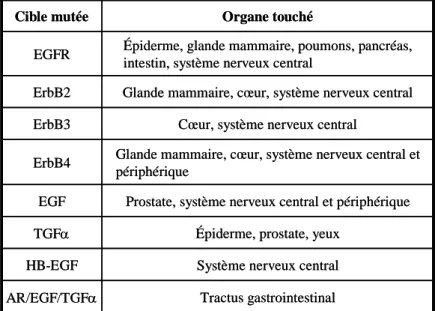
- Tableau 1 : Les ligands de la famille de l’EGF et leurs récepteurs ErbB p20
- Tableau 2 : Conséquences biologiques des mutations des ErbB ou de
leurs ligands p22
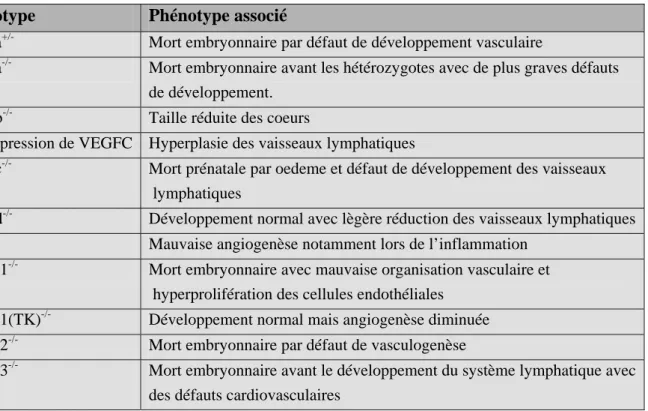
- Tableau 3 : Résumé des phénotypes associés aux différents mutants
ou KO des VEGFR et de leurs ligands p32
- Tableau 4 : Résumé des différentes phosphatases identifiées
comme régulant l’activité des RTK p38
- Tableau 5 : Liste des principaux domaines d’interaction des protéines
nécessaires pour l’initiation et l’assemblage des voies de signalisation p45
- Tableau 6 : Liste des principales associations retrouvées sur les protéines adaptatrices
des domaines SH2 ou PTB avec d’autres domaines protéiques d’interaction p47
- Tableau 7 : Liste des principaux domaines protéiques d’affinité
pour les phosphoinositides p80
- Tableau 8 : Liste des principaux composés ciblant la voie Ras/MAPK
en thérapie anti-cancéreuse p95
- Tableau 9 : Principales altérations de la voie PI3K retrouvées dans les cancers p96 - Tableau 10 : Liste des principaux composés ciblant la voie PI3K en
thérapie anti-cancéreuse p97
- Tableau 11 : Liste des principales altérations des récepteurs de la
famille ErbB retrouvées dans les cancers p99
- Tableau 12 : Liste des principaux anticorps anti-ErbB testés en
thérapie anti-cancéreuse p102
- Tableau 13 : Liste des principaux TKI anti-ErbB testés en thérapie
anti-cancéreuse p104
- Tableau 14 : Régulateurs de l’expression du VEGF dans les cellules tumorales p106 - Tableau 15 : Anticorps anti-VEGFR ou anti-VEGF utilisés en
thérapie anti-cancéreuse p108
Introduction générale
INTRODUCTION GENERALE
Ce travail de thèse s’inscrit dans le cadre de la compréhension des voies de signalisation qui permettent à la cellule d’adapter son comportement à son environnement, afin d’identifier les différents acteurs cellulaires participant aux divers processus biologiques. Ces voies sont généralement des cascades enzymatiques activées en aval de récepteurs présents à la surface cellulaire, en réponse à divers agonistes. Ainsi, en fonction du stimulus, la cellule va pouvoir initier ces voies de signalisation qui vont généralement aboutir à l’activation, dans le noyau, de la transcription de gènes spécifiques permettant d’induire un processus biologique adapté tel que la prolifération, la différenciation ou la survie. D’un point de vue physiopathologique, il est maintenant clairement admis que des altérations ou des dérégulations d’acteurs de ces voies de signalisation sont retrouvées dans de nombreuses maladies telles que les maladies du métabolisme, du développement, ou encore les cancers.
Notre laboratoire étudie plus particulièrement les voies de signalisation induites an aval des récepteurs aux facteurs de croissance autrement appelés récepteurs à activité tyrosine kinase intrinsèque ou RTK. De façon générale, la plupart de ces récepteurs sont indispensables au bon développement de l’organisme, mais aussi au maintien de son intégrité chez l’adulte. Dans un premier chapitre de l’introduction bibliographique, nous décrirons donc les caractéristiques générales de ces RTK, notamment leur structure, leurs mécanismes de dimérisation et d’activation ainsi que leur régulation. Nous nous intéresserons plus spécifiquement à deux RTK particulièrement étudiés au laboratoire : le récepteur de l’EGF (Epidermal Growth factor) ou EGFR et le récepteur du VEGF (Vascular endothelial growth factor) ou VEGFR. Ces deux récepteurs jouent respectivement un rôle majeur dans la mise en place de nombreux tissus et organes lors du développement et dans la formation du système cardiovasculaire.
Le deuxième chapitre de cette introduction bibliographique décrira, quant à lui, les voies de signalisation induites par ces récepteurs et notamment deux voies majeures : la voie des PI3K (phosphoinositide 3-kinase) et la voie des Ras/MAPK (Mitogen-activated Protein kinase). Nous verrons dans un premier temps les mécanismes généraux d’initiation et d’assemblage des voies de signalisation grâce aux différents domaines protéiques d’interaction présents sur les principaux acteurs de ces voies et notamment les protéines adaptatrices. Ensuite, nous décrirons plus précisément les voies Ras/MAPK et PI3K. Pour chacune d’entre elles, nous présenterons le modèle « canonique » d’assemblage et d’activation de ces voies, de même que leurs effecteurs et leurs régulateurs. Nous détaillerons aussi plus particulièrement leur activation en aval de nos deux récepteurs d’intérêt. Concernant l’EGFR, nous illustrerons notamment l’importance de la protéine
Introduction générale
mécanismes d’activation de la voie PI3K en réponse à ce récepteur, qui demeurent assez mal compris. Nous finirons ce chapitre en décrivant les différentes interconnexions décrites dans la littérature entre les voies MAPK et PI3K, et plus particulièrement les acteurs principaux identifiés dans ces mécanismes.
Enfin, nous clôturerons cette introduction bibliographique par un troisième chapitre illustrant l’implication physiopathologique de ces voies et de ces récepteurs, notamment dans les pathologies cancéreuses. En effet, nous verrons rapidement dans une première partie que les voies MAPK et/ou PI3K et leurs acteurs principaux sont souvent retrouvées hyperactivées dans un grand nombre de tumeurs faisant d’elles de potentielles cibles thérapeutiques. Nous nous intéresserons ensuite respectivement aux dérégulations de l’EGFR et au rôle du VEGFR dans le développement tumoral et dans l’angiogenèse qui lui est associée. Nous décrirons les composés ciblant ces deux récepteurs qui sont en cours de développement et nous discuterons des échecs thérapeutiques rencontrés lors des traitements de tumeurs par des inhibiteurs de l’EGFR, et de l’intérêt d’inhiber à la fois l’EGFR et le VEGFR en thérapie anti-cancéreuse.
Notre travail porte donc sur les mécanismes d’activation des voies MAPK et PI3K. Lors d’un premier travail, nous nous sommes tout d’abord intéressés aux mécanismes exacts d’activation de la PI3K en aval du VEGFR-2. En effet, même si cette voie est bien décrite en aval de la plupart des RTK, les données concernant le VEGFR demeurent confuses. Après avoir été longtemps controversé, il est maintenant acquis que la PI3K joue un rôle majeur dans les effets biologiques induits par le VEGF. Cependant, le VEGFR-2 ne possède pas de site consensus de liaison à PI3K suggérant donc l’intervention d’une protéine adaptatrice. Notre intérêt s’est alors porté sur Gab1, une protéine adaptatrice largement décrite pour son rôle dans l’activation de PI3K en aval des RTK, et nous avons démontré que Gab1 était recrutée à la membrane et phosphorylée en réponse au VEGF. De plus, par des stratégies utilisant un dominant négatif de la protéine ne pouvant plus lier la PI3K, nous avons identifié Gab1 comme un relais essentiel à l’activation de PI3K en aval du VEGFR-2. Nos résultats suggèrent de même que Gab1 pourrait jouer un rôle majeur dans la migration des cellules endothéliales dépendante de PI3K, migration qui est notamment impliquée dans le processus d’angiogenèse tumorale. Ces travaux sont présentés dans la première partie des résultats expérimentaux.
Dans un deuxième temps, nous nous sommes intéressés aux interconnexions décrites dans la littérature entre la voie Ras/MAPK et la voie PI3K. En effet, il est maintenant admis que dans certains modèles de cellules « saines » ou cancéreuses et dans certaines conditions, l’activation de la voie Ras/MAPK pouvait être dépendante de la voie PI3K en réponse à l’EGF. Cependant, cette observation semblait dépendre de la force du signal ou du type cellulaire considéré, et le mécanisme moléculaire permettant d’expliquer cette dépendance n’avait toujours pas été identifié. Nous avons donc réussi à démontrer que la dépendance de la voie Ras/MAPK vis-à-vis de PI3K était en fait conditionnée par le mode de recrutement de la protéine adaptatrice Gab1 et de la phosphatase SHP-2, deux acteurs indispensables à l’activation de la voie Ras/MAPK en réponse à l’EGF. Nous avons donc montré que, selon la force du signal de stimulation ou la quantité d’EGFR mobilisés, le
Introduction générale
module Gab1/SHP-2 était recruté soit par l’interaction du domaine PH (pleckstrin homology) de Gab1 avec les produits de PI3K, soit par l’intermédiaire de Grb2, rendant ainsi l’activation des MAPK dépendante ou pas de la voie PI3K. De plus, nos résultats suggèrent que cette activation des MAPK dépendante de PI3K par le mode de recrutement du module Gab1/SHP-2 pourrait être impliquée dans la résistance à un inhibiteur de l’EGFR observée dans des lignées de glioblastomes.
L’ensemble de ces résultats ont donc permis d’identifier la protéine adaptatrice Gab1 et la phosphatase SHP-2 comme deux acteurs incontournables dans l’activation de deux voies majeures impliquées notamment dans la prolifération, la croissance et la survie cellulaire : les voies Ras/MAPK et PI3K. L’implication de l’un et/ou l’autre de ces acteurs en aval de récepteurs tels que l’EGFR et le VEGFR souligne leur importance d’un point de vue physiologique lors du développement ou chez l’adulte, mais aussi lors des processus cancéreux, tant dans les cellules tumorales elles-même que dans l’angiogenèse qui leur est associée. Ainsi, Gab1 et SHP-2 pourraient être considérées comme des cibles potentielles en thérapie anti-cancéreuse.
Introduction bibliographique
PARTIE I :
Introduction bibliographique Chapitre 1 : Les RTK, exemples de l’EGFR et du VEGFR
Chapitre 1
: Les RTK, structure et mécanismes d’activation
:
Exemples de l’EGFR et du VEGFR
Chaque cellule est programmée pour répondre à divers paramètres extracellulaires lui permettant d’adapter son comportement à des modifications de son environnement. Des acteurs essentiels de ce processus sont les récepteurs membranaires car ils constituent, de par leur position à l'interface cellule/environnement, un lien indispensable entre les signaux extracellulaires et la réponse cellulaire. Dans cette famille, les récepteurs à activité tyrosine kinase intrinsèque (RTK), autrement appelés récepteurs aux facteurs de croissance, constituent un groupe majeur. En effet, ils contrôlent de nombreux processus cellulaires fondamentaux essentiels à l’homéostasie tissulaire tels que la prolifération, la différenciation, la migration, l’apoptose ou encore le métabolisme. Compte tenu de ces rôles pléiotropiques, ils sont, d’un point de vue physiopathologique, également impliqués dans diverses pathologies, notamment des maladies génétiques et de nombreux cancers où leur dérégulation est souvent associée à un mauvais pronostic.
Dans ce premier chapitre, nous ferons tout d’abord une présentation générale des RTK en résumant leurs caractéristiques communes, en matière de structure et de mécanismes d’activation. Dans un deuxième temps, nous nous intéresserons plus spécifiquement au récepteur de l’EGF (Epidermal Growth Factor) ainsi qu’au récepteur du VEGF (Vascular Endothelial Growth Factor), qui ont fait l’objet d’études au laboratoire, et représentent des acteurs majeurs dans le développement des tumeurs solides et de l’angiogenèse qui leur est associée. Nous clôturerons ensuite ce chapitre par un résumé des différents mécanismes généraux permettant la régulation des RTK.
I- Généralités sur les RTK
I-1. Introduction
Les récepteurs à activité tyrosine kinase intrinsèque (RTK) sont des glycoprotéines transmembranaires capables de lier différents agonistes par leur domaine extracellulaire et, ainsi activés, d’initier au niveau intracellulaire des cascades enzymatiques appelées voies de signalisation (Schlessinger, 2000). Ceci est rendu possible par leur activité tyrosine kinase qui leur permet de catalyser le transfert du phosphate γ d’un ATP sur des tyrosines de leur propre chaîne
Introduction bibliographique Chapitre 1 : Les RTK, exemples de l’EGFR et du VEGFR
kinase. Quelques exemples de famille de RTK sont les familles des récepteurs de l’EGF (Epidermal Growth Factor) ou EGFR/ErbB, de l’Insuline (InsR), du FGF (Fibroblast Growth Factor) ou FGFR, du VEGF (Vascular Endothelial Growth Factor) ou VEGFR, ou encore du PDGF (Platelet-Derived Growth Factor) ou PDGFR (Robinson et al., 2000). Un des RTK les plus étudiés est le récepteur de l’EGF dont nous détaillerons plus amplement la structure et les fonctions dans le paragraphe II de ce même chapitre.
I-2. Fonctions biologiques des RTK
Durant le développement, tout organisme multicellulaire doit coordonner la croissance et le maintien des différents types cellulaires et tissus qui le constituent, en intégrant des signaux environnementaux spécifiques, multiples et très complexes. De par leur capacité à réguler des processus cellulaires aussi divers que la prolifération, la migration, la différenciation et l’apoptose et compte tenu du fait que ces phénomènes sont particulièrement dynamiques au cours du développement, il semble maintenant acquis que les RTK et les voies de signalisation qu’ils induisent jouent un rôle prépondérant, lors des différentes étapes du développement. Des évidences ont tout d’abord été trouvées chez la drosophile qui présente des processus basiques de développement hautement conservés durant l’évolution et donc probablement transposables chez les mammifères. Ainsi, il a été démontré, par différentes études génétiques, que le développement de l’œil chez la drosophile, tissu modèle particulièrement utilisé pour étudier les mécanismes d’intégration du signal, se fait grâce à un réseau de connexions très finement régulées entre différents RTK, et plus particulièrement les récepteurs Sevenless et DER (l’homologue de l’EGFR chez la drosophile) (Nagaraj and Banerjee, 2004; Voas and Rebay, 2004). De même, il a été démontré que le développement des parties antérieures et postérieures des régions terminales de l’embryon dépendent de la fine régulation de Torso, RTK jouant un rôle majeur chez la drosophile (Furriols and Casanova, 2003). Enfin, il a aussi été démontré que Let-23, l’homologue de l’EGFR chez Caenorhabditis elegans, jouait aussi un rôle fondamental lors du développement, et notamment pour la formation d’une vulve normale (Aroian and Sternberg, 1991; Tan and Kim, 1999).
Mais les évidences les plus probantes de l’importance des RTK lors du développement ont été données chez les vertébrés. En effet, des souris dont on a invalidé les gènes codant pour des RTK tels que Ret ou le FGFR par exemple, montrent une létalité embryonnaire précoce associée à de profonds troubles du développement. De par leurs rôles pléiotropiques, leur activité doit être finement régulée pour donner un effet biologique adapté. De plus, la spécificité, l’intensité et la durée des différents signaux induits par les RTK sont fonction de la spécificité d’expression du récepteur lui-même et de ses cibles qui peut varier selon l’état de différenciation ou le type de cellules utilisées. Ainsi, les RTK pourront induire la même voie de signalisation dans divers types cellulaires, mais l’interprétation et l’effet biologique qui en découle pourront être différents selon les conditions considérées et le « panel » d’expression des protéines que la cellule présente (Simon, 2000).