MÉLANIE MAJOR
L’INFLUENCE DES ARBUSTES FRUITIERS SUR LA
RÉPARTITION DES OISEAUX EN SAPINIÈRE
BORÉALE
Mémoire présenté
à la Faculté des études supérieures de l’Université Laval dans le cadre du programme de maîtrise en sciences forestières
pour l’obtention du grade de maître ès sciences (M. Sc.)
DÉPARTEMENT DES SCIENCES DU BOIS ET DE LA FORÊT FACULTÉ DE FORESTERIE ET DE GÉOMATIQUE
UNIVERSITÉ LAVAL QUÉBEC
2011 © Mélanie Major, 2011
La période suivant l’envol du nid et précédant la migration est critique pour les oiseaux forestiers, particulièrement pour les juvéniles qui doivent apprendre à se nourrir efficacement avant la migration automnale. A cette période, plusieurs oiseaux forestiers deviennent principalement frugivores et se déplacent fréquemment vers les jeunes peuplements forestiers. Ce déplacement pourrait être causé notamment par (1) la recherche de petits fruits ou de couvert anti-prédateurs par les oiseaux, (2) la fréquentation des lisières de peuplements ou (3) le transit des oiseaux entre différents peuplements matures via les jeunes peuplements. J’ai testé les hypothèses de la frugivorie, des lisières et du transit à la Forêt Montmorency, Québec, durant les étés 2007 et 2008. J’ai mené une expérience de retrait de fruits testant la prédiction que le taux de capture des oiseaux dans des filets japonais serait plus faible dans les parcelles sans fruits que dans des parcelles témoins. De plus, j’ai évalué le mûrissement et la consommation des baies de sureau rouge (Sambucus
racemosa) dans les parcelles témoins suivant chaque session de capture, et modélisé les
taux de capture d'oiseaux en fonction de l’abondance des arbustes fruitiers à différentes échelles spatiales. J’ai testé les hypothèses de lisière et de transit respectivement en comparant les taux de capture à différentes distances des lisières des peuplements matures, ainsi que dans des filets placés de façon parallèle vs. perpendiculaire à ces lisières. Soixante-quatre pourcent des oiseaux, représentant 15 des 33 espèces capturées, étaient des frugivores saisonniers. Chaque année, la consommation des baies de sureau approchait 100 % en fin de saison d’échantillonnage, ce qui suggère que ces fruits pourraient être en quantité limitante. Le retrait expérimental des fruits a réduit le taux de capture des espèces frugivores de 45 %, mais n’avait aucune influence sur les autres espèces. Il n’y avait aucune relation entre l’abondance des oiseaux et des fruits au-delà de quelques mètres des filets. Les taux de capture étaient indépendants de la distance aux lisières de peuplements matures, mais ils étaient plus élevés dans les filets parallèles vs. perpendiculaires aux lisières dans le cas des espèces nichant dans les peuplements matures.
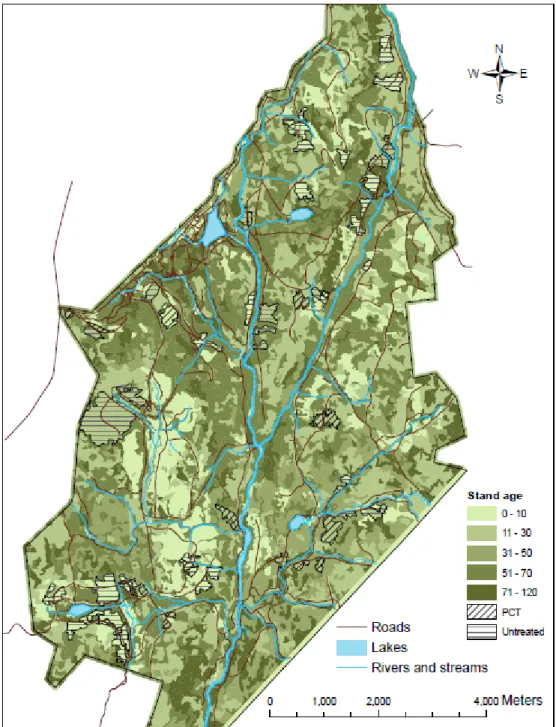
En sapinière boréale, on trouve les arbustes fruitiers surtout dans les jeunes peuplements au stade gaulis. Or, c’est à ce stade que s’applique l’éclaircie précommerciale (ÉPC), un traitement sylvicole communément utilisé pour augmenter la croissance en
Ce traitement soulève d’importantes préoccupations quant aux répercussions sur la faune associée aux milieux denses et à l’élimination possible des arbustes fruitiers dans les peuplements traités en ÉPC. Ces préoccupations ont mené à l’application expérimentale d’ÉPC à valeur faunique (ÉPCvf) à la Forêt Montmorency, où l’évaluation des tiges compétitrices est moins sévère que dans l’ÉPC conventionnelle. J'ai examiné l’impact de cette ÉPC sur l’abondance et la répartition des arbustes fruitiers en effectuant un inventaire d’arbustes fruitiers par transects dans des peuplements traités en ÉPCvf et non traités (témoins). J'ai modélisé l’abondance d’arbustes fruitiers en fonction du traitement et de variables de sites (pente, altitude et exposition). De façon générale, les résultats indiquent que le nombre d’arbustes fruitiers semble être plus élevé dans les peuplements traités en ÉPCvf par rapport aux peuplements témoins, mais la différence n’est pas significative. L’abondance d’arbustes fruitiers était très variable dans les coupes et la réponse à l’ÉPCvf variait selon les espèces. Néanmoins, l’ÉPC à valeur faunique ne semble pas avoir d’impact négatif sur l’abondance d’arbustes fruitiers. J’attribue le maintien des arbustes fruitiers au fait qu’une bonne partie ne sont pas coupés lors de l’éclaircie et, dans le cas où les tiges sont coupées, à l’augmentation de lumière et de nutriments suivant l’éclaircie qui favorise les rejets de souche. Finalement, la répartition des arbustes fruitiers était fortement agrégée (contagieuse), mais n’était pas liée à la distance aux chemins ni aux lisières de forêt mature. Cependant, l’ÉPC a significativement réduit le taux d’agrégation des arbustes fruitiers.
Il semble que l'abondance de fruits et le transit entre les peuplements matures explique en bonne partie l'abondance des oiseaux dans les jeunes peuplements en fin d'été. Ainsi, on devra s’assurer que les traitements sylvicoles en sapinière boréale maintiennent cette ressource dans une mosaïque de jeunes coupes à proximité de forêts matures. L’ÉPC pratiquée dans l’aire d’étude semble non nuisible pour les oiseaux, car l’abondance d’arbustes fruitiers y est comparable aux peuplements non-traités. Toutefois, la réduction de l’aggrégation des arbustes fruitiers pourrait avoir une influence sur l’efficacité de recherche de nourriture des oiseaux frugivores. Les résultats de cette étude sur l’abondance et la répartition des arbustes fruitiers peuvent difficilement être extrapolés à l’ÉPC conventionnelle, mais donnent un apperçu sur la dynamique des arbustes fruitiers en forêt boréale. Il est nécessaire de mieux comprendre les effets de l’ÉPC et d’autres traitements
de fruits en milieux forestier, car c’est une ressource qui est possiblement limitante pour les oisesaux en période post-reproductrice.
Abstract
The post-reproductive period is critical for many forest birds, especially for juveniles who must learn to forage on their own before the fall migration. At this period, many forest birds become mainly frugivores and songbirds of mature boreal forests often shift to early-successional stands. There are at least three explanations for this late-summer habitat shift: (1) songbirds may seek abundant fruit resources or cover against predators, (2) restrict their use of clearcuts to mature-forest edges or (3) use clearcuts mostly for transit between different patches of mature forest via early-seral stands. We tested frugivory, edge and transit hypotheses at the Forêt Montmorency, Quebec during summers 2007 and 2008. We tested the frugivory hypothesis by conducting a fruit removal experiment testing the prediction that bird capture rates in mist nets would be lower in fruit removal plots than in control plots. Additionally, we evaluated the ripening and consumption of available red elderberry (Sambucus racemosa) berries in control plots following each mist-netting session, and we modeled capture rates as a function of fruit shrub abundance at varying spatial scales. We tested edge and transit hypotheses by comparing capture rates in clearcuts near vs. away from adjoining mature forest edges, as well as in mist nets placed parallel vs. perpendicular to forest edges. Sixty-four percent of individuals, representing 15 of the 33 species captured, were seasonal frugivores. In both years of this study, fruit consumption of red elderberry approached 100 % by the end of the sampling periods, suggesting that fruit could be a limiting resource to boreal forest birds. Experimental fruit removal reduced capture rates of frugivores by 45 %, but did not affect capture rates of other species. There was no relationship between bird and fruit abundance beyond a few meters from mist nets. Capture rates were independent of distance from mature forest edges, but they were greater in nets parallel to forest edges than in those perpendicular to edges in the case of species nesting in mature forests.
In the boreal forest, fruit shrubs are mainly found in dense, early successional stands. In managed forests, it is within these stands that precommercial thinning (PCT) is applied, a treatment designed to reduce stand density so as to increase diameter growth of residual poles and orient stand species composition. The consequences of PCT on wildlife associated to dense habitat as well as the possible elimination of fruit shrubs in treated
application of wildlife-enhanced PCT at the Forêt Montmorency, where the evaluation of competing stems is less severe as in conventional PCT. I examined the impact of wildlife-enhanced PCT (wePCT) on fruit shrub abundance and distribution by conducting a fruit shrub inventory in thinned and control stands, and modeling the abundance of fruit shrubs as a function of treatment and site variables (slope, elevation and aspect). Results indicate that the abundance of fruit shrubs generally seemed higher in thinned stands, but differences were not significant. Fruit shrub abundance was highly variable in young stands and the response to thinning was species specific. Nonetheless, wePCT does not seem to have a negative impact on fruit shrub abundance. I suggest two possible explanations for the maintenance of fruit shrub following wePCT: either fruit shrubs were left uncut during thinning or, when cut, increased light and available nutrients rapidly restored stem abundance by favouring stump sprouting. Finally, fruit shrub distribution was highly aggregated, but was not related to distance to roads or to mature forest edges. However, PCT significantly reduced fruit shrub aggregation.
Given the support for frugivory and transit hypotheses, silvicultural treatments should be closely monitored to maintain fruit shrubs in small-scale patchworks of different successional stages. PCT in our study area appeared unharmful to birds, as fruit shrub abundance in PCT sites was similar to that of control stands. However, reduced fruit shrub aggregation could negatively affect foraging efficiency of frugivorous forest birds. Fruit shrub abundance and distribution results cannot be directly extrapolated to conventional PCT, but they provide insight on the fruit shrub dynamics of the boreal forest. Better understanding the effects of PCT and other sylvicultural treatments on fruits shrubs and trees is important, as fruit is possibly a limiting resource for post-breeding birds.
J’aimerais remercier André Desrochers qui, lorsque je l’ai contacté pour la première fois par rapport à une question sur le Grimpereau brun, m’a proposé de travailler pour lui comme stagiaire sans même m’avoir rencontrée. Cela m’a permis de concrétiser mon virage vers cette facette dynamique et enchanteresse de la foresterie qu’est le monde des oiseaux. Ainsi, après avoir travaillé à recueillir des données pendant 2 ans sur le Bruant à gorge blanche pour la maîtrise de Patrick Rousseau, j’ai eu la chance de m’embarquer avec André dans ce projet original sur la relation entre les oiseaux frugivores et les arbustes fruitiers à la Forêt Montmorency.
J’ai énormément appris de mon expérience à la maîtrise, particulièrement dans la planification et l’organisation de mon dispositif de recherche. J’ai rapidement découvert la nécessité de constamment s’ajuster à la variabilité de la nature, ce qui a été pour moi un défi intéressant, même si parfois frustrant, à relever! J’aimerais remercier particulièrement tous ceux qui ont participé à la récolte de données : Mathilde Jean St-Laurent, Hermann Frouin, Céline Macabiau, Ophélie Plankaert, Mélissa Chatelain et Marie-Andrée Boivert, qui ont accepté de se faire fouetter le visage à journée longue (plus souvent que jamais sous la pluie) en marchant dans du « St-Michel » pour l’inventaire d’arbustes fruitiers, qui ont accepté d’arracher du framboisier à quatre pattes et d’avoir des ampoules en faisant du débroussaillage manuel des tranchées de filet, qui ont été des compagnons « non désagréables » des matins à 3:30 et avec qui j’ai partagé l’émerveillement de tenir un oiseau dans les mains … et les accidents qui viennent avec … relâchement des muscles cloacaux en état de stress et pinçage des doigts! Merci, à toute l’équipe de la Forêt Montmorency qui a rendu nos séjours bien agréables, particulièrement à Hugues pour le support moral et à Marie et toute l’équipe de l’accueil et de la cuisine.
Pour ce qui est de la rédaction ce n’était évidemment pas ma partie préférée. Cela demande énormément de discipline et de structure, choses que j’ai du apprendre à développer! Bien que la rédaction se soit étirée plus longtemps que je ne l’aurais voulu, j’ai passé à travers lentement mais sûrement. Merci André pour les « massacres à la tronçonneuse », tes conseils ont vraiment été très appréciés et nécessaires!
des revues de calibre international.
Chapitre 1 :
Major, M. and Desrochers, A. Fruit shrub use by birds in early-successional boreal forests in the post-fledging period. À soumettre à la revue The Auk.
Chapitre 2 :
Major, M. and Desrochers, A. Precommercial thinning maintains the abundance but reduces spatial aggregation of fruit shrubs in a boreal forest. À soumettre à la Revue canadienne de recherche forestière.
Résumé ...ii
Abstract ... v
Avant-Propos ...vii
Table des matières ... x
Liste des tableaux ...xii
Liste des figures ... xiii
Introduction générale ... 14
CHAPTER 1: Fruit shrub use by birds in early-successional boreal forests in the post-fledging period ... 23 RÉSUMÉ ... 24 ABSTRACT ... 25 INTRODUCTION ... 26 METHODS ... 28 Study area ... 28
Fruit removal experiment ... 29
Fruit ripening and consumption ... 31
Fruit shrub abundance and large-scale use of clearcuts ... 31
Statistical analyses ... 35
RESULTS ... 36
Fruit removal experiment ... 37
Fruit ripening and consumption ... 39
Fruit shrub abundance and large-scale use of clearcuts ... 41
DISCUSSION ... 43
Frugivory hypothesis ... 43
Edge and transit hypotheses ... 49
Implications for forestry ... 50
ACKNOWLEDGEMENTS ... 52
LITERATURE CITED ... 52
CHAPTER 2: Precommercial thinning maintains the abundance but reduces spatial aggregation of fruit shrubs in a boreal forest ... 58
RÉSUMÉ ... 59
ABSTRACT ... 60
INTRODUCTION ... 61
METHODS ... 63
Vegetation sampling ... 65
Statistical analyses ... 67
Fruit shrub abundance among clearcuts ... 67
Fruit shrub distribution within clearcuts ... 67
RESULTS ... 68
Fruit shrub abundance among clearcuts ... 68
Fruit shrub distribution within clearcuts ... 68
DISCUSSION ... 75
Variation of fruit shrub abundance in thinned vs unthinned stands ... 75
Variation of fruit shrub abundance over time ... 77
Effects of topography on fruit shrub abundance ... 78
Fruit shrub distribution within clearcuts ... 79
Management implications ... 79
ACKNOWLEDGEMENTS ... 81
LITERATURE CITED ... 81
Conclusion générale ... 87
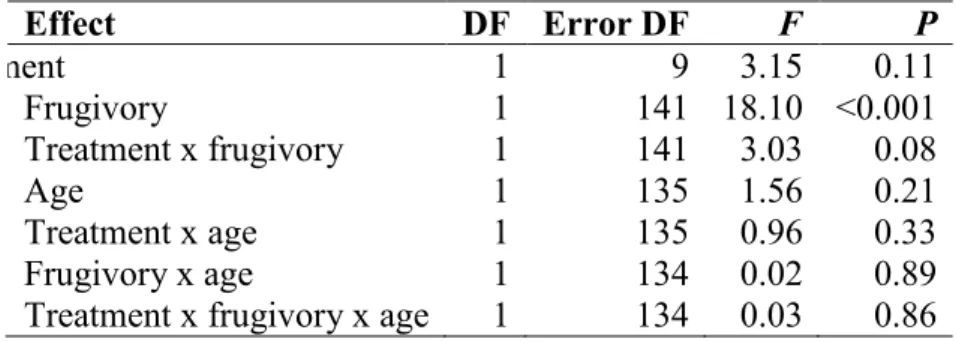
Table 1. Effects of fruit removal and bird attributes on capture rates of birds in the post-fledging period at the Forêt Montmorency. Type III tests for fixed effects are shown. ... 38 Table 2. Effect of total fruit shrub density at 4 spatial scales on capture rates of frugivores in the
post-fledging period at the Forêt Montmorency. ... 41 Table 3. Mean fruit shrub abundance, and 95 % confidence limits in untreated and
precommercial-thinned plots, as obtained from generalized models with negative binomial error distribution. Confidence limits were obtained by inversely linking the confidence bounds on the linear scale, and thus are not symmetric about the stem abundance estimates. ... 69 Table 4. Parameter estimates for fixed effects in generalized linear models of fruit shrub stem
abundance, with Negative Binomial error structure. PCT = precommercial thinning (wildlife-enhanced), with "untreated" as reference class, thus positive estimates denote greater abundance in PCT plots. ... 70 Table 5. Abundance of fruit shrubs in recent clearcuts, in relation to distance from forestry roads
and mature forest edges. Regression estimates for species abundance vs. distance were computed for each clearcut, then averaged. Regression estimates are changes in loge
(stems/0.01 ha plot) per unit increase in distance (m). ... 73 Table 6. Dispersion parameters of generalized linear models for fruit shrub distribution within
recent clearcuts, in relation to distance from forestry roads and mature forest edges. Dispersion parameters were computed for each clearcut with > 10 sampling points, then averaged. Values greater than zero represent spatially-clumped stem abundances. ... 74
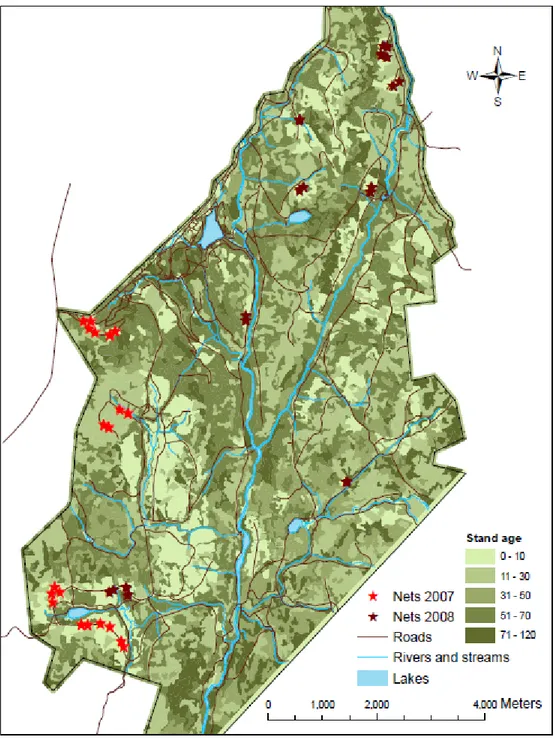
Figure 1. Mist net locations of 2007 fruit removal experimental design (in light red) and 2008 large-scale clearcut use evaluation (in dark red). ... 33 Figure 2. Example of paired plot mist net locations of 2007 fruit removal experimental
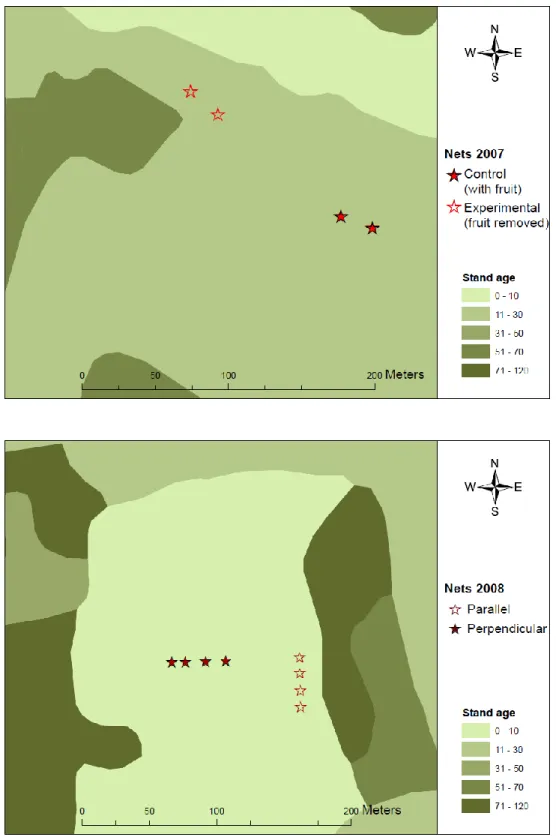
design (above). Empty stars represent mist nets where Sambucus fruits were removed within 10 m of mist nets. Example of paired plot mist net locations of 2008 large-scale clearcut use evaluation (below). Nets were places parallel (empty stars) or
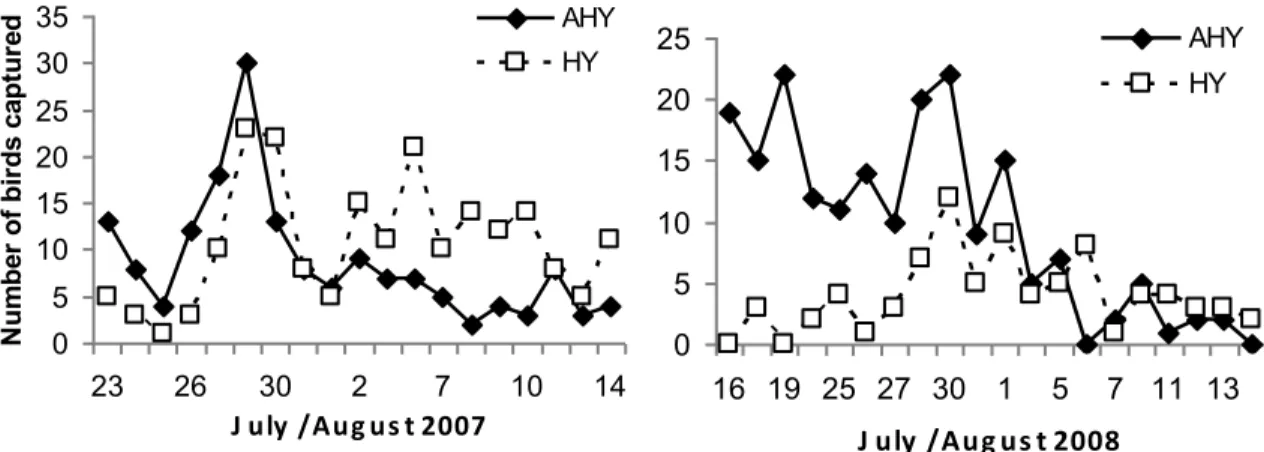
perpendicular to mature forest edge (in dark green). ... 34 Figure 3. Number of adult (AHY) and juvenile (HY) birds captured per day in 2007 / 2008.
... 37 Figure 4. Number of captured birds in treatment and control plots for frugivores and
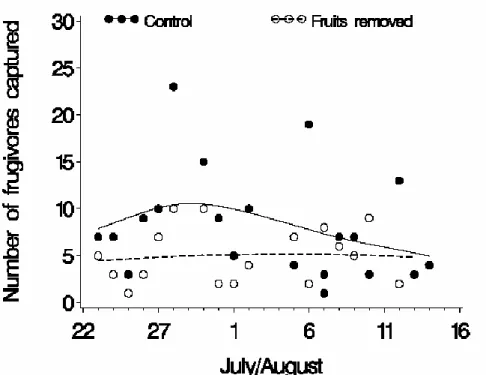
non-frugivores. ... 38 Figure 5. Number of frugivores captured in experimental (fruits removed) and control (with
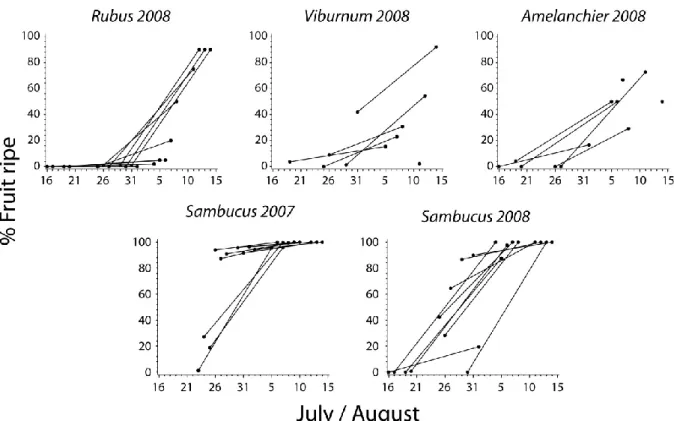
fruit) plots as a function of time. Curves obtained from a cubic spline function (SAS Institute 2008). ... 39 Figure 6. Phenology of fruit ripening at Forêt Montmorency, 2007-2008. Data from same
plots (visits 1 and 2) are joined by lines. ... 40 Figure 7. Sambucus fruit consumption at Forêt Montmorency, 2007-2008. Data points from
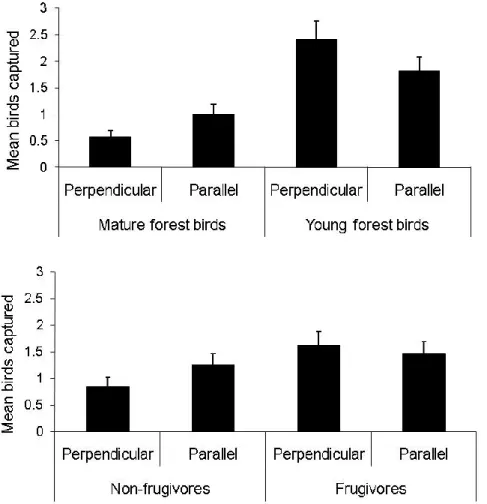
the same plots (visits 1 and 2) are joined by lines. ... 40 Figure 8. Mean number of mature and young forest birds (top) or frugivores and non-
frugivores captured (bottom) per plot, with mist nets laid out parallel or perpendicular to the nearest mature (> 40 y old) forest edge. Vertical bars are standard errors. ... 42 Figure 9. Distribution of sampled clearcuts 7 - 20 y old, with PCT (diagonal hash marks) or
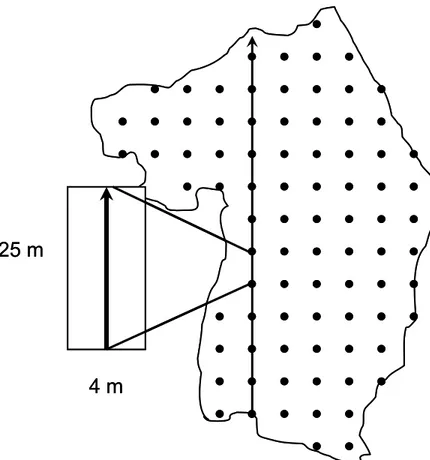
untreated (horizontal hash marks). ... 65 Figure 10. Schematic representation of the continuous vegetation sampling transects.
Transects followed a 25 m square grid and were divided into 25m segments. All fruit shrub stems were counted 2 m on either side of the central transect line and tallied per species. ... 66 Figure 11. Example of vegetation transects results for Sambucus stems. Small x marks
represent no Sambucus shrubs. Size of green circles represent number of Sambucus fruit shrubs in each 25 m transect. ... 69 Figure 12. Fruit shrub response to precommercial thinning (wePCT) following clearcut. A
spline procedure with smoothing factor of 70 was used to generate curves (SAS
Institute 2008). ... 71 Figure 13. Fruit shrub response to precommercial thinning (wePCT) in relation with slope.
A spline procedure with smoothing factor of 70 was used to generate curves (SAS Institute 2008). ... 72 Figure 14. Répartition des points d’inventaire d’arbustes fruitiers dans le peuplements
éclaircis (gris) et témoins (noir) selon le dépôt de surface, la classe de drainage et le type écologique. ... 88
Le rajeunissement des milieux forestiers par les pratiques sylvicoles a longtemps été préconisé comme pratique visant à enrichir la faune locale (Leopold 1933). Par exemple, la création de jeunes peuplements offre de nouvelles opportunités d'alimentation aux herbivores (Hughes et Fahey 1991), en rendant la végétation comestible abondante et accessible. Le maintien de peuplements matures à proximité est préconisé afin d’assurer un couvert adéquat. Ainsi, des gibiers tels que le cerf de Virginie (Odocoileus virginianus), l'orignal (Alces alces) et le lièvre (Lepus americanus) profitent d’une augmentation de la disponibilité de brout avoisinant un couvert d'abri. Comme dans les autres grands écosystèmes forestiers, à court et moyen termes, le rajeunissement des forêts profite non seulement aux herbivores, mais aussi à de nombreuses espèces d'oiseaux de milieux ouverts. En effet, suivant les pratiques forestières extensives, les communautés d’oiseaux associées aux milieux en début de succession remplacent rapidement les espèces de forêt mature en forêt boréale (Imbeau et al. 1999). Ainsi, de nombreux oiseaux migrateurs colonisent les jeunes peuplements, de sorte que la composition en espèces de ces communautés change rapidement dans les premiers 10 à 15 ans suivant la coupe (DeGraaf et Yamasaki 2003). Les jeunes peuplements sont occupés par des espèces généralistes telles que le Bruant à gorge blanche (Zonotrichia albicollis) ou spécialistes de milieux ouverts telles que la Paruline masquée (Geothlypis trichas), la Paruline à flanc marron (Dendroica
pensylvanica), le Jaseur d’Amérique (Bombycilla cedrorum) et la Paruline triste (Oporornis philadelphia).
Suivant la période de nidification, la communauté d’oiseaux dans les jeunes coupes change à nouveau, car de nombreuses espèces d’oiseaux associées aux forêts matures se déplacent vers les forêts en début de succession (Rappole et Ballard 1987, Bocetti 1993, Anders et al. 1998, Vega Rivera et al. 1998a, Pagen et al. 2000, Vitz et Rodewald 2006, 2007). Par exemple, la Grive des bois (Hylocichla mustelina) se reproduit dans les forêts matures, mais de nombreux oisillons se déplacent vers des endroits forts différents de leur site de naissance, utilisant communément des peuplements en début de succession caractérisés par une strate arbustive dense et des gaules feuillus (Anders et al. 1998, Vega Rivera et al. 1998a, Bocetti 1993). Des déplacements semblables ont aussi été observées
Pagen et al. (2000) ont aussi trouvé que certaines espèces typiques de forêts matures, comme le Viréo aux yeux rouges (Vireo olivaceus) et la Paruline à collier (Parula
americana) étaient plus abondantes dans les jeunes forêts à la période post-reproductrice
que lors de la saison de nidification. De plus, les jeunes Parulines vermivores (Helmitheros
vermivorus) étaient plus abondantes dans les jeunes forêts que dans les forêts matures à la
période post-reproductrice.
De façon générale, le déplacement des oisillons suivant la sortie du nid est influencé par les besoins immédiats de nourriture et de sécurité contre la prédation (Kershner et al. 2004). Ainsi, la fréquentation des jeunes coupes par les oiseaux de forêt mature est généralement expliquée par deux principales hypothèses. La première hypothèse est que la forte densité de ces milieux offre une importante protection contre la prédation pour les oiseaux en mue. Plusieurs études soutiennent l’hypothèse de l’évitement de la prédation et démontrent que les oiseaux sélectionnent les milieux denses pendant la saison post-reproductrice (Pagen et al. 2000, Marshall et al. 2003, Vitz and Rodewald 2006) et pendant la migration (Rodewald et Brittingham 2004). La deuxième hypothèse explique le déplacement des oiseaux vers les jeunes coupes par la présence d’arbustes fruitiers qui offrent une nourriture abondante, accessible et riche en énergie (Parrish 1997, Martin 1985, Snow 1971). En effet, plusieurs espèces d’oiseaux deviennent frugivores durant la période post-reproductrice (Suthers et al. 2000, Parrish 1997, McCarty et al. 2002). De nombreux auteurs soutiennent l’hypothèse de la frugivorie démontrant que la répartition spatiale et temporelle d’oiseaux est influencée par la disponibilité de petits fruits (White et al. 2005, Rappole et Ballard 1987, Suthers et al. 2000, Loiselle et Blake 1991). Finalement, il est possible que certaines espèces de forêt mature se trouvent dans les coupes simplement parce qu’elles les utilisent pour traverser d’un massif de forêt mature à un autre (hypothèse de transit).
Ce n’est que récemment qu’on se préoccupe des conséquences des pratiques forestières sur la faune, particulièrement de la qualité d’habitat des superficies coupées. Durant les années 70 et 80, les préceptes d'Aldo Leopold sont devenus moins populaires suite à la montée de l'écologisme. Le discours sur les mérites de l'aménagement forestier
abondance de jeunes coupes dans le paysage, garantissant la disponibilité d’habitat pour les espèces spécialistes des milieux jeunes, dépendantes de perturbations à petite ou grande échelle. Toutefois, les écologistes et gestionnaires de la faune se sont préoccupés de plus en plus des conséquences négatives de la réduction des superficies de forêt mature, de l'augmentation des lisières et de l'isolement croissant des vieilles forêts. En effet, un déséquilibre dans l’abondance des superficies des peuplements matures vs jeunes peut entraîner la disparition de certaines espèces dans le paysage (Drolet et al. 1999). La distance entre les lisières de forêt pourrait influencer l’utilisation des jeunes coupes dans le cas où les oiseaux n’y sont qu’en transit. De plus, les lisières peuvent présenter certains désavantages. Par exemple, on associe souvent un taux de prédation de nids et de parasitisme plus élevé près des lisières (Gates et Gysel 1978). Toutefois, contrairement aux forêts plus méridionales, la coupe totale ne semble pas occasionner une hausse de prédation des nids d'oiseaux dans les forêts résiduelles en régions boréales (Ibarzabal et Desrochers 2001, 2005).
Les connaissances acquises en écologie permettent de développer des méthodes de récolte et des traitements sylvicoles mieux adaptés afin de réduire l’impact sur la faune. En forêt boréale, l’évolution de la coupe à blanc vers la coupe avec protection de la régénération et des sols (CPRS) au cours des dernières années, ainsi que la limitation en superficie des aires déboisées visent à améliorer les pratiques. On assiste depuis quelques années à une renaissance des préceptes de Leopold (Leopold 1933), qui préconisait le maintien d’une hétérogénéité des milieux forestiers à l’échelle du paysage afin de répondre aux besoins d’un plus grand nombre d’espèces possible et ainsi conserver la diversité biologique (Lindenmayer et al. 2006). Par exemple, la stratégie de coupe mosaïque tente de recréer les mosaïques naturelles de composition et de structure via l’aménagement forestier en conservant des blocs de forêt résiduelle et une diversité de stades de développement (en régénération, jeunes et mûrs) dans un même paysage. On maintient des séparateurs de coupe dans l'espoir de limiter la fragmentation des habitats (Chetkiewicz et al. 2006). Les notions de filtre brut et d’espèces indicatrices sont utilisées pour assurer le maintien des éléments qui fournissent un habitat adéquat pour les espèces dans les peuplements aménagées (Noss 1990). Par exemple, le lièvre d’Amérique est une espèce clé qui a une
des prédateurs boréaux de moyenne dimension (Bujold and Bélanger 2003). Étant donnée leur sensibilité aux changements dans la structure de la végétation, les oiseaux sont aussi communément utilisés comme indicateurs biologique en vue d’évaluer l’impact de différents traitements sylvicoles sur les écosystèmes forestiers. En effet, les communautés d’oiseaux changent rapidement suite à une perturbation, témoignant de la qualité des habitats forestiers (Brawn et al. 2001).
Dans un scénario d’aménagement forestier typique en forêt boréale, la CPRS est suivie d’une éclaircie précommerciale (ÉPC) lorsque le peuplement atteint le stade gaulis. Environ une dizaine d’années suivant la coupe, ce stade de succession est caractérisé par une hauteur de 2 à 4 m ainsi qu'une forte densité de tiges en compétition. Le but de l’éclaircie est donc de réduire la densité d’un peuplement afin de concentrer la croissance sur un plus petit nombre de tiges d’essences désirées. L’effet positif de l’ÉPC sur la croissance des tiges commerciales est bien documenté (Pothier 2002, Brissette et al. 1999) et l’application de ce traitement est assez répandu en Europe et en Amérique du Nord. Dans certaines régions, particulièrement au Québec, pratiquement tous les peuplements éligibles à l’ÉPC sont traités (Canadian Council of Forest Ministers 2008). Cependant, ce traitement soulève des préoccupations par rapport à l’impact sur la biodiversité, car de nombreuses espèces affectionnent les milieux denses qui sont traités. En effet, l’ÉPC a un impact fort important sur la structure et la composition des habitats fauniques, alors que c’est la structure et la densité des tiges au stade gaulis qui assurent un milieu adéquat pour la recherche de nourriture et la nidification de nombreuses espèces d’oiseaux chanteurs (MacArthur et MacArthur 1961, Woodcock et al. 1997) et un couvert thermique et une protection contre les prédateurs pour les petits herbivores tels que les micro-mammifères (Etcheverry et al. 2005, Homyack 2003) et le lièvre d’Amérique (Sansregret 2000, Sullivan et Sullivan 1988, Ausband et Baty 2005). Les peuplements au stade gaulis offrent aussi une importante quantité de brout pour les grands herbivores tels que le cerf de Virginie (Odocoileus virginianus) et l’orignal (Doerr et Sandburg 1986, Newton et al. 1989). En appliquant l'ÉPC de manière extensive, une réduction considérable de peuplements denses dans un paysage forestier s'ensuit et réduit l’étendue de l’habitat de prédilection de nombreuses espèces telles la grive de Bicknell (Catharus bicknelli) (Connolly et al. 2002,
arbustes feuillus lors du traitement, l’ÉPC a un effet négatif immédiat sur la disponibilité de brout (Bujold et Bélanger 2003). Dans certains cas, les tiges coupées laissées au sol augmentent la disponibilité immédiate de nourriture, par exemple pour le lièvre, mais cette ressource est rapidement épuisée (Sullivan et Sullivan 1988). La diversité des communautés d’oiseaux semble être diminuée dans les années suivant une ÉPC (Sansregret 2000, Bujold et Bélanger 2003), mais la cause de ce changement demeure incertaine. On soupçonne à la fois l’effet d’une réduction de la qualité de protection contre les prédateurs ainsi qu’une diminution de la disponibilité de nourriture. Par exemple, la réduction du couvert végétal suivant l’éclaircie pourrait réduire significativement la disponibilité d'insectes et d'arbustes fruitiers, sources importantes d'alimentation pour les oiseaux chanteurs.
Afin de réduire l’impact de l’ÉPC sur la faune, des mesures de mitigation sont testées. Par exemple, des peuplements sont exclus du traitement afin de maintenir une certaine proportion de peuplements denses dans le paysage (Etcheverry et al. 2005). À l’échelle des peuplements éclaircis, des bandes ou des îlots denses sont conservés afin d’offrir un refuge pour la faune (de Bellefeuille et al. 2001, Ausband et Baty 2005, Woodcock et al. 1997). À la forêt Montmorency, Québec, une norme moins sévère est appliquée lors de l’évaluation des tiges compétitrices à dégager. Suivant cette ÉPC, dite à valeur faunique, un plus grand nombre de tiges est conservé dans les peuplements traités, entre autres des tiges feuillues qui servent de brout. Cela permet d’augmenter l’obstruction visuelle par rapport aux peuplements traités en ÉPC conventionnelle et ainsi réduire l’impact du traitement sur le lièvre et les petits mammifères (Sansregret 2000).
À ce jour, l’effet de l’ÉPC sur l’abondance des arbustes fruitiers est encore très peu documenté. Il en est de même des facteurs régissant la répartition spatiale des arbustes fruitiers en forêt aménagée. Or, l’importance des fruits pour la faune comme source de nourriture est bien connue (Martin et al. 1951). En effet, plusieurs études démontrent des corrélations importantes entre l’abondance des fruits et l'abondance locale de la faune, particulièrement pour les oiseaux frugivores en migration (White et al. 2005, Rappole et Ballard 1987, Suthers et al. 2000) et dans les milieux tropicaux (Levey et al. 1984, Levey 1988a, Levey 1988b, Martin 1985, Martin et Karr 1986, Blake et al. 1990, Loiselle et Blake
2001, Moegenburg et Levey 2003, Cohen et Lindell 2004). Des travaux récents démontrent l’importance des fruits pour les oiseaux en forêt feuillue (Vitz et Rodewald 2007), mais on connait encore très peu l’importance des fruits en forêt boréale. En sapinière boréale, les arbustes fruitiers sont principalement des essences de lumière et se retrouvent en abondance dans les jeunes milieux perturbés. Ces derniers sont donc abondants dans les jeunes coupes forestières. Dans la forêt tempérée (New Jersey), la production de fruits est maximale dans les milieux les plus ouverts, et la majorité des fruits mûrit en fin de l’été et début d’automne lorsque les arthropodes deviennent plus rares et alors que les populations d’oiseaux sont les plus élevées due au passage de nombreux migrateurs (Baird 1980). En forêt boréale, le mûrissement des fruits s’étend de la mi-juillet à la fin de l’été; ceux-ci sont donc disponibles lors de la période post-reproductrice des oiseaux forestiers. Cette période est critique, car les oiseaux doivent accumuler suffisamment de réserves de graisses pour survivre à la migration automnale, tout en subissant la mue post-reproductrice, ce qui demande un important apport d’énergie. Cette période est particulièrement critique pour les juvéniles qui sont beaucoup moins efficaces à se nourrir lorsqu’ils deviennent indépendants, ce qui influence leur taux de survie (Weathers et Sullivan 1989). Les petits fruits sont une ressource de prédilection pour accumuler des réserves énergétiques (Parrish 1997, Martin 1985, Thompson et Willson 1979, Snow 1971). De plus, ils sont facilement accessibles et abondants, ce qui minimise les dépenses énergétiques associées aux déplacements et diminue le risque de détection par les prédateurs pour les oiseaux en mue. Ainsi, la richesse en petits fruits des jeunes coupes en forêt boréale est potentiellement une ressource clé pour les oiseaux forestiers durant la période post-reproductrice.
Malgré l’importance critique de la période post-reproductrice, les connaissances sont encore limitées sur les besoins des oiseaux à cette période ainsi que sur les mécanismes qui influencent leur sélection d’habitat. Par exemple, on en sait encore très peu sur leur utilisation des jeunes coupes forestières en forêt boréale. En effet, très peu d’études ont lieu pendant la saison de la maturation des fruits, de sorte qu'il est difficile de comprendre le rôle que jouent les jeunes coupes dans le cycle annuel des oiseaux. Il se pourrait qu'en fin de saison de reproduction, les oiseaux désertent les coupes, ou au contraire y convergent soit pour s'alimenter de fruits, d'insectes ou simplement afin de
(Accipiter spp.). Il se pourrait aussi que de nombreux oiseaux se trouvent dans les coupes sans les utiliser vraiment, mais plutôt pour traverser d’un massif de forêt mature à un autre. Dans le cas où les oiseaux convergent vers les coupes pour s’alimenter de petits fruits, les activités qui ont le potentiel de réduire l’abondance d’arbustes fruitiers pourraient réduire la qualité d’habitat pour de nombreux oiseaux et donc influencer leur survie. Il est possible que l’ÉPC réduise l’abondance d’arbustes fruitiers en favorisant le dégagement des tiges résineuses. D’un autre côté, les arbustes fruitiers pourraient être favorisés par une augmentation de lumière due à une densité de tiges réduite. Afin de vérifier cela, une comparaison d’abondance d’arbustes fruitiers entre des peuplements éclaircis et témoins est nécessaire. Toutefois, cela ne permettra pas de vérifier si les fruits sont une ressource limitante pour les oiseaux en forêt boréale, c'est-à-dire si l’abondance de fruits est suffisante pour répondre à la pression de consommation. Pour évaluer cela, les méthodes expérimentales sont généralement préconisées, car même s’il est possible de vérifier s’il existe une corrélation positive entre l’abondance de fruits et l’abondance d’oiseaux, cela ne permet pas de déterminer l’impact d’une diminution de la ressource (Boutin 1990). La manipulation de la quantité de fruits permet de mesurer l’effet direct des fruits sur l’abondance relative d’oiseaux, ce qui, en lien avec le taux de consommation de ces fruits, donne un bon indice de l’importance des fruits pour les espèces frugivores. De plus, les fruits sont une ressource quantifiable de façon relativement précise et sont assez faciles à manipuler, faisant en sorte que c’est une ressource idéale pour une étude expérimentale (Blake et al. 1990a).
Objectifs
L’objectif de ce mémoire est d’évaluer l’influence des arbustes fruitiers sur la répartition des oiseaux en sapinière boréale durant la période post-reproductrice. Je répondrai à cet objectif par le biais des deux chapitres qui composent ce mémoire.
Dans le premier chapitre, l’objectif est d’évaluer expérimentalement l’importance des petits fruits pour les oiseaux forestiers et leur influence sur la sélection d’habitat des oiseaux dans les coupes récentes durant la période post-reproductrice. Je détermine aussi le taux de consommation des petits fruits. Dans un deuxième temps, j'évalue l’effet de la
mature, ainsi que le mouvement des oiseaux de forêt mature dans ces jeunes coupes. Le deuxième chapitre de ce mémoire a pour objectif d’évaluer si l’éclaircie précommerciale à valeur faunique (l’ÉPCvf) à la Forêt Montmorency à le potentiel de modifier la disponibilité de fruits (abondance) dans les coupes en sapinière pour les oiseaux en période post-reproductrice. Dans ce chapitre, j’évalue aussi l’effet de l’ÉPCvf sur la répartition spatiale des arbustes fruitiers, soit leur agrégation ainsi que leur relation avec les chemins et les lisières boisées.
Hypothèses
Si l’occurrence des oiseaux dans les jeunes coupes durant la période post-reproductrice est principalement due à la frugivorie, on devrait observer une importante diminution de cette ressource à mesure que les fruits sont consommés. Dans le même ordre d’idée, une réduction de fruits devrait avoir un impact négatif sur l’abondance relative d’oiseaux observés. Évidemment, une réduction de fruits devrait seulement affecter les oiseaux qui s’alimentent d’une telle ressource, et donc devrait avoir aucun impact sur les oiseaux non-frugivores. Dans le cas où les oiseaux utilisent activement les jeunes coupes, par exemple pour s’y nourrir ou comme endroits pour se réfugier contre les prédateurs, l’orientation des filets de capture ne devrait pas affecter le nombre d’oiseaux interceptés. D’un autre côté, si les oiseaux, particulièrement les espèces de forêt mature, se trouvent dans les coupes simplement pour effectuer un transit entre deux peuplements de forêt mature, des filets placés de façon paralèle aux lisières boisées devraient intercepter un nombre plus élevé d’oiseaux. Finalement, il est possible que les oiseaux de forêt mature frugivores utilisent plus activement les coupes pour s’y nourrir que les espèces non-frugivores. Dans un tel cas, la différence entre le taux de capture dans les filets parallèles vs. perpendiculaires aux lisières forestières devraient être réduite pour les espèces frugivores.
Puisque les arbustes fruitiers sont des tiges non commerciales qui peuvent exercer une compétition sur des tiges résineuses d’essences désirées, on s’attendrait à une réduction de l'abondance des tiges fruitières suite à l'ÉPC. Toutefois, puisque l’ÉPC à valeur faunique est moins sévère dans l’évaluation de la compétition que l’ÉPC conventionnelle, il est difficile d'anticiper son impact sur les arbustes fruitiers, qui devraient être mieux conservés
qui augmente la lumière qui atteint le sol et favorise la croissance des essences de lumière. Puisque la majorité des espèces d’arbustes fruitiers en forêt boréale sont des essences de lumière et présentent un fort potentiel de reproduction végétative, même s’ils sont coupés lors de l’ÉPC, on devrait s’attendre à ce que leur abondance soit rapidement restaurée, c'est-à-dire atteindre à nouveau les densités des sites non traités, sinon les dépasser. Dans ce contexte, on pourrait aussi s’attendre à ce que l’abondance des arbustes fruitiers soit plus importante près des chemins et des lisières, puisqu’il y a plus de lumière. Finalement, suite au traitement, on pourrait s’attendre à ce que l’aggrégation des arbustes fruitiers soit diminuée, c'est-à-dire qu’ils auront une répartition plus uniforme dans un peuplement traité, due à l’homogénéisation de la disponiblilté de ressources.
CHAPTER 1: Fruit shrub use by birds in
early-successional boreal forests in the post-fledging period
1
1 Ce chapitre sera soumis pour publication dans The Auk. Mélanie Major et André Desrochers en seront
RÉSUMÉ
La période suivant l’envol du nid et précédant la migration est critique pour les oiseaux forestiers, particulièrement pour les juvéniles qui doivent apprendre à se nourrir efficacement avant la migration automnale. A cette période, plusieurs oiseaux forestiers deviennent principalement frugivores et se déplacent fréquemment vers les jeunes peuplements forestiers. Ce déplacement pourrait être causé notamment par (1) la recherche de petits fruits ou de couvert anti-prédateurs par les oiseaux, (2) la fréquentation des lisières de peuplements ou (3) le transit des oiseaux entre différents peuplements matures via les jeunes peuplements. J’ai testé les hypothèses de la frugivorie, des lisières et du transit à la Forêt Montmorency, Québec, durant les étés 2007 et 2008. J’ai mené une expérience de retrait de fruits testant la prédiction que le taux de capture des oiseaux dans des filets japonais serait plus faible dans les parcelles sans fruits que dans des parcelles témoins. De plus, j’ai évalué le mûrissement et la consommation des baies de sureau rouge (Sambucus
racemosa) dans les parcelles témoins suivant chaque session de capture, et modélisé les
taux de capture d'oiseaux en fonction de l’abondance des arbustes fruitiers à différentes échelles spatiales. J’ai testé les hypothèses de lisière et de transit respectivement en comparant les taux de capture à différentes distances des lisières des peuplements matures, ainsi que dans des filets placés de façon parallèle vs. perpendiculaire à ces lisières. Soixante-quatre pourcent des oiseaux, représentant 15 des 33 espèces capturées, étaient des frugivores saisonniers. Chaque année, la consommation des baies de sureau approchait 100 % en fin de saison d’échantillonnage, ce qui suggère que ces fruits pourraient être en quantité limitante. Le retrait expérimental des fruits a réduit le taux de capture des espèces frugivores de 45 %, mais n’avait aucune influence sur les autres espèces. Il n’y avait aucune relation entre l’abondance des oiseaux et des fruits au-delà de quelques mètres des filets. Les taux de capture étaient indépendants de la distance aux lisières de peuplements matures, mais ils étaient plus élevés dans les filets parallèles vs. perpendiculaires aux lisières dans le cas des espèces nichant dans les peuplements matures.
ABSTRACT
The post-reproductive period is critical for many forest birds, especially for juveniles who must learn to forage on their own before the fall migration. At this period, many forest birds become mainly frugivores and songbirds of mature boreal forests often shift to early-successional stands. There are at least three explanations for this late-summer habitat shift: (1) songbirds may seek abundant fruit resources or cover against predators, (2) restrict their use of clearcuts to mature-forest edges or (3) use clearcuts mostly for transit between different patches of mature forest via early-seral stands. We tested frugivory, edge and transit hypotheses at the Forêt Montmorency, Quebec during summers 2007 and 2008. We tested the frugivory hypothesis by conducting a fruit removal experiment testing the prediction that bird capture rates in mist nets would be lower in fruit removal plots than in control plots. Additionally, we evaluated the ripening and consumption of available red elderberry (Sambucus racemosa) berries in control plots following each mist-netting session, and we modeled capture rates as a function of fruit shrub abundance at varying spatial scales. We tested edge and transit hypotheses by comparing capture rates in clearcuts near vs. away from adjoining mature forest edges, as well as in mist nets placed parallel vs. perpendicular to forest edges. Sixty-four percent of individuals, representing 15 of the 33 species captured, were seasonal frugivores. In both years of this study, fruit consumption of red elderberry approached 100 % by the end of the sampling periods, suggesting that fruit could be a limiting resource to boreal forest birds. Experimental fruit removal reduced capture rates of frugivores by 45 %, but did not affect capture rates of other species. There was no relationship between bird and fruit abundance beyond a few meters from mist nets. Capture rates were independent of distance from mature forest edges, but they were greater in nets parallel to forest edges than in those perpendicular to edges in the case of species nesting in mature forests.
INTRODUCTION
The period between fledging and migration is critical for neotropical migrant birds (Vega Rivera et al. 1998a, Pagen et al. 2000). It is characterized by high mortality rates, especially in juveniles, and thus plays a key role in the productivity of populations (Anders et al. 1997). During the post-fledging period, individuals must accumulate subcutaneous fat reserves to survive fall migration (Vega Rivera et al. 1998a, 1999, DeGraaf and Yamasaki 2003). Birds must also undergo prebasic molt which requires important energy demands to replace feathers, as well as to offset poorer insulation and flight inefficiency during this process (Jenni and Winkler 1994). During molt, birds may suffer increased predation risk because their flight ability is impaired (Pyle 1997, Vega Rivera et al. 1998b, Pagen et al. 2000). Juveniles are especially vulnerable to starvation and predation because they forage less efficiently (Heise and Moore 2003, Marchetti and Price 1989, Breitwisch et al. 1987, Weathers and Sullivan 1989) and more conspicuously (Anders et al. 1997) than adults. Additionally, juveniles must learn to interact with conspecifics and potential competitors (Weathers and Sullivan 1989).
Considering these critical needs at a period where individuals are mainly constrained by self-maintenance, we would expect birds to track resources which maximize foraging efficiency, and thus select habitats with abundant food resources (Telleria and Perez-Tris 2007). In the boreal forest, ripening of many fleshy fruits such as small berries occurs during the post-fledging period. Fruits are often abundant, which reduces searching time and thereby energy expenditure, especially for inexperienced birds (Suthers et al. 2000, Parrish 1997, Snow 1971). Foraging for fruit may be less conspicuous than foraging for insects, thereby reducing the risk of predation (Vitz and Rodewald 2007, Parrish 1997, McCarty et al. 2002). Finally, although fruits such as berries are typically low in protein (Moermond and Denslow 1985, Bairlein and Gwinner 1994), they are energy rich (Thompson and Willson 1979, Martin et al. 1951), thereby facilitating accumulation of fat reserves prior to migration (Parrish 1997, Martin 1985, Snow 1971). Thus, the increased foraging efficiency offered by feeding on fruit may increase the energy available for growth, fat deposition and molt, minimize time exposed to predation and increase time available for alternative activities (Weathers and Sullivan 1989).
Fruits in boreal and temperate forests are available on a seasonal basis. Thus, many insectivorous passerines undergo diet shifts and become seasonally frugivorous when fruits become locally available (Suthers et al. 2000, Parrish 1997, McCarty et al. 2002). However, as even the most frugivorous species (Cedar Waxwings, American Robin and Yellow-rumped Warblers, see appendix 1 for scientific names) are only seasonally so, frugivore-fruit relations have received relatively little attention in North America compared to such relations in the tropics (McCarty et al. 2002). Thus, we still have little knowledge on the importance of fruit as a food source for North American birds.
In most parts of the world, fruit shrubs are mainly found in early-successional forest stands (e.g. recent clearcuts) or in clearings (gaps) and forest edges, and become scarce in the interior of mature forests (Baird 1980, Blake and Hoppes 1986, Herrera 1995, Telleria and Perez-Tris 2007, DeGraaf and Yamasaki 2003). Mature forest birds may shift to early-successional forests following the breeding season primarily to seek fruit resources (frugivory hypothesis). Such habitat shifts during the post-fledging period have been reported by many studies. DeGraaf and Yamasaki (2003) suggest that movement of post-fledging juveniles likely reflect foraging optimization. Vega Rivera et al. (1998a) also suggested that efficient foraging most likely explained the movement of juvenile wood thrushes to early successional forests where they fed on invertebrates and fruit and later shifted back to mature forest when fruits became available in this habitat. White et al. (2005), Rappole and Ballard (1987) and Suthers et al. (2000) also found evidence supporting the frugivory hypothesis explaining habitat shifts for neotropical migrants. However, studies claiming to support the frugivory hypothesis did not rule out the possibility that birds used early-successional forests in the post fledging period mainly for reasons other than high fruit abundance. For example, Vitz and Rodewald (2007) showed that dense vegetation was responsible for most of avian use of early successional forests, although fruit was a non-negligible factor. Thus, postbreeding birds may shift habitats seeking dense cover to avoid predation as they molt (Anders et al. 1998, Vega Rivera et al. 1998b, McDermott and Wood 2010). Additionally, mature forest birds may restrict their use of recent clearcuts to mature-forest edges (Cardosa Da Silva et al. 1996) (edge hypothesis) or use such clearcuts mostly for transit between different patches of mature forest during the post-fledging period (transit hypothesis). If the occurrence of birds in
recent clearcuts during the post-fledging period is mostly due to frugivory, relative abundance of frugivores should be higher in areas with abundant fruit, and there should be a measurable decline in fruit abundance over the post-fledging period. However, to our knowledge there are no published estimates of depletion in fruit supply for the boreal forest.
There is strong evidence that individuals and populations (density) are utlimately limited by food supply (Boutin 1990). However, assessement of food limitation has been hindered by the difficulty of determining actual food availability. An alternative to measuring food availability is to experimentally manipulate food supply (Boutin 1990). Indeed, reduction in food abundance should have the most impact on birds in the case where food is in limited supply. Frugivorous birds provide a good opportunity to examine the link between resource variation and bird abundances because their primary food resource is particularly conspicuous and easily quantified and manipulated (Levey 1988a). Based on capture rates in mist nets, we conducted a fruit removal experiment to test the prediction that bird abundance will be lower in fruit removal plots than in control plots (frugivory hypothesis). Additionally, we tested edge and transit hypotheses by respectively comparing capture rates in clearcuts near vs. away from adjoining forest edges, as well as in mist nets placed parallel vs. perpendicular to mature forest edges. We predicted that capture rates would be higher near forest edges (edge hypothesis) and in mist nets parallel to forest edges (transit hypothesis), because the latter are more likely to intercept straight-line movements between mature forest patches. Finally, we provide evidence for fruit depletion based on fruit consumption rates.
METHODS
Study area
Field work was conducted at the Forêt Montmorency, Quebec (47º20’N, - 71º10’W), a 66 km2 forest managed for timber production and recreational purposes. The forest landscape is composed of regenerating forest stands of various ages, harvested primarily by the clearcut method, as well as premature and mature stands. Mature (> 40 y), premature (21 – 40 y) and regenerating (< 20 y) forest stands comprise 49 %, 23 % and 24 % of the study area, respectively (for more details see Darveau et al. 1997). Mature forest in the
study area is mostly second-growth dominated by balsam fir (Abies balsamea L.), with black spruce (Picea mariana Miller), white spruce (P. glauca Moench), and white birch (Betula papyrifera Marshall) as companion species. Regenerating forest stands are dominated by balsam fir and white birch, as well as planted white spruce and several fruit shrubs. Fruit shrub and fruit tree species at our study site are red elderberry (Sambucus
racemosa), pin cherry (Prunus pensylvanica), mountain ash (Sorbus Americana), shadbush
(Amelanchier sp.), squashberry (Viburnum edule) and raspberry (Rubus idaeus). These will subsequently be refered to as Sambucus, Prunus, Sorbus, Amelanchier, Viburnum and
Rubus respectively.
Fruit removal experiment
We selected ten clearcuts to test for frugivory as a major cause of avian use of early-successional forests in the post-fledging period. Clearcuts were randomly selected among all available clearcuts 5 - 12 y old, with vegetation 1.5 - 4 m high, slopes < 30 % and adjacent to a forestry road. In each selected clearcut, two clusters with > 6 Sambucus shrubs 100 m-2 and > 100 m apart were randomly selected and randomly assigned as control or experimental. In experimental plots, all Sambucus fruits were removed within 10 m of future mist net locations, between 16 - 18 July 2007. We focused our field experiment on
Sambucus because it is the main boreal forest fruiting shrub with available fruit in late
summer (Stutchbury et al. 2005), shrubs occurred in patches and fruit clusters were easy to remove. Also, shrubs which produce fruit in clusters, such as Sambucus may be preferred by birds as these limit traveling time while foraging (Levey et al. 1984). Species other than
Sambucus were rare (Amelanchier and Viburnum), occurred only in certain parts of the
study area (Prunus) which were excluded for the experimental design, or had ripe fruits much later than our study period (Sorbus). Rubus abundance was low and similar between paired control vs. experimental plots (Wilcoxon signed-rank test, n = 10, p = 0.94) and thus its influence on bird habitat selection was considered equal in both experimental and control plots.
Although vegetation structure of regenerating clearcuts is typically heterogeneous, structure was relatively similar among plots at a fine scale. Vegetation height ranged from 1.5 to 4 m among clearcuts, depending on clearcut age, but was similar for paired plots. No
branches or leaves were removed in experimental plots, thus no structural change occurred. We verified the absence of change in vegetation density with 4 photographs facing opposing sides of mist net locations. A 10 square grid was superimposed over each photograph and percent coverage in each was evaluated and averaged for each plot. Vegetation density was similar between paired plots (Wilcoxon signed-rank test, n = 10, p = 0.76). Removed fruits were weighed separately for each experimental plot. Samples of individual fruits and fruit cluster stems were taken and weighed separately to estimate the total number of fruits removed in each plot.
We estimated early-successional patch use by birds by using mist nets in each plot, at least one week after fruit removal (between 23 July and 15 August 2007). Mist-netting overcomes potential problems with visual or auditory detections of birds during the post-reproductive period where songbirds are often less vocal and secretive (Rappole and Ballard 1987). Dense vegetation in early-successional stands contributes to poor visual detection of birds (Pagen et al. 2000, Vitz and Rodewald 2007). Although possible biases may be associated with mist-netting (see Remson and Good 1996), it still remains the most reliable and effective method of sampling in this type of habitat (Pagen et al. 2000). Using mist nets also avoids observer biases associated with individuals’ visual and auditory detection and identification skills (Levey 1988a).
We used 2 mist nets (12 x 2.6 m, 30 mm mesh) per plot and sampled control and experimental plots from a given clearcut simultaneously. All vegetation was removed from within 1 m of mist nets. Each plot was sampled twice, with the second visit made in the same temporal sequence as the first visit. Nets were placed to maximize proximity to fruit shrubs, either perpendicular to each other or along a straight line. To avoid bias, we used the same net layout between the two plots within a given clearcut. At least 10 days separated each visit in the same plot, thus minimizing the possibility that birds learned net locations (Malizia 2001, Loiselle and Blake 1991).
Nets were opened half an hour before sunrise, closed 4 hours later and cleared of birds every 20 minutes. Banding effort for the fruit removal experiment totaled 320 mist-net-hours (4 hours x 4 nets x 20 days). All birds captured were banded with a Canadian Wildlife Service numbered leg band, and photographed, aged and sexed (when possible)
according to Pyle (1997). Age was mostly determined using wing molt patterns. Sex was determined when possible by plumage (Pyle 1997).
Fruit ripening and consumption
In summers 2007 and 2008, available Sambucus fruit in control plots was evaluated following each mist-netting session (twice per control plot overall) within 10 m either side of each mist net. Since individual fruit were impossible to count, we recorded the total number of Sambucus fruit clusters as a measure of fruit abundance (Blake et al. 1990). Clusters were classified in three groups: small, medium and large, according to a visual estimate of the number of fruit per cluster. Mean number of fruit per cluster for each cluster size was obtained from a sample of 33 clusters of each class size (117 ± 51 SD, 286 ± 63 and 439 ± 102 fruits per cluster for small, medium and large cluster sizes respectively) and was used to estimate total individual fruit abundance. Although fruit vary in size and nutrient content, Loiselle and Blake (1991) indicate that number of fruit provides a reasonable resource estimate.
Ripe and unripe fruit clusters were counted separately to determine fruiting phenology (Blake et al. 1990). Fruit clusters with fruits green to pinkish in color were identified as unripe whereas a cluster with more than half of red fruits was identified as ripe. All fruit were considered available to birds (Blake et al. 1990), since Sambucus fruit ripened rather synchronously (Figure 6) and some individuals were observed eating (likely testing) unripe fruit early in the fruiting season (M. Major, pers. obs). To estimate consumption following each mist netting session, we recorded the proportion of Sambucus fruits remaining on all clusters of shrubs within 10 m of mist nets. Mean percent of fruit remaining on Sambucus fruit clusters was then calculated for each mist netting session (Figure 7).
Fruit shrub abundance and large-scale use of clearcuts
To estimate avian use of early-successional forests and associated fruit shrubs at a larger scale, vegetation sampling was conducted throughout June to mid-July 2008 in 10 recent (5 –12 y old) clearcuts other than those used in the fruit removal experiment to evaluate species composition, abundance and distribution of fruit shrubs. Within each clearcut, we
generated a grid of points spaced 25 m apart. Fruit shrub abundance was estimated following linear transects along this grid. The total number of stems was tallied per fruit shrub species 2 m on either side of the central transect line in 25 m long segments (between two points of the grid). Thus, each point of the 25 m square grid, representing a 0.01 ha plot (4 m x 25 m), was assigned fruit shrub abundance values for each shrub species.
Two plots, each composed of three mist nets (12 x 2.6 m, 30 mm mesh) along a straight line, were placed in each of the 10 selected clearcuts (Figure 1). One plot was placed at the farthest possible point from a mature (> 40 y old) forest edge. The second plot was placed at a random location at least 18 m (half a plot's length) from the nearest mature forest edge. Distances between nets and the nearest mature forest edge ranged from 17 to 207 m. Random points with any physical constraints to mist netting (rocks, windthrow, rough topography, surface water, etc.) were eliminated. Net orientation (parallel or perpendicular) to nearest forest edge was randomly assigned between plots within each clearcut. Vegetation density was evaluated using the same method as with the fruit removal experiment and was similar between paired plots (Wilcoxon signed-rank test, n = 10, p = 0.28). With ArcGIS 9.3 (ESRI 2008), the vegetation sampling grid results were used to evaluate the number of fruit shrubs within different distances (25 m, 50 m, and 100 m) from plots to examine the extent of a possible influence of fruit abundance on bird capture rates. Species and abundance of fruit-bearing shrubs were also recorded within 2 m of net lanes on both sides and parallel to mist nets (Loiselle and Blake 1991, Suthers et al. 2000). Individual plants were tagged along mist-net transects to sample fleshy fruits and estimate fruit ripeness and consumption (McCarty et al. 2002, Poulin et al. 1992, Blake and Hoppes 1986, Thompson and Willson 1979). Following each mist net session, percent ripe fruit and fruit consumption were determined on each tagged plant.
Birds were captured between 16 July and 14 August 2008. Two paired plots from the same clearcut were sampled simultaneously each day. Each plot was sampled twice in the season, with the second visit made in the same temporal sequence as the first visit. Capture and banding procedures were similar to those for the fruit-removal experiment, with banding effort totaling 480 mist-net hours (4 hours x 6 nets x 20 days).
Figure 1. Mist net locations of 2007 fruit removal experimental design (in light red) and 2008 large-scale clearcut use evaluation (in dark red).
Figure 2. Example of paired plot mist net locations of 2007 fruit removal experimental design (above). Empty stars represent mist nets where Sambucus fruits were removed within 10 m of mist nets. Example of paired plot mist net locations of 2008 large-scale clearcut use evaluation (below). Nets were places parallel (empty stars) or perpendicular to mature forest edge (in dark green).
Statistical analyses
Capture rates in the fruit removal experiment were analyzed with a generalized mixed model (PROC GLIMMIX, SAS Institute 2008), with a Poisson distribution for the response variable, clearcut ID as a random effect, a repeated-measures design, and the Kenward-Roger method for the determination of error degrees of freedom (Schaalje et al. 2001). We examined the following fixed effects: fruit-removal treatment, frugivory, age (adult or juvenile) and interactions between the three variables. Numbers of birds captured, for different species were pooled (excluding recaptures) according to post-breeding food habits: frugivores or non-frugivores. Frugivory was a species attribute reflecting the presence of fruit in their diet (Poole 2005; see appendix 1). Species were considered seasonal frugivores if fruit-eating was mentioned in the breeding habitat sections of their Birds of North America account (Poole 2005). We expected a significant treatment x frugivory interaction, where response of frugivores to fruit removal should be greater than that of non-frugivores. We predicted that the relative abundance of non-frugivores would be independent of treatment. We also expected the effect of fruit removal would be greater on juvenile frugivores than adults, thus resulting in a significant treatment x frugivory x age interaction and a higher estimate for juvenile frugivores in plots with fruit. Finally, we tested individual species effects for most abundant species captured: White-throated Sparrow (for scientific names see appendix 1), Swainson’s Thrush and Empidonax flycatchers (least and alder pooled).
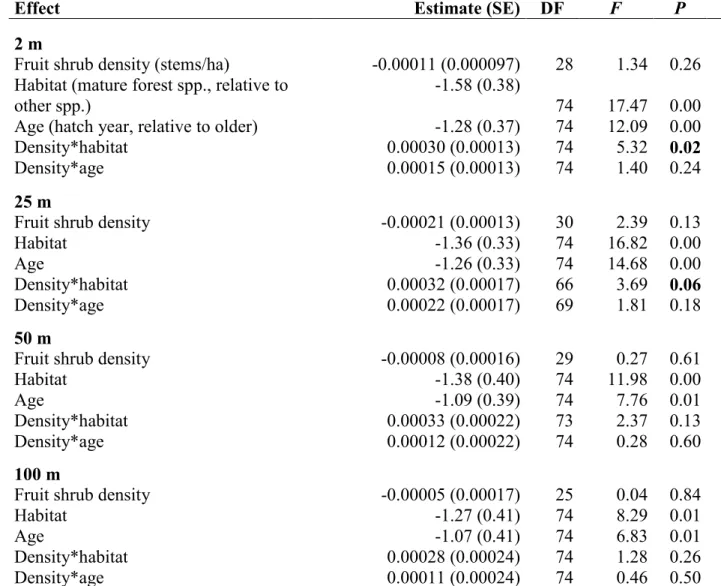
We also used Poisson mixed models to analyze capture rates as a function of large-scale fruit shrub abundance, mist net orientation and distance from mature forest edge. Because of small sample size, we limited model complexity by examining two models separately. First, we tested the relationship between the number of frugivores captured and the following fixed effects: fruit shrub stem density at different scales (2 m, 25 m, 50 m and 100 m from nets), habitat guild and bird age, as well as habitat guild x stem density and age x stem density interactions. Association with mature forest in the nesting period ('habitat guild') was obtained from Poole (2005). Species were considered as mature forest breeders if their BNA account considered mature forests as their primary or unique breeding habitat. Second, we tested the relationship between capture rates and the following fixed effects: net orientation (parallel or perpendicular to edge), distance to edge, frugivory, age, habitat
guild and interactions of last three variables with net orientation. As with the fruit removal experiment, clearcut ID was treated as a random effect, but numbers of captures from both repetitions were summed here because of lack of convergence in model estimation procedures with repeated measures.
RESULTS
In summer 2007, we captured 375 birds of 24 species. In summer 2008, we captured 275 birds of 27 species. Out of the 33 species captured, 15 were classified as seasonally frugivorous, represented by 251 and 164 individuals in 2007 and 2008 respectively (see
Appendix 1). On many occasions, we observed birds foraging in Sambucus shrubs near mist
nets, often with more than one bird foraging in the same shrub. Fruit stains on beaks of captured birds confirmed frugivory for many species. Thirteen species captured were associated to mature forest. They represented 93 (25 %) and 78 (29 %) individuals caught in 2007 and 2008 respectively.
The number of birds captured per mist-netting session was highly variable, ranging from 5 to 54 birds in 2007 and from 2 to 34 birds in 2008. White-throated Sparrow, Swainson’s Thrush, Least and Alder Flycatchers, Magnolia Warbler, and American Redstart accounted for 72 % and 65 % of total captures in 2007 and 2008 respectively. Thirty-four percent and 24 % of adult birds captured showed signs of molt in 2007 and 2008 respectively (juveniles molting: 17 % and 30 % in 2007 and 2008 respectively). More juveniles than adults were captured in 2007 (201 vs. 164), but the converse was true in 2008 (80 vs. 193). In both years, the ratio of juveniles to adults increased from mid-July to mid-August (Figure 3).