Évaluation des impacts de la fréquentation humaine sur
une colonie de mise-bas de phoques communs du
Saint-Laurent (Phoca vitulina concolor): étude de
l’échouerie de Petit-Gaspé
Mémoire
Rae Couture
Maîtrise en biologie
Maître ès sciences (M. Sc.)
Québec, Canada
© Rae Couture, 2014
Résumé
La popularité croissante de l’industrie écotouristique en milieu marin soulève des questions quant à ses effets potentiels sur les populations animales. Ce projet visait à évaluer ces effets sur la colonie de mise-bas de phoques communs du Saint-Laurent de Petit Gaspé. L’étude des réponses à court terme a révélé qu’une réduction de la distance d’approche, une augmentation du bruit ou de l’agitation de la source de dérangement et, dans une moindre mesure, de sa vitesse, augmentait l’intensité des comportements d’anti-prédation observés, mais aussi que les chiots étaient plus sensibles au dérangement. L’examen du budget d’activité de la colonie a également permis de mettre en évidence une diminution du temps dédié au repos lors des périodes de fort achalandage touristique. Par ailleurs, la fréquentation touristique n’a pas modifié l’utilisation de l’échouerie à l’échelle spatio-temporelle considérée. Le niveau de dérangement actuel pourrait avoir des répercussions néfastes sur les colonies de phoques communs ciblées et nécessiter des mesures de conservation afin d’en limiter les effets.
Abstract
The growing popularity of the ecotourism industry in marine environments raises questions about its potential effects on animal populations. This project aimed at assessing these effects over the St. Lawrence harbour seals breeding colony of Petit-Gaspé. The study of short-term responses revealed that a reduction in the approach distance, increased noise or agitation, and to a lesser extent, speed of the disturbance source, increased the intensity of anti-predatory behaviour observed, and also that the pups were more sensitive to disturbance than older seals. The study of the colony daily activity budget also revealed a decrease in the time spent resting during periods of heavy touristic traffic. Moreover, the touristic affluence did not change the use of the haul-out to the spatio-temporal scale considered. This level of disturbance could affect the targeted harbour seals colonies and requires conservation measures to limit those impacts.
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ... vii
Liste des tableaux ... ix
Liste des figures ... xi
Avant-Propos ... xiii
CHAPITRE 1 : INTRODUCTION ... 1
Risque de prédation ... 1
Impacts du dérangement aux niveaux individuel et populationnel... 3
L’industrie écotouristique ... 4
Règlementation ... 5
Biologie du phoque commun ... 6
Contexte local ... 8
Objectifs ... 9
CHAPITRE 2 SHORT AND MID-TERM IMPACTS OF HUMAN-INDUCED DISTURBANCES IN A HARBOUR SEAL (PHOCA VITULINA) COLONY ... 11
Résumé ... 12 Abstract ... 13 Introduction ... 14 Methods ... 15 Results ... 20 Discussion ... 30 Implications... 32
CHAPITRE 3 ÉVOLUTION DE LA FRÉQUENTATION DES ÉCHOUERIES DE PETIT-GASPE ET CAP-BON-AMI PAR LE PHOQUE COMMUN ... 35
Introduction ... 36
Méthode ... 37
Résultats... 39
Discussion ... 42
CHAPITRE 4 : DISCUSSION GENERALE ... 45
Fréquentation touristique ... 46
Impacts à court terme ... 47
Impacts à moyen terme ... 49
Implications... 51
Perspectives ... 52
Conclusion ... 53
Liste des tableaux
Table 1: Ethogram (modified from Boren et al. 2002) used to define harbour seals (Phoca vitulina) activity budget. ... 19 Table 2: Behavioural categories used to define anti-predatory responses displayed by harbour seals (Phoca
vitulina) during a disturbance event ... 19
Table 3 : Results from the multivariate analysis of variance with permutation (PERMANOVA) using type II SS comparing daily activity budget of harbour seals (Phoca vitulina) between different environmental conditions, tourism frequentation and life cycle stages. ... 23 Table 4 : Two-step selection of cumulative mixed logit models evaluating the influence of environmental-,
time-, seals- and disturbance-related attributes on harbour seal response intensity. Number of individual observations (n), number of parameters estimated in the model (K), model likelihood, Akaike information criterion (AIC), difference between the model with the lowest AIC and the given model (∆AIC) and AIC weight (Wt) are presented. ... 26 Table 5 : Odds ratio and inverse odds ratio† estimates‡ of disturbance attributes present in the best-supported
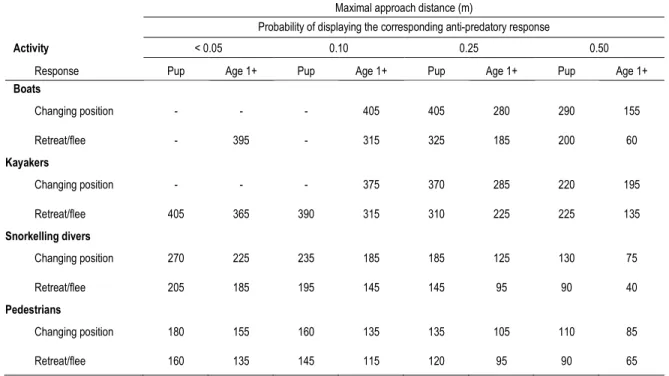
model (model E1) for each activity type... 28 Table 6: Recommended maximal approach distances from the St. Lawrence harbour seals for each activity
type considering different probabilities of occurrence of anti-predatory behaviour for pups and older seals. ... 33 Table 7 : Sélection de modèles des paramètres influençant l’évolution de la fréquentation des échoueries de
Petit-Gaspé et de Cap-Bon-Ami par le phoque commun du Saint-Laurent. ... 41 Table 8: Sélection de modèles des paramètres influençant l’évolution de la fréquentation de la portion ouest de
l’échouerie de Petit-Gaspé et de l’échouerie de Cap-Bon-Ami par le phoque commun du Saint-Laurent. ... 41
Liste des figures
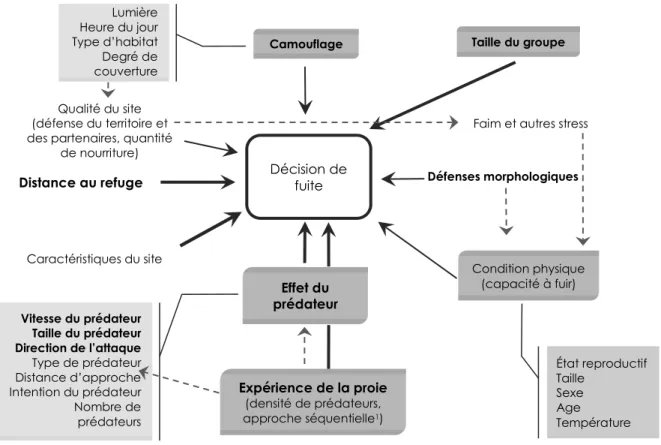
Figure 1: Résumé des facteurs potentiels pouvant influencer la décision de fuite chez les animaux. La taille et l’épaisseur des lignes pleines ainsi que des caractères indiquent les relations les plus fortes d’un point de vue statistique et théorique (i.e. les caractères gras indiquent les facteurs les plus importants). Les lignes pointillées indiquent des relations indirectes possibles entre les facteurs (adapté de Stankowich and Blumstein 2005). ... 2 Figure 2: Map of the Petit-Gaspé study site, located along the Gaspesian peninsula, Québec, Canada. The
shaded area represents the Forillon National Parc whereas the areas outside the park limits are marked with a stripped pattern. A rounded rectangle shows the location of the colony and the vantage point from where observations were completed is indicated by a star. ... 16 Figure 3 : Average number of touristic visits observed (all types pooled) at Petit-Gaspé harbour seal haul-out
site by A) week (from the first week of June) and B) time of the day during the summer seasons of 2011 and 2012... 21 Figure 4 : Average percentage of time (95% CI) dedicated by the seals to each activity for A) the low, high and
post tourism frequentation periods and B) during and outside the mating season at the Petit-Gaspé colony (Canada). Significant differences between periods are denoted by an asterisk. ... 24 Figure 5: Cumulative predicted probabilities (lines) and their 95% confidence intervals (shaded areas) for the
behavioural response of harbour seals (Phoca vitulina) to anthropogenic disturbance at Petit-Gaspé, Canada, in early June according to disturbance type (pedestrians: left columns, snorkelling divers: center-left column, kayakers: center-right columns and boats: right columns), speed level (low, moderate or high) and tourist behaviour or noise (silent, calm/low noise, moderate, agitated/ loud noise). ... 29 Figure 6 : Localisation des échoueries de mise-bas de phoques communs de Petit-Gaspé et de Cap Bon-Ami
dans le Parc national du Canada Forillon. La zone hachurée représente la portion de la péninsule exclue des limites du parc et les rectangles arrondis représentent l’étendue des échoueries de Petit-Gaspé et de Cap-Bon-Ami. ... 38 Figure 7 : Trajet et orientation des transects suivi lors de la réalisation des dénombrements sur les échoueries de Petit-Gaspé et de Cap-Bon-Ami situées dans le Parc national du Canada Forillon. Les lignes pleines représentent le trajet effectué en embarcation le long des échoueries, la ligne pointillée représente le trajet de transit entre les deux sites et la ligne discontinue représente le transect suivi lors du
dénombrement terrestre. ... 38 Figure 8 : Évolution de la fréquentation des échoueries de Gaspé (PG), Cap-Bon-Ami (CBA), et
Petit-Gaspé Ouest (PG_ouest) au cours des saisons estivales 2011 et 2012. Chaque dénombrement est présenté sous forme de pourcentage de variation par rapport à la moyenne. ... 40 Figure 9: Facteurs ayant influencé la distance de fuite chez le phoque commun du Saint-Laurent à l’échouerie de Petit-Gaspé. La taille et l’épaisseur des lignes pleines ainsi que des caractères indiquent les relations les plus fortes (i.e. les caractères gras indiquent les facteurs les plus importants). Les caractères en gris indiquent les paramètres qui n’ont pas été investigués. Les lignes pointillées indiquent des relations indirectes possibles entre les facteurs (inspiré de Stankowich and Blumstein 2005). ... 48
Avant-Propos
Ce mémoire comprend un article scientifique rédigé en anglais traitant des modifications comportementales à court et moyen termes causées par le dérangement anthropique chez le phoque commun du Saint-Laurent (Chapitre 2) et dont je suis l’auteure principale. Il sera soumis pour publication dans un journal scientifique du domaine de la conservation suivant le dépôt de ce mémoire. Le Dr. Gwénaël Beauplet a assuré la direction de ce projet et Stéphanie-Carole Pieddesaux a élaboré le protocole préliminaire et fourni son expertise technique sur le terrain. Ils figurent tous deux à titre de co-auteurs de cet article. Le troisième chapitre, plus concis, est consacré à l’analyse comparative des résultats de l’évolution de la fréquentation des échoueries de Petit-Gaspé et de Cap-Bon-Ami.
Ce projet n’aurait pu être mené à terme sans direction et je remercie Gwénaël Beauplet de m’avoir accompagnée durant ces années et de m’avoir permis d’évoluer et de grandir en tant que chercheuse et en tant que professionnelle. Il a eu confiance en mes capacités et mon autonomie et m’a laissé tenir les rênes de mon projet tout au long de sa réalisation. J’ai eu à faire des choix personnels difficiles, mais il n’a jamais douté de mon implication dans cette aventure et je l’en remercie. Ce fut un parcours ardu et exigeant, mais qui m’a permis d’exploiter mes connaissances, de développer mes capacités et d’élargir mes champs de compétences. J’en ressors plus aguerrie et prête aux défis qui m’attendent tout au long de ma vie professionnelle.
Cette étude a été réalisée avec le concours de deux précieux collaborateurs, le Parc national du Canada Forillon et le Réseau d’observation de mammifères marins (ROMM), qui m’ont apporté aide, ressources et support durant les sessions de terrain 2011 et 2012. Du Parc national du Canada Forillon, je remercie Daniel Sigouin et Mathieu Côté pour leur disponibilité et leur aide durant toute la durée du projet. Ils se sont avérés être des partenaires indispensables, toujours à l’écoute et impliqués dans le bon déroulement de cette étude. Un grand merci également à Serge Brodeur et François Tremblay qui m’ont accompagnée lors de toutes les sorties en mer. J’en ressors indemne, malgré les vagues imposantes et la brume dense qui nous ont surpris à quelques reprises. Ils étaient toujours présents, disponibles et souriants, ils ont été plus que des collègues durant ces deux étés et je me plais à croire que nous sommes devenus amis. Du ROMM, je tiens à remercier Stéphanie-Carole Pieddesaux qui a établi les fondations de ce projet et a élaboré le premier protocole, sur lequel je me suis basée pour produire celui qui m’a servi durant ces deux étés de terrain. Elle a aussi été présente pour faire le pont entre moi et le Parc au départ et m’a aidé à organiser la logistique avant mon arrivée en Gaspésie. Un grand merci également à Esther Blier, directrice du ROMM, qui a apporté son
soutien et celui de son organisme à ce projet. C’est grâce à eux que j’ai pu bénéficier de l’aide de techniciennes durant les deux saisons de terrain.
Je tiens également à témoigner toute ma gratitude à Charlène Dupasquier et Gabrielle Alix qui m’ont accompagnée durant les longues heures d’observation. Elles ont été ma 2ème paire d’yeux et mes bras de substitution, disponibles, dévouées et volontaires. Elles n’ont jamais hésité à « se mettre les mains dedans » au sens le plus littéral du terme. Avec Joannie Genesse, elles ont aussi partagé mon quotidien, supporté mes coups de barre et ont rendu ces long mois d’été loin de mes proches supportables.
J’ai vécu deux étés bien remplis durant lesquels j’ai appris à connaître la Gaspésie et les gens qui y vivent. Ils m’ont accueillie à bras ouverts et j’ai fait partie des leurs durant cette courte période. C’est en mer que j’ai vécu les plus beaux moments, entre les souffles des baleines et les oiseaux du sanctuaire. Je garde un souvenir impérissable de ces images magnifiques.
Le retour dans un bureau nécessite un certain temps d’adaptation, mais j’ai toujours été heureuse de retrouver mes collègues avec qui j’ai eu une complicité et un plaisir toujours renouvelé. Katherine Gavrilchuk, concentrée, travailleuse et discrète, mais avec qui les conversations étaient toujours productives, elle m’a aidée à avancer et j’espère ne pas trop l’avoir freinée ; Joanie Van de Walle, joviale et énergique qui m’a aidée à traverser les moments les plus pénibles de mes longs mois d’analyse de données et qui a toujours été disponible pour apporter son aide, son expérience ou 15 minutes de son temps (d’accord, soyons honnêtes plutôt 30) pour discuter ; Pierre-Étienne Lessard, au flegme proverbial qui a toujours su profiter du moment pour détendre l’atmosphère, j’espère un jour être aussi zen qu’il l’a été durant toute la durée de nos études ; et enfin, Caroline Sauvé, pétillante et décidée, la petite dernière sur laquelle on s’est un peu acharné, mais c’était fait avec toute notre affection. J’espère que « Les mots de Caroline » auront leur vie propre et continueront de faire sourire les gens qui auront la chance de la côtoyer.
CHAPITRE 1 : INTRODUCTION
L’ensemble des comportements animaux a émergé par le biais des différentes pressions sélectives auxquelles les espèces ont été soumises au cours de leur histoire évolutive (Smith et al. 2001). Parmi ces pressions figurent les interactions entre les individus et leurs conspécifiques, les autres espèces et leur environnement (Smith et al. 2001). Plusieurs processus biologiques importants comportent une composante comportementale, on peut donc en déduire qu’ils influent sur la valeur sélective des individus (Krebs and Davies 1997). Par exemple, la reproduction, l’acquisition de ressources et l’évitement de la prédation sont tous dirigés par un éventail comportemental particulier. La réalisation optimale de ces activités repose sur la valeur adaptative des comportements impliqués et influence conséquemment la survie et l’efficacité de reproduction, permettant aux gènes associés de se perpétuer dans la population. Lorsque l’environnement se modifie ou que les interactions intra et interspécifiques changent, cela entraîne une réponse évolutive réciproque des comportement impliqués qui se modulent à leur tour (Krebs and Davies 1997).
Les comportements d’anti-prédation, ont subi une pression sélective particulièrement intense, considérant qu’un échec entraine la mort de l’individu interrompant la transmission de son patrimoine génétique (Ydenberg and Dill 1986). Les stratégies permettant aux proies d’éviter la prédation sont multiples et peuvent être morphologiques ou combiner des aspects comportementaux et physiologiques (Smith et al. 2001). Par exemple, le développement d’une bonne acuité visuelle et auditive associé au comportement de vigilance permet une détection hâtive d’un prédateur potentiel et peut permettre son évitement. Les capacités de fuite sont aussi intimement liées aux aptitudes motrices de l’animal et à l’efficacité de la réponse neuroendocrine au stress (Cannon 1915). Ces comportements, déclenchés par un stimulus perçu comme menaçant (Cannon 1915), permettent à l’animal d’adapter sa réaction en fonction du risque encouru (Bouskila and Blumstein 1992).
Risque de prédation
Lors d’une rencontre avec un prédateur potentiel, l’individu doit évaluer le risque qu’il représente à partir de l’information qu’il a acquise (Stephens 1989, Giraldeau 1997, Hemmi and Pfeil 2010). Cependant, au moment de la rencontre, l’animal dispose d’un temps restreint pour réaliser cette acquisition d’information et est limité par ses propres capacités, tant cognitives que sensorielles, résultant le plus souvent en une
Figure 1: Résumé des facteurs potentiels pouvant influencer la décision de fuite chez les animaux. La taille et l’épaisseur des lignes pleines ainsi que des caractères indiquent les relations les plus fortes d’un point de vue statistique et théorique (i.e. les caractères gras indiquent les facteurs les plus importants). Les lignes pointillées indiquent des relations indirectes possibles entre les facteurs (adapté de Stankowich and Blumstein 2005).
1 Approche discontinue composée de séquences d’approches et d’arrêts.
Vitesse du prédateur Taille du prédateur Direction de l’attaque Type de prédateur Distance d’approche Intention du prédateur Nombre de prédateurs Décision de fuite Camouflage Lumière Heure du jour Type d’habitat Degré de couverture Distance au refuge Taille du groupe Effet du prédateur Expérience de la proie (densité de prédateurs, approche séquentielle1) Qualité du site (défense du territoire et des partenaires, quantité
de nourriture)
Faim et autres stress
Défenses morphologiques Condition physique (capacité à fuir) État reproductif Taille Sexe Age Température Caractéristiques du site
analyse incomplète et/ou inadéquate du risque réel de prédation (Bouskila and Blumstein 1992). L’individu extrait ces informations du prédateur et de son comportement, mais aussi de son environnement immédiat et considère sa propre condition afin d’évaluer le danger auquel il fait face (Stankowich and Blumstein 2005) et de soupeser ce risque par rapport aux bénéfices de l’activité en cours au même moment (Cooper and Frederick 2007). Stankowich et Blumstein (2005) ont résumé schématiquement les facteurs affectant directement ou indirectement la décision de prendre la fuite lors d’une rencontre avec un prédateur potentiel (Figure 1), qui appartiennent à trois catégories distinctes de facteurs, soit les caractéristiques du prédateur potentiel, de la proie et du lieu de rencontre. Le cumul des informations acquises appartenant à ces différentes sphères permet la prise d’une décision propre à chaque situation. Ces comportements ne sont par ailleurs pas associés à une image précise du prédateur (Dill 1974a, b), mais plutôt à certains signaux particuliers indicateurs du risque de prédation. Certains de ces signaux peuvent revêtir une signification particulièrement importante dans le processus d’évaluation; ils sont spécifiques à chaque espèce et sont intimement liés à son histoire évolutive (Bernays and Wcislo 1994). Un stimulus remplissant certaines conditions particulières (eg. vitesse élevée, taille importante), peut être catégorisé comme représentant une menace certaine et pousser l’animal à réagir en conséquence. Frid et Dill (2002) ont élaboré une hypothèse selon laquelle le risque de prédation est équivalent au risque de perturbation (i.e. évènement non-menaçant pouvant potentiellement dévier un individu de son comportement habituel) du point de vue de la proie et qu’elle devrait suivre les même principes économiques qui gouvernent ses réactions lors d’une rencontre avec un prédateur. Un stimulus de nature anthropogénique (i.e. présence humaine) peut donc transmettre suffisamment de signaux adéquats pour représenter un risque de prédation du point de vue de l’animal et donc entraîner un comportement d’anti-prédation.
Impacts du dérangement aux niveaux individuel et populationnel
Le dérangement des individus peut occasionner toute une gamme de répercussions, dont certaines sont immédiates telles que l’interruption du comportement de l’animal et d’autres plus diffuses découlant d’une exposition chronique au dérangement. À court terme, des activités biologiquement importantes telles que l’alimentation, les soins aux jeunes et le repos (Lima and Dill 1990, Bouskila and Blumstein 1992) peuvent être interrompues. Il peut s’écouler un temps variable avant le retour à l’état initial (Cooper 2009) et certaines opportunités (par exemple un accouplement avec un partenaire) peuvent être perdues suite à une perturbation (Cooper 1999). À plus long terme, la perception constante d’une pression de prédation, entretenue par une fréquence élevée d’événement de dérangement, peut générer un stress physiologique (Creel et al. 2002). Les impacts de ce stress physiologique chronique incluent un éventail variable de conséquences telles qu’une diminution de l’immunocompétence, de la fertilité et de la croissance ainsi qu’un vieillissement accéléré
(Johnson et al. 1992). À moyen terme, ces effets peuvent compromettre la survie et la reproduction des individus et donc diminuer leur valeur adaptative (Gill et al. 2001). Ces effets individuels cumulés peuvent se traduire à l’échelle de la population par une baisse du taux de natalité, une augmentation du taux de mortalité et donc affecter le taux de croissance de la population à plus long terme (Frid and Dill 2002). Les individus peuvent se soustraire à ces effets en désertant les zones sujettes au dérangement (Bejder et al. 2006), mais ce changement dans l’utilisation de l’habitat peut se faire au détriment de l’individu si le nouveau territoire occupé est moins profitable, par exemple si les ressources alimentaires sont moins abondante ou les opportunités de reproduction moins fréquentes.
L’industrie écotouristique
Les interactions entre les humains et les animaux sauvages sont le plus souvent involontaires et résultent d’un chevauchement dans l’utilisation de l’espace ou d’une ressource, largement dû à la croissance des populations humaines (Ramphal et al. 1996). Certaines situations font exception à cette règle comme c’est le cas de l’industrie écotouristique qui poursuit activement cet objectif, donnant aux touristes un accès privilégié permettant l’observation des animaux dans leur habitat naturel. Outre les intérêts économiques qui motivent cette industrie, elle poursuit également des objectifs d’éducation et de conservation et est perçue comme une forme durable et non-destructive d’exploitation des ressources naturelles (Powell and Ham 2008, Okech 2009, Waylen et al. 2009). Cette industrie a connu un essor important au niveau mondial durant les dernières décennies (Garrod and Wilson 2003, Samuels et al. 2003, O'Connor et al. 2009) et son caractère durable est de plus en plus remis en question (Fennell and Weaver 2005), étant donné le nombre croissant d’études démontrant les impacts à court et à long terme d’une exposition répétée au dérangement sur les individus et les populations (Kovacs and Innes 1990, Grubb and King 1991, Born et al. 1999, Suryan and Harvey 1999, Nordstrom 2002, Theuerkauf et al. 2003, Manor and Saltz 2005, Bejder et al. 2006, Lynch et al. 2010, Junge et al. 2011).
Une grande variété d’espèces, autant marines que terrestres, sont visées par l’écotourisme. Leur attrait dépend de la facilité avec laquelle il est possible de les localiser et de les observer, mais aussi de leur spécificité régionale. Les mammifères marins sont tout particulièrement prisés par l’industrie, mais les longues migrations des cétacés et leur localisation souvent difficile rend leur observation plus aléatoire. Par ailleurs, la philopatrie des pinnipèdes et leur besoin de revenir à terre périodiquement permet de prévoir leur localisation et les rend très facile à observer autant à terre qu’en mer (Cunningham et al. 2009). Les populations de pinnipèdes sont réparties un peu partout autour du globe et font l’objet d’une attention grandissante de la part de l’industrie écotouristique.
Les impacts du dérangement sur les pinnipèdes sont très variables et dépendent de l’espèce, mais aussi de l’histoire récente des populations ciblées (Constantine 1999, Cassini 2001, Kucey and Trites 2006). Kovacs et Innes (1990) ont démontré que la présence de touristes réduisait le temps de repos chez les chiots et la fréquentation maternelle du site, mais aussi le temps d’allaitement des chiots chez le phoque du Groenland (Phoca groendlandica). Chez l’éléphant de mer (Mirounga leonina), l’effet à court terme du dérangement s’est révélé modéré, soit une augmentation de la fréquence des comportements de vigilance lors d’un événement de perturbation (Engelhard et al. 2002). Les études réalisées chez les otaridés (Arctocephalus autralis, Arctocephalus forsteri) ont mis en évidence une perturbation du comportement des individus à court-terme lors de dérangements de nature anthropogéniques lorsqu’un seuil de distance était franchi (Boren et al. 2002, Cassini et al. 2004). Chez le phoque commun (Phoca vitulina), une grande variabilité a été observée dans la réponse comportementale au dérangement, rendant difficile l’élaboration de directives efficaces (Suryan and Harvey 1999, Nordstrom 2002, Andersen et al. 2012, Osinga et al. 2012).
Des études ont aussi été réalisées pour investiguer les impacts du dérangement à plus long terme sur les populations de pinnipèdes. Gerrodette et Gilmartin (1990) ont rétrospectivement constaté un abandon du site principal d’échouerie par les phoques moine d’Hawaï (Monachus schauinslandi) au profit d’un site beaucoup plus exposé aux conditions maritimes extrême et à la prédation, menant à une diminution de la survie chez les chiots. Le déclin d’une population d’otaries à fourrure australe (Arctocephalus australis) a aussi été observé au Pérou dans les années 90 et il s’est avéré que les sites exposés aux dérangements humains étaient abandonnés en plus grande proportion ou moins utilisés que les sites isolés (Stevens and Boness 2003).
Règlementation
Considérant les migrations importantes qu’effectuent annuellement les grands cétacés et la répartition étendue des différentes espèces de pinnipèdes autour du monde, plusieurs pays sont directement concernés par leur conservation et se sont dotés d’une règlementation afin de gérer les problématiques liées à l’utilisation de ces espèces ou de leur habitat. Aux Etats-Unis, le Marine Mammals Protection Act (MMPA) régit depuis 1972 les interactions entre les humains et les mammifères marins. Il interdit notamment le harcèlement des mammifères marins défini tel que :
« Tout acte de poursuite, de tourment ou d'agacement ayant le potentiel de blesser ou de déranger un mammifère marin ou une population de mammifères marins en provoquant la perturbation des comportements, y compris, mais sans s'y limiter, la
migration, la respiration, la reproduction, le soin des jeunes, l'alimentation ou l’atteinte d’un refuge. »
et interdit également les activité de plongée ou de nage avec les dauphins et les phoques (Constantine 1999, Carlson 2008). La Nouvelle-Zélande et l’Australie ont quant à eux émis une règlementation quant à la distance d’approche qui varie entre 50 et 300 mètres en fonction de l’espèce rencontrée (Carlson 2008). Au Canada, leur protection est régie par la Loi sur les pêches et la règlementation actuelle concernant les mammifères marins stipule qu’ « il est interdit d’importuner ou de déplacer un mammifère marin vivant des environs immédiats où il se trouve » (Canada 2000). Une règlementation plus précise existe dans le Sanctuaire marin du Saguenay qui limite notamment la distance d’approche entre 100 et 400m, mais cette règlementation vise exclusivement les cétacés (Carlson 2008). La popularité grandissante de l’industrie écotouristique au niveau mondial accentue le besoin de règlementations spécifiques afin de tenir compte de cette nouvelle réalité pour assurer la conservation des espèces ciblées.
Biologie du phoque commun
Le phoque commun est une espèce de phocidé largement distribuée sur les côtes de l’hémisphère Nord qui compte cinq sous-espèces se différenciant surtout par leurs aires de distribution respectives (Thompson et al. 2008). Phoca vitulina concolor est la sous-espèce répartie sur les côtes nord-ouest de l’océan Atlantique et dans les eaux arctiques (Boulva and McLaren 1979, Baird 2001).
Bien que menant une vie principalement aquatique, les pinnipèdes doivent revenir sur un substrat solide de manière périodique afin d’accomplir plusieurs activités biologiques essentielles (Riedman 1990). La mise-bas et l’allaitement des chiots ont lieu sur les échoueries ou à proximité à partir de la mi-mai dans l’estuaire et le golfe du Saint-Laurent (Dubé et al. 2003). La saison des naissances se poursuit jusqu’à la mi-juin et les chiots sont allaités durant une période moyenne de 33 jours. (Dubé et al. 2003), après quoi ils sont sevrés et doivent s’alimenter de façon autonome. Le sevrage est suivi d’une période de jeûne plus ou moins longue durant laquelle le jeune dépend essentiellement des réserves de graisses accumulées via le transfert d’énergie mère-chiot durant la lactation (Muelbert and Bowen 1993). Dès le sevrage des chiots, une ovulation est déclenchée chez les femelles ce qui débute la période d’accouplement (Boulva and McLaren 1979, Boyd 1991). Les comportements sexuels se déroulent à proximité des échoueries, où les mâles forment des leks sous-marins, permettant aux femelles de sélectionner leur partenaire (Boness et al. 2006). Après l’accouplement, l’embryon reste en diapause pour une durée moyenne de trois mois, puis s’implante dans la paroi utérine et se développe durant neuf mois de gestation active (Boulva and McLaren 1979).
La mue nécessite aussi un accès prolongé à un substrat solide à l’extérieur de l’eau, étant donné que le processus se complète plus facilement lorsque la peau et le pelage de l’animal sont complètement secs (Montagna and Harrison 1957). L’âge et le sexe des individus déterminent la phénologie de la mue : les chiots d’un an entament le processus de mue les premiers, suivi par les juvéniles, puis par les femelles adultes et enfin les mâle adultes (Daniel et al. 2003). Ce processus physiologique est influencé par le statut reproducteur de l’individu, étant donné que l’œstrogène et la progestérone sont impliqués dans la régulation de la croissance des poils chez les femelles et qu’elle est inhibée par la testostérone chez les mâles (Daniel et al. 2003) dont la concentration décline dans l’organisme après la période d’accouplement (Coltman et al. 1999). Le processus entier de perte et de régénération des poils se déroule en quatre à six semaines (Ashwell-Erickson et al. 1986) durant lequel les animaux passeront davantage de temps au sec (Daniel et al. 2003, Burns 2009). La période de mue débute au mois de juillet dans l’est du Canada (Boulva and McLaren 1979) et se poursuit jusqu’en septembre. Ce besoin accru de temps au sec durant la période estivale coïncide avec l’accroissement de la période touristique, amplifiant ainsi les impacts potentiels sur les phoques communs.
Le phoque commun occupe une position élevée dans le réseau trophique, ce qui fait de lui un prédateur important au sein des environnements marins. Il ne se situe toutefois pas au sommet du réseau et est lui aussi sujet à la prédation, faisant partie du régime alimentaire de plusieurs prédateurs supérieurs. Parmi les prédateurs importants figurent les jeunes ours polaires (Ursus maritimus; Thiemann et al. 2008), le grand requin blanc (Carcharodon carcharias; Stewart and Yochem 1985), le laimargue dormeur (Somniosus
pacificus; Sigler et al. 2006) et l’orque (Orcinus orca; Saulitis et al. 2000), soit une variété de prédateurs tant
terrestres que marins ayant chacun des stratégies de chasses particulières. Plus récemment dans l’histoire évolutive de cette espèce, les humains sont devenus un prédateur important. Le phoque commun a été chassé dans plusieurs pays du monde, presque jusqu’à l’extinction locale dans certains cas (Teilmann and Dietz 1994, Henriksen and Kondakov 1997). Au Canada, en plus d’être traditionnellement chassé par les communautés Inuit (Teilmann and Dietz 1994), une prime était offerte pour chaque phoque commun tué entre 1950 et 1976, réduisant la population des Maritimes du deux tiers (Boulva and McLaren 1979). Une si grande variété de pressions sélectives a fait en sorte que le phoque commun a aussi évolué en tant que proie et par conséquent, développé un éventail de comportement d’anti-prédation lui permettant de faire face à cette menace. Toutefois, aujourd’hui les prédateurs aquatiques et terrestres sont très peu nombreux près des côtes du Saint-Laurent et se limitent à quelques rares occurrences de requins blanc ou d’orques surtout dans le Golfe. Par contre, bien qu’aucun événement de prédation par des ours bruns et des loups n’aient été documentés, il est possible que le phoque commun fasse occasionnellement partie de leur alimentation (Thompson et al. 2008). Ces deux prédateurs potentiels sont toutefois absents de l’aire d’étude. Par ailleurs, outre la mortalité induite par la chasse traditionnelle, il arrive que des animaux soient accidentellement capturés dans des filets de pêche et s’y emmêlent conduisant le plus souvent à leur mort par suffocation
(Bjørge et al. 2002). Les phoques communs sont aussi perçus comme nuisibles par l’industrie de la pêche et sont souvent abattus étant donné qu’ils consomment certaines espèces de poissons ciblées comme le hareng, bien que leur impact sur les stocks soit considéré comme négligeable (Boulva and McLaren 1979).
Contexte local
La Gaspésie borde en partie le golfe et l’estuaire du Saint Laurent qui accueille chaque année plusieurs espèces de cétacés lors de leur migration annuelle. Il abrite également des espèces mammifères marins résidentes comme le béluga (Delphinapterus leuca) et le phoque commun, en plus de nombreuses espèces d’oiseaux marins. Cette importante biodiversité a permis l’émergence d’une industrie écotouristique florissante, soutenue en partie par la présence de plusieurs parcs nationaux qui ont pour vocation de concilier la conservation et l’utilisation des ressources naturelles à des fins récréotouristiques.
Le parc national Forillon, situé à l’extrémité est de la péninsule Gaspésienne, permet aux visiteurs d’observer une diversité faunique unique qui combine les richesses du milieu marin et du milieu terrestre. L’industrie locale s’est développée autour de ces attractions naturelles et plusieurs petits exploitants ont vu le jour, offrant majoritairement des excursions en mer et visant l’observation des mammifères marins. Trois contrats de concession sont accordés annuellement par le parc et permettent aux exploitants qui s’en portent acquéreurs d’opérer leur entreprise à l’intérieur de ses limites. La première concession est octroyée à une compagnie offrant des excursions de plongée, la seconde à l’une des deux compagnies offrant des excursions en kayaks et la troisième à une compagnie offrant des croisières d’observation de mammifères marins en bateau. Le Parc Forillon comporte trois échoueries de phoques communs situées à Petit-Gaspé, Cap-Bon-Ami et Penouille. L’échouerie de Petit-Gaspé, la plus accessible et la plus densément occupée par les animaux, est visitée quotidiennement durant la saison estivale par ces deux exploitants. La délimitation du parc fait en sorte qu’un tiers de cette échouerie est située à l’extérieur de ses limites et donc non-assujetties aux règlementations en vigueur, permettant au second exploitant offrant des excursions en kayak ainsi qu’aux randonneurs et plaisanciers qui ont été renseignés sur l’existence du site de s’y rendre pour observer les animaux. La configuration du parc fait donc en sorte que la fréquentation touristique est à la fois plus importante et moins contrôlée dans la portion située en dehors de ses limites.
Au cours des dernières années, une augmentation de la fréquentation du site a été constatée, soulevant une certaine inquiétude quant aux effets éventuels de cette hausse sur la pérennité de cette colonie. Étant donné les évidences d’effets anthropiques délétères accumulées chez plusieurs espèces de pinnipèdes tant à court qu’à long terme (Gerrodette and Gilmartin 1990, Kovacs and Innes 1990, Born et al.
1999, Suryan and Harvey 1999, Boren et al. 2002, Becker et al. 2011), une concertation des acteurs locaux a été initiée afin de répondre aux principales préoccupations concernant cette colonie. Ce projet de recherche s’inscrit dans cette initiative et vise ainsi à mieux comprendre l’impact de l’écotourisme sur le phoque commun du Saint-Laurent.
Objectifs
Cette étude a pour objectif principal d’évaluer les impacts découlant de la fréquentation humaine sur
l’échouerie de mise bas de phoques communs du Saint-Laurent de Petit-Gaspé afin d’être en mesure
d’effectuer des recommandations de directives permettant d’en atténuer les effets.
L’engouement pour l’observation des phoques communs au Parc Forillon est plutôt récent et l’achalandage touristique réel auquel les animaux sont soumis quotidiennement est inconnu. Sachant que l’affluence est variable durant la saison, il importe de déterminer la période où l’achalandage est maximal pour déterminer les classes d’âge et les processus biologiques les plus susceptibles d’être affectés par le dérangement. Afin de dresser un portrait de la situation actuelle, ce projet visait à:
1) Caractériser l’utilisation de l’échouerie de Petit-Gaspé tant par les phoques communs que par l’industrie écotouristique durant la saison estivale, soit de juin à septembre 2011 et 2012.
Considérant que les visiteurs peuvent être perçus comme des prédateurs potentiels et étant donné la variabilité des réponses comportementales observées dans les études précédentes, l’objectif spécifique suivant visait à :
2) Évaluer les impacts à court terme du dérangement sur les phoques communs de la colonie de Petit-Gaspé.
Plus particulièrement, nous cherchons à déterminer dans quelle mesure le comportement des animaux est perturbé et de définir les facteurs environnementaux, les caractéristiques individuelles et les caractéristiques de la source de dérangement influençant l’intensité des réponses d’anti-prédation afin d’en dégager des lignes directrices pour l’établissement de recommandations spécifiques.
Enfin, compte tenu des répercussions que peuvent avoir une exposition chronique aux perturbations de nature anthropique, le dernier objectif spécifique avait pour but de :
3) Évaluer les impacts à moyen terme de la fréquentation humaine sur la colonie de phoques communs de Petit-Gaspé.
L’étendue de ces impacts a été examinée sous deux angles, soit en 1) établissant le budget d’activité des animaux pour des périodes d‘achalandage touristique différentes, permettant ainsi de déterminer si un achalandage touristique important modifie l’allocation du temps aux différentes activités effectuées à terre et 2) en comparant les variations d’abondance au fil de la saison de la colonie de Petit-Gaspé (où l’achalandage touristique est important) avec celles de la colonie témoin (très peu fréquentée) de Cap-Bon-Ami. Une diminution de l’abondance relative d’individus à Petit-Gaspé durant la période où l’achalandage touristique est important pourrait indiquer un déplacement des phoques de Petit-Gaspé en réponse au dérangement. De plus, une telle diminution couplée à une augmentation de l’abondance relative d’individus à Cap-Bon-Ami pourrait indiquer un déplacement des phoques de Petit-Gaspé vers cette échouerie.
CHAPITRE 2
SHORT AND MID-TERM IMPACTS OF
HUMAN-INDUCED DISTURBANCES IN A HARBOUR SEAL
(PHOCA VITULINA) COLONY
Rae Couture1,2, Stéphanie-Carole Pieddesaux3 & Gwénaël Beauplet1,2
1. Department of Biology, Université Laval, Québec, Canada 2. Québec-Océan, Québec, Canada
Résumé
1. L'industrie écotouristique a démontré un intérêt croissant pour les pinnipèdes durant les deux dernières décennies, conduisant à une diversification des méthodes d’observation des phoques à leurs sites d’échouerie. Considérant que la présence humaine peut potentiellement provoquer des réponses d’anti-prédation et perturber les activités biologiques chez les animaux, cette étude visait à comparer les effets de différents types de perturbations humaines sur une colonie de phoques communs (Phoca vitulina). 2. L’achalandage touristique de quatre types d'activités (bateaux, kayakistes, plongeurs et piétons) ayant
cours à une colonie de reproduction de phoques communs du Saint-Laurent située dans le parc national Forillon (Québec, Canada) a été déterminée. Les effets de la présence humaine sur le comportement des individus et le budget d’activité quotidien de la colonie ont été évalués au long des étés 2011 et 2012.
3. Des échantillonnages par balayage et focaux ont été utilisés respectivement pour évaluer le budget d’activité quotidien de la colonie et pour enregistrer chaque événement de perturbation. L'intensité des réponses des phoques à la présence humaine a été évaluée en tenant compte des facteurs environnementaux et temporels ainsi que des caractéristiques des phoques et de la source de dérangement.
4. La fréquentation touristique était plus importante entre la fin-juillet et la mi-août, coïncidant avec la période post-sevrage, l'accouplement et la mue. Pendant cette période, les phoques ont passé moins de temps au repos et plus de temps dans un état actif, ce qui pourrait différer l'achèvement de fonctions vitales, telles que le maintien de l'équilibre énergétique chez les chiots.
5. De tous les types d'activités, les piétons ont provoqué le plus haut niveau de perturbation sur de courtes distances suivis par les plongeurs. Bien que les phoques aient eu tendance à présenter des réponses moins intense envers les embarcations, le niveau de perturbation causé par les bateau se déplaçant silencieusement ou les kayakistes agités se déplaçant à vitesse modérée était considérable.
6. Considérant les résultats de cette étude, nous recommandons que des distances d'approche maximales spécifiques pour chaque type d'activité touristique soient mises en place afin de minimiser le dérangement des colonies de reproduction de phoques communs.
Abstract
1. Ecotourism industry has shown a growing interest for pinnipeds, leading to a diversification of methods to observe seals in their easily accessible haul-out sites in the past two decades. As human proximity to wildlife may potentially provoke anti-predatory responses and disrupt biological activities, this study aimed to compare the impacts of different types of human disturbance on a harbour seal (Phoca vitulina) colony.
2. Touristic affluence was assessed for four types of activities (boats, kayakers, snorkelling divers and pedestrians) occurring at a St. Lawrence harbour seal breeding colony located in the Forillon National Park (Québec, Canada). The effects of human presence on the colony’s daily activity budget and seal behaviour were assessed throughout the summer of 2011 and 2012.
3. Scans and focal group sampling were used to assess the daily activity budget of the colony and to monitor each disturbance event, respectively. The intensity of seal responses to human presence was evaluated while taking into account environmental, temporal, seal-related, and disturbance type factors.
4. Tourism frequentation was higher from the end of July to mid-August, coinciding with the post-weaning, mating and moulting stages for seals. During this period, seals spent less time resting and more time in an active state, which could delay the completion of vital functions, such as maintenance of energetic balance in seal pups.
5. Of all activity types, pedestrians provoked the highest level of disturbance at close range followed by snorkelling divers. Although seals tend to exhibit milder responses toward vessels, disturbance level caused by silent boat or agitated kayakers moving at moderate speed was substantial.
6. Based on the results of this study, we recommend that specific maximal approach distances for each tourist activity type be established in order to minimize disturbance to harbour seal breeding colonies.
Introduction
During the course of evolutionary history, anti-predatory behaviour s have evolved under selective pressures to assist the survival of prey organisms (Edmunds 1974). Avoiding predation can be costly and individuals must acquire adequate information to perform a valid assessment of the potential threat (Stephens 1989, Giraldeau 1997, Hemmi and Pfeil 2010). However, animals seldom have sufficient information to evaluate the actual risk associated with every encounter (Bouskila and Blumstein 1992) and rely on simple cues like predator size, number and speed, as well as olfactory and auditory signals (Frid and Dill 2002) rather than a precise image of the predator (Dill 1974b, a). The risk assessment might also be influenced by several factors such as individual and populational traits, environmental parameters and costs related to anti-predatory behaviour (Bouskila and Blumstein 1992, Frid and Dill 2002, Stankowich and Blumstein 2005). Following the risk-disturbance hypothesis, prey encountering a disturbance stimuli should display a stronger response when the perceived risk is greater (Frid and Dill (2002). Human encounters may be interpreted as predation attempts and provoke an anti-predatory response even if not a real threat for the animal.
Recreational viewing of wildlife is usually well perceived, being a non-consumptive form of resource exploitation which promotes sustainability and pursues a conservation-oriented educational goal (Powell and Ham 2008). However, the ecotourism industry has undergone considerable growth during the last two decades (Garrod and Wilson 2003, Samuels et al. 2003, O'Connor et al. 2009) and its sustainability has been questioned (Fennell and Weaver 2005). An increasing number of studies have shown that anthropogenic disturbance cause both short-term (Kovacs and Innes 1990, Grubb and King 1991, Born et al. 1999, Suryan and Harvey 1999, Nordstrom 2002, Jansen et al. 2010, Andersen et al. 2012, La Manna et al. 2013) and long-term (Theuerkauf et al. 2003, Bejder et al. 2006, Lynch et al. 2010, Becker et al. 2011, Junge et al. 2011) impacts both at the individual- and population-level. Disturbance events may lead to acute effects like disrupting the animals’ behaviour and subsequently preventing them from carrying out essential biological activities (Lima and Dill 1990, Bouskila and Blumstein 1992). Additionally, individuals may avoid disturbed areas in favour of less profitable ones (Stevens and Boness 2003, Seuront and Prinzivalli 2005, Bejder et al. 2006). In the long term, recurrent disturbance may cause physiological stress and lead to accelerated ageing, disrupted growth and reproductive function, as well as altered immunocompetence (Johnson et al. 1992). These effects could eventually translate into higher mortality rates (Kerbiriou et al. 2009) and lower breeding frequency for the population (Giese 1996, Beale and Monaghan 2004).
The St. Lawrence Estuary and Gulf constitute rich and diversified ecosystems favoring the development of a flourishing ecotourism industry. Most activities are dedicated to whale observation, although seals have raised a growing interest in light of their philopatric use of haul-out sites that are easily accessible to tourists
from both land and sea (Stanley et al. 1996, Goodman 1998, Harkonen and Harding 2001). Among the different harbour seal colonies along the St. Lawrence coastline (Lesage et al. 1995), some are visited on a daily basis by various types of touristic activities from June to August. This peak in tourism coincides with a biologically important period for harbour seals that use haul-out sites to breed, moult and rest (Boulva and McLaren 1979). These sites also allow seals to minimize energetic expenses between foraging trips (Watts 1996). Frequent human disturbance may thus lead to the disruption of vital processes and could subsequently compromise the health of these colonies and the sustainability of ecotourism. While current regulations under the Canadian Fisheries Act (Canada 2011b) prohibit the disturbance of marine mammals, a better understanding of the impacts generated by ecotourism would enable the development of specific guidelines (e.g. maximal approach distance) to manage anthropogenic interactions with the St. Lawrence harbour seals.
This study aimed at assessing the extent of behavioural changes induced by four different types of anthropogenic disturbance in a St. Lawrence harbour seal colony at different time scales. Specific objectives were to: 1) define the current level of tourism frequentation on the colony and determine which life cycle stages could most likely be affected; 2) assess the mid-term effects of anthropogenic disturbance on the daily activity budget of harbour seals; 3) determine the characteristics of disturbance events leading to an intensification of seal anti-predatory response (short-term) and 4) compare the disturbance levels provoked by each of the four activity types (short-term). The goal of this research is to provide further insight on anthropogenic disturbance to the St. Lawrence seals, which could be used to develop specific guidelines and ensure the sustainability of the ecotourism industry.
Methods
Study area and species
The study site is located at Petit-Gaspé (PG) along the coast of the Gaspesian peninsula, southside of the Forillon National Park (48°48′ N; 64°16′ W; Figure 2). The haul-out site, separated from the road by a 75 m-wide forested strip, consists of a 3 km-long rocky shore mostly made of boulders.. This shore is mainly used as a pupping, breeding and moulting ground by the harbour seal (Phoca vitulina concolor) colony from May to September, but also as a resting site for grey seals (Halichoerus grypus) from early July. The availability of haul-out space is determined by the sea level (tidal range: 1.2-1.7 m), with almost no space available at high tide. St. Lawrence harbour seals give birth in late May and rear their young during a lactation period of 33 days after which weaning and mating occurs (Dubé et al. 2003, Van de Walle et al. in prep-b).
There are four different types of anthropogenic activities which occur regularly at the PG haul-out site from June through August : a) snorkelling tours accessing the site by boat; b) two kayak tours (only one of which has a license to operate within the park limits; Figure 2); c) summer boat traffic (marine mammal observation tours, small private vessels and a few lobster-fishing boats operating from mid-April to early July) and d) pedestrian groups, either guided or independent..
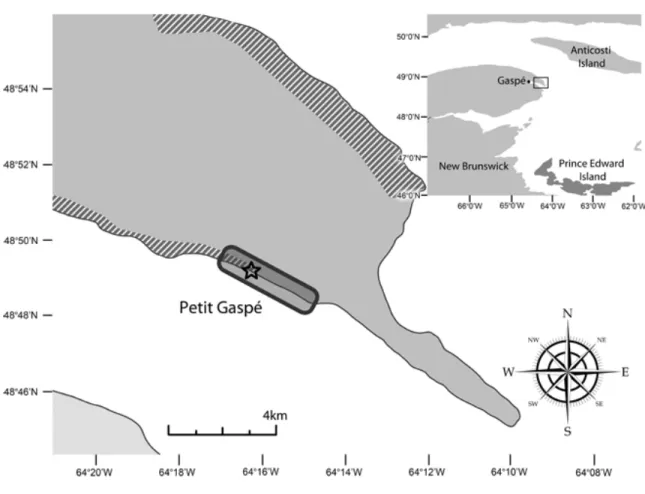
Figure 2: Map of the Petit-Gaspé study site, located along the Gaspesian peninsula, Québec, Canada. The shaded area represents the Forillon National Parc whereas the areas outside the park limits are marked with a stripped pattern. A rounded rectangle shows the location of the colony and the vantage point from where observations were completed is indicated by a star.
Observation sessions
From early June to late August 2011-2012, a four-hour observation session was undertaken every day, weather permitting (no fog, heavy rain or 2-3 m waves). Half of the observation sessions were randomly assigned to one of three 4h time windows starting at 0800, 1200 or 1600. From July 15, the last time window was gradually shifted backwards from 1600 to 1430 to ensure daylight throughout the entire observation session. As seals haul out more often during low tide (Pauli and Terhune 1987), the remaining 50% of the observation sessions were conducted during this tidal phase to optimize observation of disturbance events (i.e. any response caused by a disturbance source which disrupted seal behaviour). A first observer performed all observations in 2011 and 2012 while an additional observer was present during one third of the 2012 study period, allowing the monitoring of two simultaneous disturbance events in the study site. Observers were positioned at a fixed vantage point (Figure 2) located on an elevated natural platform at the periphery of the haul-out site with an unobstructed view. A rangefinder (Bushnell ScoutArc® 1000) allowed a 375 m-observation radius, except for small moving targets in the water (e.g. scuba divers) whose m-observation radius was 220 m.
Behavioural samplings
To assess the activity budget of hauled-out seals, instantaneous scan sampling (Altmann 1974) was performed every 30 min throughout the four-hour observation session. Each scan was completed within 1 to 3 min, during which the behaviour of each hauled-out seal was observed using binoculars (Bushnell Sportsman® 10x42) and attributed to a behavioural category as defined in Table 1. No scan sampling was undertaken during a disturbance event up until 15 min following the departure of the disturbance source (i.e. source no longer in observer’s sight). At the beginning of each observation session, environmental variables that could influence behaviour were recorded, such as tidal stage (converted to sea level with tide tables (Fisheries and Oceans Canada 2012)), wave height (1: little wave, no out space restriction, 2: > 25% out space restriction, 3: 25-50% haul-out space restriction, 4: 50-100% haul-haul-out space restriction) and weather (1: less than 50% of cloud cover, 2: more than 50% of cloud cover, 3: rainy or foggy).
Each disturbance event was recorded using focal sampling (Altmann 1974), with the group of seals disturbed considered as the focal unit. Here, a group was defined as hauled-out seals where the two most distant individuals were not separated by more than 20 m. A disturbance event started when the first seal responded and ended when all seals resumed their original behaviour, returned to a resting state, or fled into water. Both seals and disturbance sources were alternately observed using binoculars throughout the entire disturbance duration. The most intense behaviour displayed by each seal within the focal group was recorded according to the behavioural
categories defined in Table 2.
Characteristics of the disturbance source (human group size, activity, speed and behaviour) as well as seal characteristics (seal group size, age (pup or age 1+)) were also recorded. Human group size was defined as the number of people and/or boats present; speed was ranked as slow, medium or fast within each activity while behaviour was ranked from 1 to 4 (1: silent, 2: calm, low noise level, 3: moderate agitation or noise level, 4: agitated, loud or shouting). When the disturbance source was a boat, only the noise emission was ranked. The minimal distance between the seal group and the disturbance source (𝑎) was calculated using the following formula:
𝑎 = �𝑏2+ 𝑐2− 2𝑏 ∙ 𝑐 ∙ cos 𝜃
where b is the distance between the observer and the seal group, c is the distance between the disturbance source and the observer and θ represents the angle between b and c, measured using a laser rangefinder and a compass
when the space separating the seals and the disturbance source was deemed minimal. The duration of the disturbance event was defined as the time elapsed between the first seal’s response and the time when all disturbed seals resumed their original behaviour, returned to a resting state, or fled into the water. All data were recorded directly on a spreadsheet using a mobile digital tablet (iPad®, Apple 2010).
Data analysis
All analyses were performed within the R environment (R Core Team 2012). Generalized linear models (GLM) with a quasi-Poisson distribution where used to examine the temporal trend of touristic frequentation daily (AM, PM and evening) and over the season (number of weeks since June 1). A multiple comparison test (Tukey HSD) was applied when significant differences were detected.
Consistency between the two observers in 2012 was tested using a AC1 statistic (Gwet 2002). From this test, observer bias was deemed minimal for both types of sampling (scan and focal sampling = 0.794 ± 0.049 and 0.958 ± 0.005, respectively), thus data from both observers were pooled. The relationship between seal activity budget, environmental conditions (weather, wave height and sea level), life cycle stage (nursing, mating and moulting), and the tourist affluence was investigated using PERMANOVA (package vegan; Oksanen et al. 2013) instead of a regular MANOVA because the assumption for normality was not met upon arcsin transformation. Significant relationships were further analyzed with ANOVA using permutation tests (package lmPerm; Wheeler 2010). A significance level of 10% was considered to protect against type II error and results are presented as means ± SE, unless stated otherwise.
Table 1: Ethogram (modified from Boren et al. 2002) used to define harbour seals (Phoca vitulina) activity budget.
Behavioural category Description
Resting Lying down with eyes closed or open Grooming Scratching, comfort movements
Care Cow lying on the side, pup in physical contact with his mother (including suckling events) Changing position Shifting position without moving from one point to another
Moving Active displacement, on land or towards water
Agonistic behaviour Growl, foreflipper scratch or wave toward another individual
Table 2: Behavioural categories used to define anti-predatory responses displayed by harbour seals (Phoca vitulina) during a disturbance event
Behavioural category Description
Neutral No changes in behaviour
Alertness Head up towards disturbance source, eyes open, aware with or without continuous attention Changing position Waddle or align themselves in the direction of the disturbance source
Retreat Move towards the sea but stop before reaching it Flee Move into the water
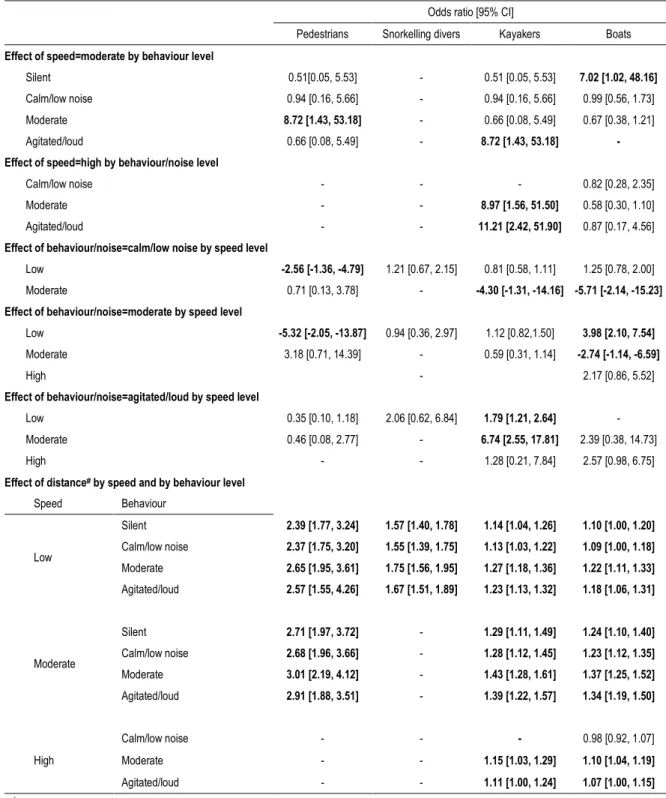
To assess which components of the interaction between human and seals were more contributory in initiating anti-predatory behaviour, the intensity of the response was modelled using cumulative logit mixed models (Agresti 2003) with the ordinal package (Christensen 2012). Such analysis allows for assessing the effect of the predictor variables over an ordinal response variable, such as behaviours of variable intensity. The response variable here was the anti-predatory response and ranged from “neutral” to “flee”. Because of the several different categories of predictor variables, candidate models were developed in two separate steps, following St-Louis et al. (2013). During the first step, different sets of candidate models were built for each category of predictor: 1) environmental attributes (waves height, weather and sea level); 2) time attributes (number of hours since dawn, number of days since June 1 and year); 3) seal attributes (age and group size); and 4) disturbance attributes (activity type, approach distance, speed, behaviour and humans or boat group size). Relevant interactions were tested when appropriate. To control for the effect of the neighbours response and for multiple sampling of the same individual, the group to which the individual belonged was set as a random effect. The threshold estimates for the “retreat” and “flee” response categories were very similar and were therefore collapsed. The resulting models were sorted according to their Akaike’s Information Criterion score (AIC; Akaike 1973), with all the models having a ∆AIC value of <4 considered to have a relatively strong support given the data, and passed on to the final step (Burnham and Anderson 2002, Burnham et al. 2011). In the final step, these candidate models were combined to create a set of global models that were subsequently ranked according to their AIC. The contribution of each variable included in the final step was assessed with their respective odds ratio (OR) and associated 95% confidence intervals. When a variable had a negative effect on the intensity of the response, the inverse odds ratio (IOR; 1/OR) was used instead to present the effect on the same metric as the ones with positive coefficients (DesJardins 2001). Estimates for variables present in an interaction term with the approach distance were calculated based on a disturbance event occurring at 100 m from the colony, which is in line with the current voluntary guidelines recommended by the Department of Fisheries and Oceans Canada.
Results
Tourism frequentation
In 2011, a total of 244 hours of observation were made during which 239 touristic visits (68 boats, 87 kayaks groups, 51 pedestrian groups and 33 snorkelling groups) occurred while 228 touristic visits (61 boats, 107 kayak groups, 23 pedestrian groups and 37 snorkelling groups) were observed during 198 hours of observation conducted in 2012. The daily tourism frequentation was similar between the two study years (2011: 3.8 ± 3.1 groups/day; 2012: 4.5 ± 3.1 groups/day; GLM, t = 0.503, p = 0.62). However, the effect of the
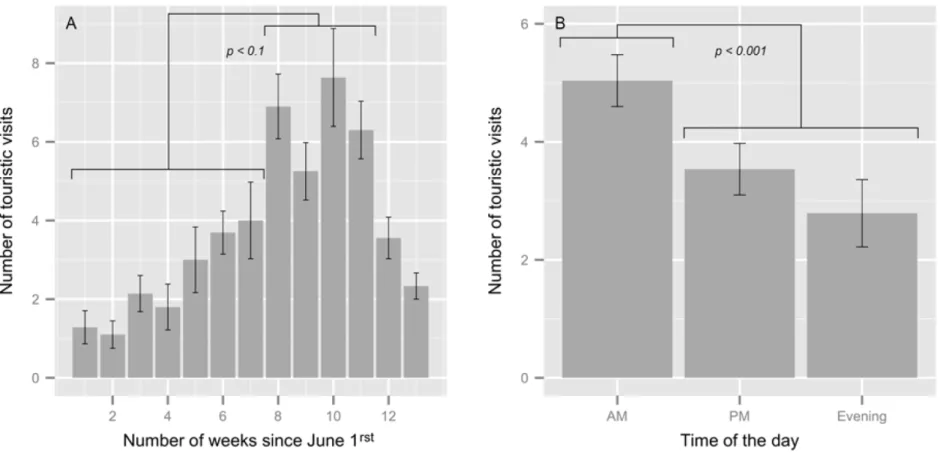
Figure 3 : Average number of touristic visits observed (all types pooled) at Petit-Gaspé harbour seal haul-out site by A) week (from the first week of June) and B) time of the day during the summer seasons of 2011 and 2012.
date was equivocal and post-hoc Tukey’s HSD tests were subsequently performed, resulting in the identification of two distinct touristic periods (Figure 3(A)): one of low frequentation from the first week of June to the third week of July (1.5 ± 1.2 groups/day) and another period of high frequentation extending from the last week of July to the third week of August (6.1 ± 3.3 groups/day). By the end of August, the tourism frequentation was not different from either of these two periods (p > 0.15 in all cases; Figure 3(A)) and was thereafter regarded as the onset of the post-tourism period (3.6 ± 1.6 groups/day). Time of the day (AM, PM or evening) had a significant effect on visit occurrence. Tourism frequentation was higher during the morning and decreased throughout the afternoon and evening (Tukey HSD, AM-PM, z = -3.796, p < 0.001; AM-evening, z = -4.49, p < 0.001, PM-evening, z = 1.747, p = 0.184; Figure 2 (B)). The duration of a touristic visit at the haul-out site ranged between 1 and 141 min (overall mean 44 ± 1 min), and averaged 46 ± 2 min for kayakers, 59 ± 2 min for divers, 28 ± 3 min for pedestrians and 44 ± 3 min for boats.
Activity budget
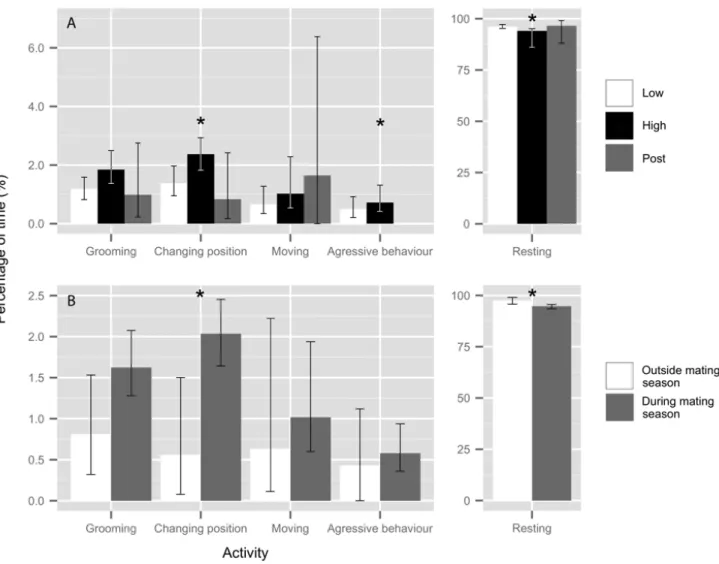
Instantaneous scan sampling of the colony yielded a total of 10,141 individual behavioural records from 281 scan samples collected over 106 days. Throughout the study period, the vast majority of the time seals spent hauled out was dedicated to rest (95.0 ± 0.1%) while grooming, changing position, moving and aggressive behaviour accounted for a smaller portion of the daily activity budget (Figure 4). The activity budget was altered by touristic affluence and during the mating season (Table 3). During the period of high tourism frequentation, seals spent less time resting and tended to spend more time changing position compared to the two other periods (ANOVA, resting: F2,95= 3.66, p = 0.03; changing position: F2,95= 3.00, p = 0.06; Figure
4(A)). Interestingly, aggressive behaviour was never observed during the post tourism period (Figure 4(A)), while the time dedicated to other activities did not differ between tourism frequentation periods (ANOVA, grooming: F2,95 = 1.1, p = 0.34; moving: F2,95= 0.14, p = 0.87; Figure 4(A)). Similarly, during the mating
season, resting time was significantly lower; whereas the time spent changing position was higher than the rest of the study period (ANOVA, resting: F1,96 = 4.48, p = 0.04; grooming: F1,96= 1.55, p = 0.22; changing
position: F1,96= 5.56, p = 0.02; moving: F1,96 = 0.05, p = 0.82; aggressive behaviour: F1,96= 2.05, p = 0.16;
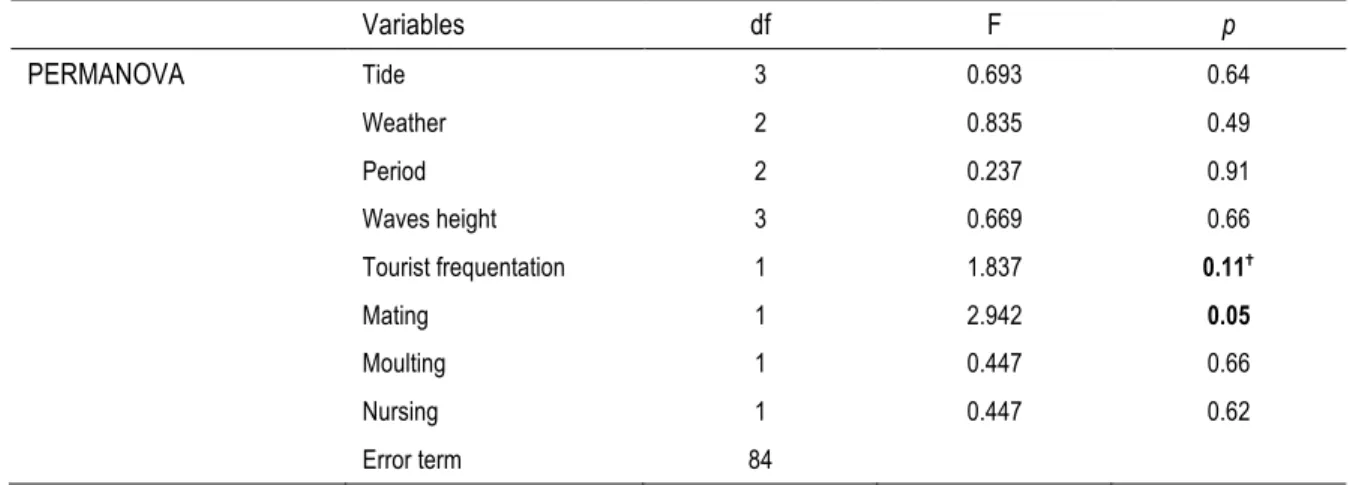
Table 3 : Results from the multivariate analysis of variance with permutation (PERMANOVA) using type II SS comparing daily activity budget of harbour seals (Phoca vitulina) between different environmental conditions, tourism frequentation and life cycle stages.
Variables df F p PERMANOVA Tide 3 0.693 0.64 Weather 2 0.835 0.49 Period 2 0.237 0.91 Waves height 3 0.669 0.66 Tourist frequentation 1 1.837 0.11✝ Mating 1 2.942 0.05 Moulting 1 0.447 0.66 Nursing 1 0.447 0.62 Error term 84
Figure 4 : Average percentage of time (95% CI) dedicated by the seals to each activity for A) the low, high and post tourism frequentation periods and B) during and outside the mating season at the Petit-Gaspé colony (Canada). Significant differences between periods are denoted by an asterisk.
Short-term response
A total of 1471 interactions (573 in 2011 and 898 in 2012) following human approaches were documented. Overall, 28.4% of these interactions resulted in the retreat or the flee of at least one seal, which represented 45.2% for pedestrians, 56.0% for snorkelling divers, 21.0% for kayakers and 22.5% for boats.
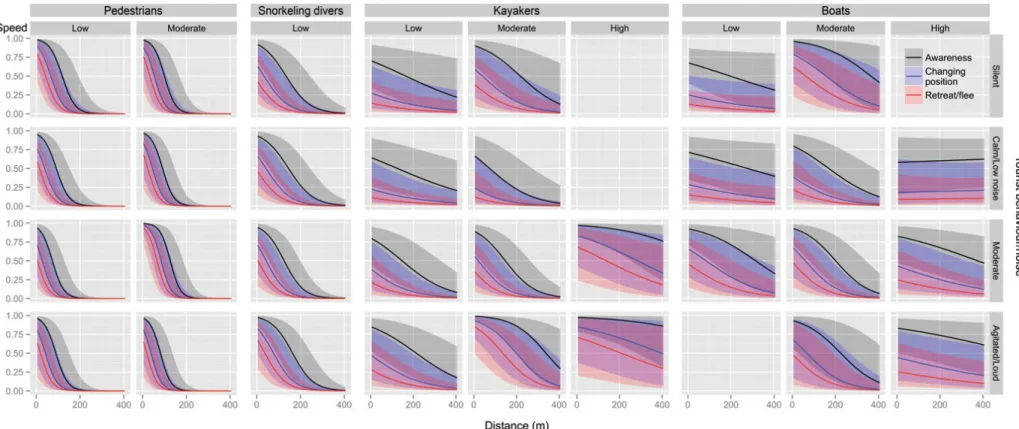
All disturbance attributes tested showed a strong influence on the response intensity of seals, with the exception of visitor group size, which was not retained in any of the models best explaining our data (Table 4 models A1-A3 and E1-E2). Pedestrian approaches were the most disturbing at short distances, considering that the probability for a seal to be disturbed was over 0.75 within 50 m under all conditions while the probability to retreat or flee was over 0.50 within 25 m under most conditions (Figure 5). Disturbances caused by vessels (boats or kayaks) were generally milder, although some conditions (silent or agitated/loud noise and moderate speed) provoked strong responses nonetheless (Figure 5). Snorkelling divers caused an intermediate disturbance level that was relatively constant across all conditions (Figure 5).
Pedestrian behaviour and speed did affect the disturbance level, as seal response was similarly high when slow moving pedestrians were silent or agitated but less intense when they were calm (Table 5; Figure 5). Furthermore, when pedestrians approached at moderate instead of low speed, the disturbance level was similar in most cases, except when pedestrian behaviour was moderate, then seal response was 9 times more likely to be stronger (Table 5; Figure 5). The latter conditions (moderate speed and behaviour) provoked the highest disturbance level for pedestrians and the highest of all close range disturbance events (within 50m). The approach distance had a strong effect on seal behaviour. A very high disturbance level was recorded within the first 50-100 m of approach and a sharp increase in the odds of an intense response as seals were 2.5-3 times more likely to be strongly disturbed when pedestrians approaching 25 m closer (Table 5; Figure 5). The predicted probabilities of displaying an anti-predatory response were not influenced by snorkelling diver behaviour (Table 5; Figure 5). However, distance from colony influenced seal behaviour, leading to a moderately-high level of disturbance within the first 100 m (Figure 5), and a response up to 1.75 times more likely to be stronger when divers approched 25 m closer (Table 5; Figure 5).
As for approaches by kayakers, the disturbance level was low when moving slowly, whereas a higher speed approach with a moderate to high level of agitation led to a seal response up to 11 times more likely to be stronger (Table 5; Figure 5). Kayaker behaviour had also an effect on seal behaviour but to a lesser extent,