ETUDE DU SITE ACTIF
DE LA CHITOSANASE DE Streptomyces sp. N174
par
Marie-Eve Lacombe-Harvey
these presentee au Departement de biologie en vue de l'obtention du grade de docteure es sciences (Ph. D)
FACULTE DES SCIENCES UNIVERSITE DE SHERBROOKE
1*1
Library and Archives Canada Published Heritage Branch BibliothSque et Archives Canada Direction du Patrimoine de l'6dition 395 Wellington Street Ottawa ON K1A 0N4 Canada 395, rue Wellington Ottawa ON K1A0N4 CanadaYour file Votre reference ISBN: 9 7 8 - 0 - 4 9 4 - 6 4 2 0 1 - 6 Our file Notre reference ISBN: 9 7 8 - 0 - 4 9 4 - 6 4 2 0 1 - 6
NOTICE: AVIS:
The author has granted a
non-exclusive license allowing Library and Archives Canada to reproduce, publish, archive, preserve, conserve, communicate to the public by
telecommunication or on the Internet, loan, distribute and sell theses
worldwide, for commercial or non-commercial purposes, in microform, paper, electronic and/or any other formats.
L'auteur a accorde une licence non exclusive permettant a la Bibliothgque et Archives Canada de reproduire, publier, archiver, sauvegarder, conserver, transmettre au public par telecommunication ou par I'lnternet, preter, distribuer et vendre des theses partout dans le monde, a des fins commerciales ou autres, sur support microforme, papier, electronique et/ou autres formats.
The author retains copyright ownership and moral rights in this thesis. Neither the thesis nor substantial extracts from it may be printed or otherwise reproduced without the author's permission.
L'auteur conserve la propriete du droit d'auteur et des droits moraux qui protege cette these. Ni la these ni des extraits substantiels de celle-ci ne doivent etre imprimes ou autrement
reproduits sans son autorisation.
In compliance with the Canadian Privacy Act some supporting forms may have been removed from this thesis.
While these forms may be included in the document page count, their removal does not represent any loss of content from the thesis.
Conformement a la loi canadienne sur la protection de la vie privee, quelques formulaires secondaires ont ete enleves de cette these.
Bien que ces formulaires aient inclus dans la pagination, il n'y aura aucun contenu manquant.
I + I
Canada
Le 14janvier 2010
le jury a accepte la these de Madame Marie-Eve Lacombe-Harvey dans sa version finale.
Membres du jury
Professeur Ryszard Brzezinski Directeur de recherche Departement de biologie
Professeure Carole Beaulieu Membre
Departement de biologie
Professeure Joelle Pelletier Membre externe
Departement de chimie, Universite de Montreal
Professeur Brian Geoffrey Talbot President rapporteur Departement de biologie
SOMMAIRE
Depuis plusieurs annees, la recherche scientifique s'est penchee sur 1'etude d'un polymere naturel nomme le chitosane. Cet interet marque pour le chitosane et ses derives, en particulier ses oligomeres, n'est pas le fruit du hasard puisqu'il touche a des applications aussi vastes que 1'agriculture, la pharmaceutique et le biomedical. L'obtention d'oligomeres de chitosane de qualite passant par l'hydrolyse enzymatique du chitosane, la recherche concernant 1'etude des chitosanases s'est done intensifiee au cours des dernieres annees. L'utilisation des chitosanases est d'ailleurs deja passee au stade industriel dans plusieurs entreprises quebecoises. Malgre ce stade industriel avance, nous ne possedons que peu de connaissances fondamentales sur la fa?on dont ces enzymes clivent le chitosane. L'objectif principal des travaux de doctorat presentes ici a ete de parfaire les connaissances concernant les rapports structure-fonction des chitosanases. Cet objectif a ete atteint en etudiant le role specifique de plusieurs residus du site actif de la chitosanase de Streptomyces sp. N174 (Csn N174), enzyme utilisee pour la production de chitooligomeres a l'echelle industrielle. Les ressemblances structurales et fonctionnelles entre la Csn N174, membre de la famille 46 des glycosides hydrolases (GH), et les chitinases et les lysozymes des families GH 19, GH 22, GH 23 et GH 24 font de cette chitosanase un modele experimental de choix pour l'acquisition de nouvelles donnees concernant les enzymes membres de toutes ces families.
Dans un premier temps, ce projet a permis d'expliquer l'activite enzymatique residuelle considerable observee chez la Csn N174 privee de son residu catalytique nucleophile (mutant D40G). II a ete suggere que, dans le contexte ou 1'aspartate 40 est substitue en glycine, un rearrangement du lobe superieur du sillon catalytique de l'enzyme permet a un autre acide amine carboxylique, le glutamate 36, de prendre le relais de la fonction de nucleophile. Cette situation peu commune serait possible grace a l'augmentation de la flexibility du lobe superieur du domaine catalytique engendree par l'ajout d'une glycine en position 40.
Dans un deuxieme temps, certains residus conserves au sein des chitosanases appartenant a la meme famille de glycosides hydrolases que la Csn N714 (GH 46) ont ete etudies en details. Les travaux realises ont permis de mettre en relief le caractere essentiel de la threonine en position 45 pour le fonctionnement efficace de l'enzyme. Ce residu serait implique dans le positionnement adequat de la molecule d'eau nucleophile participant a l'attaque des liens p-1,4 qui unissent les unites glucosamines et A^-acetylglucosamines contenues dans le chitosane. L'implication de l'arginine situee en position 42 a fait l'objet de travaux. L'etude de plusieurs mutants de cette position a demontre que l'arginine 42 contribue a la creation d'un microenvironnement electrostatique favorable a l'accomplissement de la fonction nucleophile du residu catalytique aspartate 40. Cette meme etude a permis d'etablir que cette arginine serait non seulement impliquee dans le maintien de l'integrite de la fonction catalytique, mais aussi dans le mecanisme de liaison enzyme-substrat de la Csn N174.
Dans un troisieme temps, les travaux effectuees ont mis en lumiere la participation des residus a caractere acide glutamate 197 et aspartate 201 dans le mecanisme d'attachement enzyme-substrat de la Csn N714 en plus de confirmer I'importance majeure du residu aspartate 57 dans ce mecanisme.
Dans l'ensemble, les resultats issus de ce projet de doctorat ont permis d'approfondir les connaissances sur les rapports structure-fonction de la chitosanase de Streptomyces sp. N174 et, par extension, sur les enzymes partageant des similitudes structurales et fonctionnelles avec cette derniere.
REMERCIEMENTS
Je tiens a remercier en tout premier lieu mon directeur de recherche, Dr Ryszard Brzezinski, pour avoir eu confiance en mes capacites et avoir partage avec moi sa passion pour la recherche fondamentale. Je le remercie aussi pour m'avoir guidee, conseillee, encouragee et donne 1'opportunity de voyager par le biais de la participation a des congres internationnaux.
Merci a mes conseillers, Dre Carole Beaulieu et Dr Brian G. Talbot, pour votre disponibilite et vos conseils pertinents tout au long de mes etudes graduees.
Merci au Dre Joelle Pelletier pour avoir accepte d'agir en tant que jury externe et avoir consacre du temps a la lecture de cette these.
Mes remerciements vont egalement a tous les membres du laboratoire pour les nombreuses discussions a caractere scientifique ou non... Un merci tout special a Nancy, Gabi, Melanie et Karine R. pour vos precieux conseils et surtout pour votre soutien dans les moments difficiles.
Mille merci a mes parents, Andree et Rodrigue, pour m'avoir inculque, toujours avec amour, que 1'excellence nait de 1'effort, et a mon copain Eric pour la patience qu'il a manifestee a mon egard durant mes annees d'etudes graduees.
Merci egalement aux organismes subventionnaires suivants, le Conseil de recherche en sciences et en genie du Canada (CRSNG) et les Fonds quebecois de la recherche sur la nature et les technologies (FQRNT), pour le soutien financier qu'ils m'ont accorde pour la realisation de mes etudes graduees.
TABLE DES MATIERES
SOMMAIRE ii
REMERCIEMENTS iv
TABLE DES MATIERES v
LISTE DES ABREVIATIONS vii
LISTE DES TABLEAUX ix
LISTE DES FIGURES xi
INTRODUCTION 1
LA CHITINE ET LA CHITOSANE . 1
LES GLYCOSIDES HYDROLASES 4
Generalites et systemes de classification des glycosides hydrolases 4
Structures des glycosides hydrolases 5 Principaux mecanismes d'action des glycosides hydrolases 10
Mecanismes d'action alternatifs des glycosides hydrolases 14
GENERALITES SUR LES ENZYMES IMPLIQUEES DANS LA DEGRADATION DU CHITOSANE 17
LES CHITOSANASES 18
Chitosanases de la famille 5 des glycosides hydrolases 23 Chitosanases de la famille 7 des glycosides hydrolases.... 24 Chitosanases de la famille 8 des glycosides hydrolases 25 Etudes portant sur le site actif des cellulases-chitosanases bifonctionnelles 27
Chitosanases de la famille 75 des glycosides hydrolases 28 Chitosanases de la famille 80 des glycosides hydrolases 32 Chitosanases de la famille 46 des glycosides hydrolases 33
CHITOSANASE DE Streptomyces sp. N174 4 0 Isolement, expression, production et purification 40
Mecanisme d'hydrolyse, structure tridimensionnelle et stabilite de la structure 41
Specificite de clivage et liaison au substrat 55 Autres residus impliques dans le fonctionnement adequat de l'enzyme 63
ELABORATION DES OBJECTIFS DU PROJET DE DOCTORAT ET RESULTATS PRELIMINAIRES 7 4
RESULTATS 76 CHAPITRE 1 76
1.0 Role des residus accessoires du site actif de la chitosanase de Streptomyces sp. N174 76 1.1 Accessory active site residues of Streptomyces sp. N174 chitosanase
-variations on a common theme in the lysozyme superfamily 79 1.2 Influence du degre d'acetylation du chitosane sur l'actvite enzymatique de la chitosanase de
Streptomyces sp. NI74 - type sauvage et mutant D40G 110
1.2.2 Comparaison de l'influence du degre d'acetylation du chitosane sur l'activite enzymatique-de la chitosanase enzymatique-de Streptomyces sp. N174 enzymatique-de type sauvage avec celle du mutant D40G Ill
CHAPITRE 2 ,114 2.0 Implication du residu argine 42 du site actif de la chitosanase de Streptomyces sp. N174
dans la catalyse et dans la liaison enzyme-substrat 114 2.1 A highly conserved arginine residue of the chitosanase from Streptomyces sp. N174 is
involved both in catalysis and substrate binding 116
CHAPITRE 3 146 3.0 Role des acides amines carboxyliques dans la formation du complexe enzyme-substrat chez
la chitosanase de Streptomyces sp. N174 146 3.1 Role of acidic amino acid residues in chitooligosaccharide-binding to Streptomyces sp. N174
chitosanase 149
DISCUSSION GENERALE 166
CONCLUSION 176
ANNEXE 179 Annexe 1. Alignement des sequences completes en acides amines des chitosanases de la famille GH
46 179
LISTE DES ABREVIATIONS
A : angstroms
CBM : module de liaison au carbohydrates CMC : carboxymethyl-cellulose
C°: degre Celcius
Csn N174 : chitosanase de Streptomyces sp. N174 D.A. : degre d'acetylation
D.D.A. : degre de deacetylation DC : dichroi'sme circulaire EC : Enzyme Commission g : gramme GH : glycoside hydrolases GlcN : p-D-glucosamine GlcNAc : N-acetyl-P-D-glucosamine GlcNAcases : endo-P-jV-acetylglucosaminidases GlcNases : exo-P-D-glucosaminidases HC1 : acide chlorhydrique HNO2: acide nitrique
IUB : Union Biochimique Internationale kcat '• constante catalytique
k D a : kilodalton Km: constante de Michaelis 1: litre M : molaire mg : milligramme min : minute m l : millilitre mM : millimolaire
m o l : mole mU : milliunite N : normal
NAD+ : nicotinamide adenine dinucleotide sous sa forme oxydee
NADH : nicotinamide adenine dinucleotide NAM : iV-acetylmuramique
pA'a: constante de dissociation acide PM : poids moleculaire
RMN : resonnance magnetique nucleaire RPM : revolution par minute
Tm : temperature de demi-transition de denaturation
U : unite
v/v : concentration en volume sur volume Vm a x: vitesse maximale |j.l: microlitre jaM : micromolaire p,mol: micromole a : alpha P : beta % : pourcentage
LISTE DES TABLEAUX
INTRODUCTION
1. Principales structures retrouvees chez les glycosides hydrolases 8 2. Proprietes generates des families de glycosides hydrolases comprenant les
chitosanases 21 3. Specificite de clivage du chitosane chez les chitosanases 23
4. Caracteristiques physico-biochimiques de la chitosanase de Streptomyces sp. N174 43 5. Residus potentiellement impliques dans Fattachement du substrat de la chitosanase de
Streptomyces sp. N174 58 6. Activites relatives des mutants du residu valine 148 de la chitosanase de
Streptomyces sp. N174 67
CHAPITRE 1
1. Specific activities and kinetic parameters of purified wild-type and mutant Csn N174 84 2. Thermodynamic parameters of unfolding transition of wild type and D40G
chitosanases 86 3. Key structural motifs for active site residues in the lysozyme superfamily 99
4. Degres d'acetylation et frequences relatives des liens entre les residus des
divers chitosanes utilises 111
CHAPITRE 2
I. Specific activities and kinetic parameters of purified wild-type and mutant 128 II. Specific activities toward hexaglucosamine of purified wild-type and mutant
III. Transition temperatures of thermal unfolding of wild type and mutated CsnN174
chitosanase in the absence or presence of (GlcN)3 131 IV. Specific activities of of wild type and mutated CsnN174 chitosanases towards
chitosans with different degrees of vV-deacetylation 133
CHAPITRE 3
1. Relative activities, association constants, and binding free energy changes for wild
LISTE DES FIGURES
INTRODUCTION
1. Structure de la chitine et du chitosane 1 2. Structures tridimensionnelles retrouvees chez glycosides hydrolases 7
3. Topologies des sites actifs des glycosides hydrolases 10 4. Mecanismes generaux des glycosides hydrolases 11 5. Reaction d'hydrolyse suivi de la reaction de transglycosylation des P-mannosidases
de la famille 2 des glycosides hydrolases . 13 6. Mecanisme general de catalyse assistee par le substrat 15 7. Mecanisme d'hydrolyse NAD+-dependant des glycosides hydrolases de la famille 4 16
8. Arbre phylogenique des families de glycosides hydrolases comprenant des
chitosanases 20 9. Alignement des, sequences en acides amines du site catalytique et de la partie
N- terminale des chitosanases de la famille GH 75 31 10. Alignement des sequences en acides amines des chitosanases de la famille GH 46 37
11. Arbre phylogenique des chitosanases de la famille GH 46 38 12. Alignement des sequences en acides amines des chitosanases de la famille GH 46 et
GH 80 39 13. Mecanisme d'hydrolyse de la chitosanase de Streptomyces sp. N174 43
14. Structure tridirnensionnelle de la chitosanase de Streptomyces sp. N174 45 15. Reseaux d'interactions conserves des sites actifs de quatre glycosides hydrolases
appartenant aux families 46, 24 et 19 48 16. Similitudes entre les structures tridimensionnelles de la chitosanase de Streptomyces
sp. N174 (CSN NI74), du lysozyme du bacteriophage T4 (T4L), de la chitinase de
l'orge (CHI 26) et du lysozyme du blanc d'ceuf de l'oie (GEWL) 50 17. Comparaison des surfaces moleculaires des sillons catalytique des chitosanases de
/
18. Suivi dans le temps de la degradation des hexaglucosamines par la chitosanases de
Streptomyces p. N174.. 59 19. Comparaison de la densite des charges de surface de la chitinase de l'orge et de la
chitosanase de Streptomyces sp. N174 60 20. Alignement des sequences partielles de la chitosanase de Streptomyces sp. N174 et de
la chitinase de l'orge 63 21. Gros plan du site catalytique de la chitosanase de Streptomyces sp. N174 64
22. Gros plan n°2 du site catalytique de la chitinase de l'orge 70 23. Gros plan n° 3 du site catalytique de la chitosanase de Streptomyces sp. N174 72
24. Gros plan n°4 du site catalytique de la chitosanase de Streptomyces sp. N174 73
CHAPITRE 1
1. Thermal unfolding curves of the wild type (a) and D40G (b) chitosanases in the
presence or absence of (GldSTb 85 2. Time-courses of (GlcN)5 and (GlcN)4 hydrolysis catalyzed by wild type and D40G
endochitosanases monitored by real-time mass spectrometry 87 3. Anomer production from the D40G mutant chitosanase hydrolysis of (GlcN)6.
(a) Time-dependent 'H-NMR spectra, (b) Time-course of anomer production 88 4. Chemical rescue experiments.
(a) Time-courses of the enzymatic degradation of (GlcN)6 by D40G in the absence or
presence of sodium azide (0.65 and 2.6 M) 89 5. A) Structural view of the active site cleft of chitosanase Csn-N 174 94 92
5. B) Alignment of portions of the primary structure of GH46 chitosanases including
active site residues 93 6. Time-courses of (GlcN)6 hydrolysis catalyzed by wild type and
D40G+T45D chitosanases monitored by real-time mass spectrometry 94 7. Influence de la frequence des liens GlcN-GlcN contenus dans le chitosane sur
CHAPITRE 2
1. Structural view of a part of the upper hinge of the active site cleft
of chitosanase Csn-N174 125 2. Alignment of portions of the primary structure of experimentally active-proven GH46
chitosanases including active site residues . 126 3. Specific activity of Csn N174 R42X mutants from crude extract of E. coli cells 127
4. Comparison of pH-activity profiles of Csn N174 WT, R42E and R42K 130 5. Thermal unfolding curves of R42K (A) and R42E (B) chitosanases in the absence or
presence of (GlcN)3 133 6. Effect of the degree of 7V-deacetylation of chitosan on wild-type and Arg42-mutated
chitosanases actvity. A value of 100% was attributed to chitosanase activity toward
chitosan Sigma (82.5% -deacetylated) 134 7. Time-courses of (GlcN)6 hydrolysis catalyzed by wild-type, R42K and R42E D40G
chitosanases monitored by real-time mass spectrometry 135 8. Cleavage frequencies proportions of wild-type, R42E and R42K chitosanases at pH
5.2 calculated from data on Figure 6 137
CHAPITRE 3
1. X-ray crystal structure of Streptomyces sp. N174 chitosanase 152 2. Fluorescence spectra of wild type chitosanase from Streptomyces sp. N174 in the
presence of GlcN (A), (GlcN)2 (B), and (GlcN)3 (C) 156
3. Scatchard plots of the fluorescence quenching induced by oligosaccharide binding to
the wild type chitosanase 158 4. Saturation binding curves of the wild type, D57A, E197A, and D201A 160
5. Scatchard plots of the fluorescence quenching induced by oligosaccharide binding to
DISCUSSION GENERALE
25. Alignement des sequences en acides amines du site actif des chitosanases de la
famille GH 46 168 26. Alignement structurel des sequences en acides amines du site actif de la Csn N174 et
de trois chitinases appartenant a la famille 19 des glycosides hydrolases 170 27. Cladogramme simplifie illustrant les relations evolutives entre les membres de la
I N T R O D U C T I O N
L A C H I T I N E ET LE C H I T O S A N E
Au cours des dernieres annees, la recherche scientifique s'est penchee sur un polymere naturel nomine le chitosane, obtenu a partir de la chitine, second biopolymere en abondance sur la Terre, apres la cellulose. L'abondance de la chitine n'est pas surprenante, puisque cette derniere constitue la principale composante de la carapace des crustaces, de l'exosquelette des insectes et de la paroi cellulaire de certains champignons. A la difference de la chitine, polymere compost presque exclusivement d'unites N-acetylglucosamines (GlcNAc) reliees par des liens glycosidiques P-1,4, le chitosane contient aussi des unites glucosamines (GlcN) (figure 1).
(a)
(b)
Figure 1. Structure (a) de la chitine et (b) du chitosane.
De ce fait, la deacetylation (complete ou partielle) de la chitine permet d'obtenir du chitosane de divers degres d'acetylation et etalant des proprietes differentes. La presence de charges positives sur le groupement amine des unites glucosamines (a un pH inferieur a 6,5) fait du chitosane un polymere cationique et permet sa solubilisation a des pH legerement acides. Ces deux dernieres proprietes le distinguent de la chitine et font du
chitosane un compose unique et plus interessant au point de vue des applications industrielles. En effet, les charges positives du chitosane lui permettent de reagir avec des composes biologiques charges negativement, tels que les graisses, les proteines et les enzymes. D'ailleurs, le chitosane est deja utilise depuis plusieurs annees dans plusieurs industries oeuvrant dans des domaines aussi vastes que 1'agriculture, le traitement des eaux usees, la cosmetique, l'alimentaire, la biopharmaceutique ainsi que le biomedical (Hadwiger, 1999 ; Choi et al., 2004). Devant toutes ces applications, il n'est done pas surprenant que la recherche concernant le chitosane et ses derives soit, encore aujourd'hui, en plein essor.
Une des proprietes les plus interessantes du chitosane est son action antimicrobienne. II est effectivement demontre que le chitosane inhibe la croissance de certaines bacteries telles Escherichia coli et Staphylococcus aureus (Hwang et al., 1998 ; Cuero et al., 1999). Deux principaux mecanismes sont ainsi proposes pour expliquer la toxicite du chitosane vis-a-vis des microorganismes. Dans le premier mecanisme, la nature cationique du chitosane serait responsable de son activite antimicrobienne. Selon l'hypothese emise par
I
Helander et al. (2001), le chitosane interfere avec les macromolecules a charges negatives presentes a la surface des cellules, par exemple les lipopolysaccharides. Cette interaction avec la membrane resulte en une disorganisation de cette derniere provoquant une alteration de la permeabilite membranaire et, par consequent, une perte de la fonction de barriere membranaire. De la, il advient une sortie, un relargage, de certains constituants cellulaires, tels que les proteines et les electrolytes, entrainant la mort du microorganisme. Des travaux plus recents axes sur la morphologie de l'integrite de la membrane cellulaire et de la permeabilite des membranes externe et plasmique de E. coli et de S. aureus en presence de chitosane semblent confirmer et preciser cette hypothese (Liu et al., 2004); les interactions electrostatiques entre les groupements (NH3)+ du chitosane et les
groupements phosphoryles des phospholipides des membranes bacteriennes seraient responsables en majeure partie de la mort des bacteries via un relargage du contenu cellulaire. Le second mecanisme propose plutot un mode d'action via une liaison du chitosane avec l'ADN. Selon cette hypothese, certaines molecules de chitosane de petits poids moleculaires penetreraient dans la cellule, se fixeraient ensuite a l'ADN entravant
la transcription des genes et, de cette fa9on, empecheraient la synthese de proteines, ce qui provoquerait la mort du microorganisme (Tsai et Su, 1999). II est important de mentionner que l'activite antibacterienne du chitosane est fonction de plusieurs parametres, tels que le degre d'acetylation (D.A.) du chitosane, son poids moleculaire, le pH, la temperature et la concentration en sels du milieu (Liu et ah, 2006 ; Fujimoto et ah, 2006). De surcroit, il a ete demontre au cours des dernieres annees que le chitosane et les oligomeres issus de sa degradation agissent efficacement comme composes antifongiques et comme eliciteurs des mecanismes de defense des plantes en induisant la production de proteines reliees a la pathogenese (Rabea et al; 2003 Choi et ah, 2004).
Plusieurs equipes de recherche se sont penchees sur le potentiel pharmaceutique et medical du chitosane et de ses derives. A ce propos, une recente etude clinique (Jaffer et
Sampalis, 2007) demontre l'efficacite d'hydrolysats de chitosane dans le controle de l'hypercholesterolemie. Parallelement, d'autres travaux confirment son potentiel en tant qu'agent antitumoral (Harish Prashanth et Tharanathan, 2005) et en tant qu'activateur de la reponse immunitaire (Okawa et ah, 2003). Tout comme dans le cas de l'activite antibacterienne du chitosane mentionne precedemment, la longueur de la chaine de chito-oligosaccharides de meme que son degre d'acetylation exercent vine influence substantielle sur l'activite biologique du chitosane. Consequemment, la mise en place de methodes permettant un controle rigoureux a la fois de la longueur de la chaine et du degre d'acetylation s'avere une condition sine qua non a la production d'oligomeres de chitosane de valeur. Bien que l'hydrolyse par un acide (HC1 ou HNO2) soit une methode relativement peu onereuse pour cette production, le faible rendement en terme d'oligosaccharides produits, l'heterogeneite des produits, la toxicite des acides utilises, en particulier du HNO2, la presence d'acides forts residuels et de sous-produits de reaction limitent son utilisation specialement en ce qui a trait a ses applications dans les domaines pharmaceutiques et alimentaires (Rupley, 1964 ; Choi et ah, 2004 ; Su et ah, 2006). Pour obtenir des oligomeres de chitosane de haute qualite et biocompatibles ayant les proprietes desirees, l'hydrolyse par le biais d'enzymes s'avere la methode de choix. Dans le cas present, cette hydrolyse est assumee par des enzymes nommees les chitosanases.
L E S G L Y C O S I D E S H Y D R O L A S E S
Generalites et systemes de classification des glycosides hydrolases
Les chitosanases font partie d'un vaste groupe d'enzymes capables d'hydrolyser les liens glycosidiques existant chez les divers polysaccharides. Ces enzymes, nominees les glycosides hydrolases ou glycosidases, sont tres diversifies et retrouvees pratiquement chez toutes les formes de vie. Cette diversite s'explique a la fois par 1'importance du metabolisme des sucres et par l'innombrable variete de polysaccharides existant dans la nature. A titre d'exemple, le nombre d'isomeres possibles pour un simple hexasaccharide
12
est evalue a 10 . Aussi, les glycosides hydrolases jouent un role crucial dans plusieurs reactions biologiques essentielles, telles que la degradation des polysaccharides de structure ou de reserve, la defense contre les agents pathogenes, la signalisation cellulaire et le renouvellement des polysaccharides a la surface de la cellule (Davies et Henrissat, 1995). Ces enzymes ont ete, en premier lieu, classifies selon le systeme de l'Enzyme Commission (EC), soit la nomenclature adoptee par l'Union Biochimique Internationale (IUB). Dans ce systeme conventionnel, les chitosanases possedent le code suivant : EC.3.2.1.132. Les trois premiers chiffres designent une enzyme hydrolysant les liens O-glycosides, tandis que le dernier chiffre definit le type de substrat. Le nombre imposant d'enzymes appartenant aux glycosides hydrolases a mene a l'elaboration d'un systeme de classification specialement con^u pour ces dernieres. Cette nomenclature, proposee par Bernard Henrissat en 1991, basee sur la sequence en acides amines, regroupe les glycosides hydrolases en differentes families. De plus, elle se veut plus representative de la specificite plus ou moins grande pour plusieurs substrats de ces enzymes. Contrairement a la classification EC, la classification par famille (GH) rend compte de revolution convergente ou divergente empruntee par ces enzymes et rend realisable un regroupement d'enzymes de plusieurs families en clans possedant une structure tridimensionnelle et un mecanisme d'action conserves (Davies et Henrissat, 1995). Les glycosides hydrolases fonctionnent via deux mecanismes distincts d'hydrolyse du substrat: le mecanisme a retention anomerique ou le mecanisme a inversion anomerique (les details des mecanismes catalytiques des glycosides hydrolases seront discutes
ulterieurement). Cette derniere caracteristique peut done etre utilisee comme critere de classification et est maintenue au sein d'une meme famille. A ce jour, les glycosides hydrolases comprennent plus de 115 families, dont six comprennent des chitosanases. Une des prerogatives importantes de ce systeme est qu'il permet d'extrapoler les connaissances acquises sur une enzyme en particulier, entre autres, le type de mecanisme d'hydrolyse aux autres enzymes de la famille, voire aux membres d'un meme clan. Quatorze clans de glycosides hydrolases, identifies par les lettres de A a N, sont recenses a ce jour. A titre d'exemple, les families de GH 8 et 48 sont regroupees sous un seul clan, le clan GH-M, puisqu'elles possedent toutes une structure de type (a/a)6 ainsi qu'un mecanisme de clivage dit a inversion anomerique. Au cours des dernieres annees, cette classification a d'ailleurs ete appliquee a d'autres groupes d'enzymes reliees au metabolisme des sucres, notamment aux transglycosylases, aux glycosyltransferases, aux polysaccharides lyases et aux esterases degradant les hydrates de carbone (Henrissat et Davies, 2000). Bien qu'ils ne soient pas des enzymes, les modules de liaison au carbohydrates (CBM) sont aussi regroupes sous cette classification. II est aussi possible de pousser plus loin le regroupement des glycosides hydrolases en reunissant les families ayant des ressemblances au niveau de leurs structures tertiaires en superfamilles. Contrairement aux families d'un clan, les membres d'une superfamille ne partagent pas necessairement le meme type de mecanisme d'hydrolyse et ne sont done pas toujours proches parents les uns des autres d'un point de vue evolutif.
Structures des glycosides hydrolases
D'un point de vue structural, les glycosides hydrolases etalent un large eventail de types de reploiements proteiques {protein fold) (figure 2). L'existence d'un si grand nombre de structures differentes aptes a catalyser une meme reaction, en Voccurrence l'hydrolyse de liens glycosidiques, temoigne de la versatility des glycosides hydrolases. A titre d'exemple, l'hydrolyse du chitosane peut etre accomplie par des chitosanases appartenant a trois differents types structuraux: P+a, (p/a)g et (a/a)6. Inversement, des glycosidases detenant un meme type de structure peuvent hydrolyser des substrats differents. Cette caracteristique s'illustre parfaitement par la polyspecificite des substrats reconnus par les
enzymes de structure (p/a)s. En effet, ces dernieres comprennent des cellulases, des glucanases, des mannanases, des licheninases, des chitosanases, des xylanases, et ce pour n'en citer que quelques-unes.
La structure tridimensionnelle la plus repandue parmi les glycosides hydrolases est le tonneau (p/a)g ou TIM barrel (en reference a l'isomerase de triose phosphate (TIM), enzyme pour laquelle cet arrangement structural fut observe pour la premiere fois). Le tonneau consiste en une alternance de huit brins P paralleles et de huit helices a le long de la chaine polypeptidique. Au cours du reploiement de la proteine, les brins P se disposent de fa<?on a former le centre du tonneau, alors que les helices a viennent border le pourtour du tonneau (figure 2a). La composition en acides amines des brins P confere souvent au coeur du tonneau, site catalytique de 1'enzyme, un caractere hydrophobe. Cette structure est observee chez pres d'une trentaine de families de glycosides hydrolases dont celles appartenant aux clans A, D, H et K (tableau 1), ainsi que chez les families 14, 29, 31, 67 et 84. Le tonneau en « roule suisse P » ou (3-jelly roll barrel est un autre type structural retrouve chez les glycosidases (figure 2b). Forme par quatre paires de brins P antiparalleles enroules de maniere a former un tonneau, cet arrangement structural est beaucoup moins frequent que celui en tonneau (p/a)g et n'est retrouve que chez cinq families de glycosides hydrolases (GH 7, 11, 12, 16 et 54) parmi lesquelles figurent celles reliees aux clans B et C (tableau 1). La turbine P ou P-propeller est, a l'instar du tonneau en « roule suisse P », une structure composee essentiellement de brins p. Observee au sein des families membres des clans E, F, J, et de la famille 74, cette structure se caracterise par la presence de quatre a huit feuillets P en forme de pale arranges symetriquement autour d'un axe central (tableau 1). Chacune de ces pales est generalement composee de quatre brins p antiparalleles replies de maniere a rendre le premier brin perpendiculaire au quatrieme brin (figure 2c, d et e). Contrairement aux trois types de structures decrits precedemment, certaines glycosides hydrolases revel ent une architecture composee presque exclusivement d'helices a . En effet, les enzymes des families 9, 88 et 105 de meme que les families relevant des clans G, L et M montrent une structure tridimensionnelle dite en tonneau (a/a)6 (tableau 1).
Figure 2. Structures tridimensionnelles retrouvees chez glycosides hydrolases : (a) tonneau (p/a)g; (b) tonneau en roule suisse P ; (c) turbine p a 5 feuillets; (d) turbine P a 6 feuillets; (e) turbine p a 7 feuillets; (f) tonneau (a/a)6; (g) tonneau ( a / a )7 (h) helice
P de pas droit; (i) dehydrogenase/dehydratase-like et (j) «+P (Figure tiree de Hancock et Withers, 2007)
Tableau 1. Principales structures retrouvees chez les glycosides hydrolases Structure ou
type de reploiement proteique Clans de GH Families de GH
tonneau (P/a)s GH-A 1, 2, 5, 10, 17, 26, 30, 35, 39, 42, 50, 51,53,59, 72, 79, 86 et 113 tonneau (P/a)s GH-D 27,31 et 36 tonneau (P/a)s GH-H 13, 70 et 77 tonneau (P/a)s GH-K 18, 20 et 85 tonneau (P/a)s 14, 26, 31, 67 et 84 tonneau en « roule suisse P »
GH-B 7 et 16
tonneau en « roule suisse P » GH-C 1 l e t 12 tonneau en « roule suisse P »
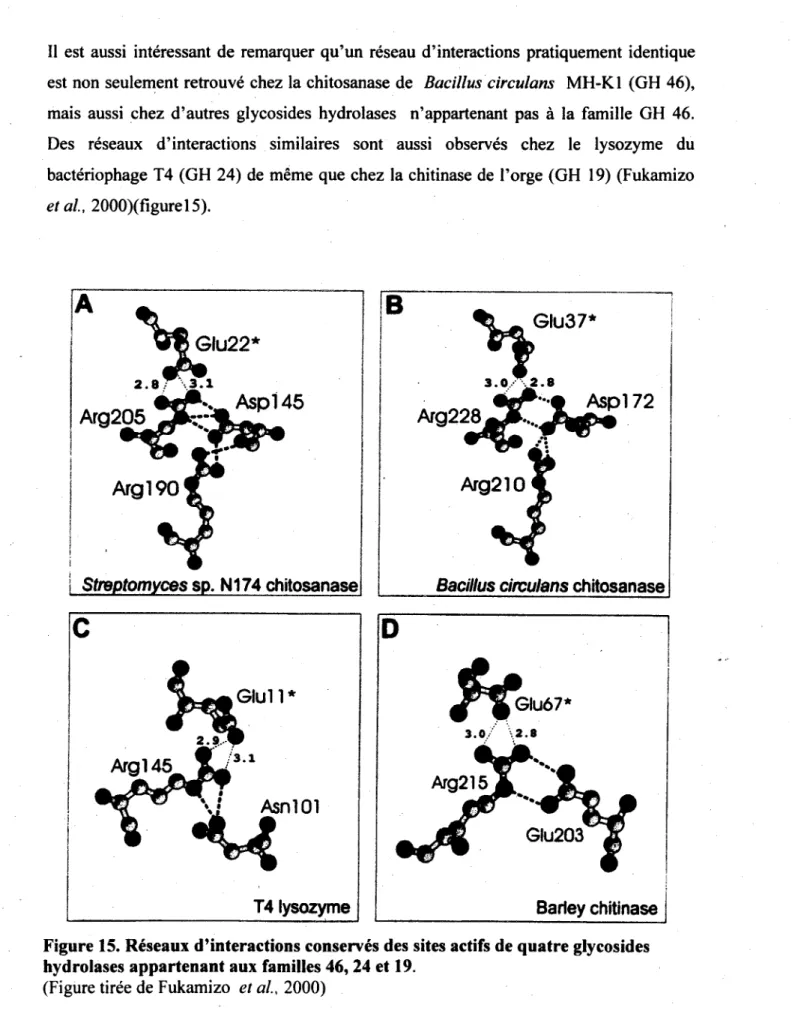
54 turbine P GH-E 33, 34 et 83 turbine P GH-F 43 et62 turbine P GH-J 32 et 68 turbine P 74 tonneau (a/a)6 GH-G 37 et 63 tonneau (a/a)6 GH-L 15 et 65 tonneau (a/a)6 GH-M 8 et 48 tonneau (a/a)6 9, 88 et 105 helice P GH-N 28 et 49 helice P 82 et 90 dehydrogenase/dehydratase-like 4 a + p GH-I 24, 46 et 80 a + p 19, 22 et 23
Ce type de tonneau est constitue d'une premiere serie de six helices a paralleles, formant les douves interieures du tonneau, entouree par une seconde serie de six autres helices a (figure 2f). Une variation peu commune de cette structure, le tonneau (i01/01)7, est rencontre chez les glycosides hydrolases de la famille 47 (figure 2g). Les enzymes appartenant au clan N de meme que celles des families 82 et 90 possedent une structure peu commune appelee helice [3 (tableau 1). Cet arrangement structural se caracterise par une suite de brins p paralleles s'enroulant autour d'eux-memes et produisant ainsi un motif helicoi'dal de pas droit (figure 2g). Cette structure semble n'etre utilisee que chez des glycosidases hydrolysant des polysaccharides polyanioniques tel 1'acide pectique. Une structure tridimensionnelle singuliere distingue les membres de la famille 4 des autres glycosides hydrolases. Cette structure est qualifiee de dehydrogenase/dehydratase-like (en reference a celle retrouvee typiquement chez les dehydrogenases et les
dehydratases) et comprend deux series de pi is de Rossman (succession d'au moins trois brins P paralleles joints par deux helices a creant un motif d'elements secondaires de type p-a-p-a-p). La presence de ces pi is de Rossman permet au nicotinamide adenine dinucleotide (NAD(H)), un cofacteur, de s'attacher adequatement aux enzymes de la famille 4. Le dernier type structural observe parmi les membres des glycosides hydrolases est la structure a+p. Cette derniere se caracterise par un assemblage, sans motif particulier, de brins P antiparalleles et d'helices a. Les chitinases, les chitosanases et les lysozymes des families 19, 22, 23 et du clan I partagent un noyau a+p invariable, nomme noyau typique au lysozyme (lysozyme core), fait de deux helices a et d'un feuillet P a trois brins servant a la fois de site catalytique et de site de liaison au substrat (Hancock et Withers, 2007 ; Cantarel et al, 2009 ; Coutinho et Henrissat, 1999 ; Carbohydrate Active Enzymes database (http://www.cazy.org/)).
Topologies des sites actifs des glycosides hydrolases
II existe trois differents types de structures tridimensionnelles du site catalytique (topologies) chez les glycosides hydrolases : le cratere ou la poche (crater or pocket), le sillon (cleft or groove) et le tunnel {tunnel) (figure 3). La premiere topologie, celle nommee cratere, est optimale pour la reconnaissance de l'extremite non reductrice du sucre et comprend alors principalement des exopolysaccharidases telles que les glucoamylases, les P-galactosidases, les P-glucosidases et les P-amylases. La seconde topologie, le sillon, est une structure dite ouverte autorisant un attachement aleatoire de plusieurs unites de polymeres. Cette topologie maximise l'affinite d'attachement pour les longs substrats polymeriques et est frequemment retrouvee chez les endopolysaccharidases comme les lysozymes, les endocellulases, les chitinases, les chitosanases, les a-amylases, les xylanases, les P-1,3-1,4-glucanases et les P-1,3 glucanases. La derniere topologie, celle nommee tunnel, se retrouve chez les cellobiohydrolases ainsi que chez les p-l,6-glucanases. Cette structure particuliere permet a ces enzymes de liberer le produit d'hydrolyse tout en demeurant attachees au substrat polysaccharidique creant alors les conditions pour la processivite (Davies et Henrissat, 1995 ; Hancock et Withers, 2007).
Figure 3. Topologies des sites actifs des glycosides hydrolases : (a) cratere; (b) sillon et (c) tunnel. La position des acides amines catalytiques est illustree en rouge. (Figure tiree de Hancock et Withers, 2007)
Principaux mecanismes d'action des glycosides hydrolases
Les glycosides hydrolases clivent les liens glycosidiques via deux types generaux de mecanismes distincts: le mecanisme a retention anomerique ou le mecanisme a inversion anomerique. Tel que mentionne precedemment, il n'existe qu'un seul type de mecanisme pour chaque famille. Decrits pour la premiere fois en 1953 par Daniel Koshland, ces mecanismes impliquent tous deux une catalyse acide necessitant un donneur de proton et un nucleophile/base, un ion oxocarbenium a l'etat de transition, ainsi qu'une paire d'acides carboxyliques au site actif, mais ils different par la configuration des produits generes (Koshland, 1953 ; Rye et Withers, 2000).
Le mecanisme a inversion anomerique opere par un deplacement direct d'une molecule d'eau situee entre le substrat et l'acide amine catalytique (ou un residu carboxylique tient le role d'un acide et l'autre d'un nucleophile) provoquant alors une inversion de la configuration anomerique (figure 4a). Ce type d'hydrolyse est accompli par deux reactions concomitantes : une protonation du substrat par le residu dit acide, ce qui permet de liberer le groupe partant du sucre, et une attaque nucleophile realisee par une molecule d'eau activee par le residu dit nucleophile. Dans ce cas, l'ion oxocarbenium est issu de la liaison d'une molecule d'eau placee entre le residu catalytique nucleophile et le
carbone CI du substrat. Une hydrolyse d'un lien glycosidique P generera done une produit avec une conformation a ou vice et versa.
(a) (b) 105A
t
S
-q r
l R S.S ;(
Y H O r^o Oh - c A o H O R r . - c r ^ oFigure 4. Mecanismes generaux des glycosides hydrolases : (a) mecanisme a inversion anomerique; (b) mecanisme a retention anomerique. (Figure tiree de Zechel et Whiters, 2000).
Le second type de mecanisme, mecanisme a retention anomerique, procede via un mecanisme a double deplacement ou un residu carboxylique agit comme nucleophile alors que l'autre agit a la fois comme acide et base (figure 4b). Au cours de la premiere etape, dite etape de glycosylation, le groupement carboxylique du residu acide/base agit comme un acide et entraine la protonation de l'atome d'oxygene du lien glycosidique correlativement au clivage du lien glycosidique, tandis que le residu nucleophile attaque
le carbone anomerique du sucre. Un intermediaire covalent enzyme-substrat est alors obtenu. Cet intermediaire est forme via un lien covalent entre le residu catalytique nucleophile et le carbone en CI du substrat. Lors de la seconde etape, dite etape de deglycosylation, le groupement carboxylique du residu acide/base agit comme une base et soustrait un proton d'une molecule d'eau entrante, laquelle attaque a son tour le centre anomerique de 1'intermediaire enzyme-substrat provoquant ainsi la rupture du lien covalent enzyme-substrat (Zechel et Withers, 2000). Dans ce cas, le substituant hydroxyle en CI conserve la meme stereochimie que le substrat (Rye et Withers, 2000).
Dans les deux mecanismes, 1'acide amine dit donneur de protons, se situe a l'interieur d'une distance d'un lien hydrogene par rapport a l'oxygene du glycoside, soit autour de 2 angstroms ( A ) . Pour le residu dit nucleophile/base, il se retrouve dans l'environnement immediat du carbone anomerique du sucre dans le cas des enzymes a retention, alors qu'il doit se situer plus loin pour accommoder la presence d'une molecule d'eau entre le substrat et le sucre chez les enzymes a inversion. Par voie de consequence, la distance entre les deux acides amines catalytiques est plus grande chez les enzymes a inversion que chez celles a retention, soit respectivement, 9.5 A versus 5 A en moyenne (figure 4). II est aussi interessant de remarquer que, chez la plupart des glycosides hydrolases, les acides amines catalytiques, generalement le glutamate ou l'aspartate, sont situes dans des regions conservees pour ce qui est de leurs sequences acides amines (Henrissat et Bairoch,
1993), ce qui soutient l'implication de plusieurs autres acides amines dans I'efficacite du fonctionnement de ces enzymes.
Une particularity interessante des enzymes procedant via le mecanisme a retention est la possibility pour ces dernieres d'effectuer un second type de reaction enzymatique (figure 5). Cette seconde activite, nommee la transglycosylation, catalyse un transfert de groupement glycosidique parallelement au clivage d'un lien glycosidique. Cette catalyse s'effectue sans l'utilisation d'une molecule d'eau (ce n'est done pas une hydrolyse) et ne genere pas d'eau comme produit (ce n'est done pas une hydrolyse inverse). Ce phenomene se manifeste lorsque que la concentration en produits generes par la premiere (l'hydrolyse) reaction est elevee. De plus, les conditions optimales (temperature, force
ionique, par exemple) pour la reaction de transglycosylation sont differentes de celles de la reaction initiale d'hydrolyse (Palcic, 1999).
Figure 5. Reaction d'hydrolyse (haut) suivi de la reaction de transglycosylation (bas) des P-mannosidases de la famille 2 des glycosides hydrolases. (Figure tiree de Stoll et al., 2000)
II est aussi a noter que le nouveau lien engendre par la transglycosylation peut etre ou non du meme type que celui initialement hydrolyse. II resulte de tout ceci que la transglycosylation permet la synthese d'oligosaccharides differents et de poids moleculaire egal ou plus haut que celui des oligosaccharides de depart. Bien que la transglycosylation soit consideree comme une activite secondaire, son importance dans la regulation du metabolisme des sucres n'est toutefois pas negligeable. Chez E. coli, 1'expression de l'operon lactose est regulee, en partie, par le produit genere par la reaction de transglycosylation de la P-galactosidase (GH 2). En effet, une partie du glucose et du galactose engendree par l'hydrolyse du lien P-1,4 du lactose par la p-galactosidase est transformee, via la formation d'un lien P-1,6 par transglycosylation, en allolactose, un inducteur naturel de l'expression de l'operon lac (Juers et al., 2001). II est aussi interessant de souligner que, depuis quelques annees, certaines endoglycosidases, ayant une activite de transglycosylation elevee, sont utilisees pour la synthese de composes glycosyles bioactifs.
A titre d'exemple, les endo-P-A'-acetylglucosaminidases (GlcNAcases) de Mucor hiemalis (Endo-M) et de Arthrobacter protophormiae (Endo-A), toutes deux membres de la famille GH 85, sont employees dans la fabrication de glycopeptides synthetiques, tels que les formes glycosylees de la calcitonine, hormone cle dans la regulation du taux de calcium sanguin des mammiferes (Wang et al, 2004 ; Hadena et al., 2006). Outre les GlcNAcases de la famille GH 85, plusieurs autres enzymes impliquees dans la degradation de la chitine et du chitosane, telles que les chitinases de la famille GH 18 et les exo-P-D-glucosaminidases (GlcNases) de la famille GH 2, sont capables de transglycosylation (Sasaki et al., 2002 ; Cote et al., 2006).
Mecanismes d'action alternatifs des glycosides hydrolases
Bien que la majorite des glycosides hydrolases operent via un des deux mecanismes decrits precedemment (inversion ou retention anomerique), quelques-unes font exception. Par exemple, chez les sialidases et des neuraminidases des families GH 33 et GH 34, la fonction de nucleophile (habituellement assuree par un aspartate ou un glutamate), est
assuree par une tyrosine activee (pour une attaque nucleophile) par un aspartate adjacent. Cette substitution de nucleophile permet de contourner un probleme rencontre chez les enzymes ayant le sialoside comme substrat ; la presence d'une charge negative (due a la presence d'un groupement carboxyle) adjacente au centre anomerique rend defavorable une attaque nucleophile par un residu anionique. Le cas des myrosinases de la famille GH 1, enzymes chvant les thioglycosides anioniques chez les plantes, fait aussi exception. Afin de minimiser la repulsion entre le substrat et le site catalytique de 1'enzyme, le residu glutamate dit acide/base est substitue par une glutamine. La nature reactive des substrats thioglycosides permet la formation de 1'intermediaire enzyme-substrat par catalyse acide et ce, malgre l'absence du residu catalytique dit acide. Un residu catalytique ayant la fonction de base est cependant necessaire pour 1'etape de deglycosylation. Pour pallier a ce probleme, les myronidases utilisent une molecule d'acide ascorbique (lie a l'enzyme via un site de liaison specifique) pour assurer le role de la base catalytique. Ce type d'hydrolyse conserve la conformation anomerique initiale du substrat et est done considere comme une variante du mecanisme a retention anomerique classique. Un autre type de mecanisme alternatif, un derive du mecanisme a retention anomerique, est observe chez les N-acetyl-hexoaminidases, les chitinases et les hyaluronidases des families GH 18, 20, 56 et 84. Chez ces enzymes, le residu catalytique nucleophile est remplace par le groupement N-acetyl (acetamido) en position 2 du substrat meme (figure 6). Le lien glycosidique est ensuite scinde, par catalyse acide, via la formation d'un intermediaire de reaction, nomme ion oxazolinium. Le lien cree lors de la formation de l'ion oxazolinium est ensuite hydrolyse par une molecule d'eau activee par un residu carboxylique (agissant comme une base) du site actif de l'enzyme (Dall'aqua et Carter, 2000 ; Rajan et al., 2004). Ce type de mecanisme d'hydrolyse est couramment appele « catalyse assistee par le substrat » (substrate-assisted catalysis).
e o ' ^ o H0"^v0 ROH ' n ( ^ H H — Aro ' W ^ V - O H r\ -H NHAc Hp.>0 CH3
Figure 6. Mecanisme general de catalyse assistee par le substrat. Exemple de la N-acetyl-p-hexoaminidases (Figure tiree de Zechel et Withers, 2000).
Tel que mentionne precedemment, les enzymes appartenant a la famille GH 4 possedent une structure tridimensionnelle unique parmi les glycosides hydrolases. Or, cette singularity se reflete aussi en ce qui a trait au mecanisme d'hydrolyse de ces enzymes. Une premiere particularity de la famille GH 4 vient du fait qu'elle regroupe des enzymes specifiques aux a glycosides et aux {3 glycosides. De plus, la presence du NAD+ et d'ions
| I 0-1- I
divalents (Mn , Ni , Coz ou Fe ) est essentielle au fonctionnement des enzymes
membres de cette famille. II est a noter que le NAD+ n'est pas consomme durant la
catalyse. Des etudes structurales et mecanistiques detaillees ont revele que ces enzymes utilisent un mecanisme d'hydrolyse sequentiel de type oxydation-elimination-addition-reduction faisant intervenir des etats de transition anioniques, ce qui s'oppose aux etats de transition cationiques normalement observes au cours de la catalyse par deplacement direct ou de celle par double deplacement. Comme le montre la figure 7, la premiere etape de la reaction d'hydrolyse consiste en la formation d'une cetone en position C3 suite a l'oxydation du groupe hydroxyle par le NAD+. Ceci a pour consequence de rendre
le proton present en C2 du substrat plus acide, permettant ainsi, avec l'aide d'un residu carboxylique agissant comme base, le clivage du lien entre le CI et le 0 1 par le biais d'une reaction d'elimination. Une molecule d'eau est ensuite ajoutee au centre anomerique par une addition de Michael-1,4, suivi de la reprotonation du carbone en position 2. Enfin, la cetone en C3 est reduite par le NADH transitoire, produit a la suite de l'etape d'oxydation, ce qui a pour effet de regenerer l'etat initial de l'enzyme (Rajan et al., 2004 ; Hancock et Withers, 2007).
I l l :
OB HB SB GE
Hn BOH XOA / u Hrt Ho
* ^.. OR V J-A o ^ ^ • V" > -Y--OH ^ ^ 4-OH
!h ! ih — ' ry > v v r ^y-NH,
NADH NAD*
Figure 7. Mecanisme d'hydrolyse NAD+-dependant des glycosides hydrolases de la
famille 4. (X=P032" ou H). (Figure tiree de Zechel et Withers, 2000). •A.-OR
\ /H °H
1 \ H O
Selon une etude recemment publiee par Liu et al. (2007), une a-N-acetylgalactosaminidase de la famille GH 109 utiliserait un mecanisme similaire a celui des enzymes de la famille GH4.
G E N E R A L I T E S S U R L E S E N Z Y M E S I M P L I Q U E E S D A N S L A D E G R A D A T I O N DU C H I T O S A N E
Le fait que le chitosane soit compose de deux monomeres en proportions largement inegales donne lieu a plusieurs types de liens entre ces divers residus. En effet, le chitosane partiellement acetyle contient, respectivement, deux types de liens dits « hetero » et deux types de liens dits « homo » : GlcNAc-GlcN, GlcNAc, GlcN-GlcN et GlcN-GlcNAc-GlcN-GlcNAc (Fukamizo et Brzezinski, 1997). L'hydrolyse des trois premiers types de liens est assuree par les chitosanases. Ces dernieres sont des endo-enzymes, c'est-a-dire qu'elles hydrolysent aleatoirement les liens presents a l'interieur de la chaine de chitosane. En tant qu'exception a ce qui precede, les travaux de Cote et al. (2006) ont recemment demontre l'existence, d'une exochitosanase chez l'actinomycete Amycolatopsis orientalis. Cette exochitosanase est en fait une exo-p-D-glucosaminidase qui peut degrader, outre les residus terminaux des oligomeres de GlcN, les residus GlcN se trouvant a l'extremite non reductrice des chaines de chitosane de hauts poids moleculaires. Le quatrieme et dernier type de lien, celui unissant deux GlcNAc consecutifs, n'est pas reconnu par les chitosanases. Le clivage de ce type de lien est assure par un autre groupe d'enzymes nommees les chitinases. Tout comme les chitosanases, la majorite des chitinases sont des endo-enzymes. II convient de preciser ici que les deux types de liens dits « hetero » presents au sein du chitosane peuvent aussi etre reconnus et clives par les chitinases. En ce sens, il est possible de dire que les specificites de substrats des chitinases et des chitosanases se chevauchent. Ceci n'a rien d'etonnant puisque le chitosane est souvent defini comme un derive plus ou moins deacetyle de la chitine. Bien que les endo-chitosanases puissent degrader en petits oligomeres le chitosane, elles sont incapables d'en achever la degradation complete. L'obtention de monomeres de GlcN et de GlcNAc est assuree par deux autres categories de glycosides hydrolases, les glucosaminidases (GlcNases) et les A-acetylglucosaminidases (GlcNAcases). Contrairement aux chitosanases et aux chitinases, ces enzymes sont dites
« exo », car elles enlevent successivement des residus GlcN ou GlcNAc a partir de l'extremite non reductrice de la chaine d'oligosaccharides. Sans entrer dans plus de details, il appert important de rappeler que la degradation du chitosane est un processus complexe impliquant non seulement Taction des chitosanases, mais aussi d'autres enzymes hydrolytiques faisant partie du grand groupe des glycosides hydrolases.
Enfin, il est a noter que le chitosane n ' a pas necessairement besoin d'etre degrade en monomeres pour qu'il soit efficacement utilise comme source de carbone et d'azote par les microorganismes. II a d'ailleurs ete recemment constate que la plupart des actinomycetes pourvus de chitosanases ne possedent pas de glucosaminidases. II semble que ces organismes soient en mesure de transporter et d'absorber les oligosaccharides de chitosane directement sous les formes de dimeres ou de trimeres (Cote et al., 2006). Bien que les metabolismes du chitosane et de la chitine soient etroitement lies et que plusieurs des enzymes mentionnees ci-haut travaillent en synergie lors de la degradation du chitosane dans un contexte in vivo, la suite de ce document se concentrera essentiellement sur les chitosanases.
L E S C H I T O S A N A S E S
Tel que decrit precedemment, les chitosanases sont generalement definies comme des enzymes capables de reconnaitre et de cliver le chitosane sans toutefois etre capable de cliver la chitine. Les chitosanases sont . produites par une grande variete de microorganismes. Depuis plus de trente ans, 1'etude des chitosanases a permis de conclure que la plupart des chitosanases bacteriennes sont inductibles par la presence du chitosane (Monaghan et al., 1973). De plus, leur implication dans la degradation et 1'utilisation du chitosane exogene est maintenant bien documentee (Shimosaka et al., 2005). Contrairement aux chitosanases bacteriennes, la plupart des chitosanases fongiques sont produites en absence de chitosane. Parallelement, les etudes portees sur les chitosanases d'origine fongique montrent non seulement que la plupart de ces dernieres ne sont pas en mesure d'utiliser efficacement le chitosane exogene comme source de carbone, mais aussi que l'ajout de chitosanase exogene dans le milieu de
cultures inhibe la croissance de certains champignons (Shimosaka et al., 1993; Shimosaka et al,, 2005). Bien que le role des chitosanases fongiques demeure obscur, il semble que ces dernieres participent au metabolisme de la paroi fongique (Shimosaka et al., 2005).
Les chitosanases appartiennent a plusieurs families du groupe des glycosides hydrolases, soit les families 5, 7, 8, 46, 75 et 80. Certaines de ces families comprennent des membres ayant des activites catalytiques differentes de l'activite chitosanase. C'est le cas des families GH 5, 7 et 8. En effet, la famille 5 comprend a la fois des mannosidases, des endoglycosylceraminidases, des cellulases, des licheninases, des glucosidases et des xylanases alors que la famille 7 comprend, mis a part des chitosanases, des cellobiohydrolases et des endoglucanases. En ce qui a trait a la famille 8, elle comprend des chitosanases, des cellulases, des licheninases et des endo-P-1,4-xylanases. En revanche, les families 46, 75 et 80 ne comprennent, jusqu'a ce jour, que des chitosanases.
Suite a un alignement de sequences en acides amines provenant des six differentes families de chitosanases, un arbre phylogenique a ete cree (figure 8). Ce dernier indique clairement la singularity de la sequence en acides amines de chaque famille, c'est-a-dire que ces sequences ne possedent que peu de similarity entre elles. En effet, tel qu'illustre par la longueur des branches de l'arbre, les distances phylogeniques separant les families les unes des autres sont considerables. Aussi, il est possible de voir qu'il existe une diversite considerable au sein des membres d'une meme famille. De ce fait, il est possible de retrouver des membres d'une meme famille ayant un contenu faiblement similaire (30 % - 40 %) en terme de sequences primaires.
La diversite des chitosanases se reflete egalement par la variete de leurs structures tridimensionnelles. Ces structures sont d'ailleurs differentes chez les quatre families de chitosanases pour lesquelles la structure tridimensionnelle a ete resolue (tableau 2). D'un point de vue mecanistique, les chitosanases ne different pas des autres glycosides hydrolases et ainsi fonctionnent selon un des deux types principaux de mecanisme d'hydrolyse.
0 . 1 _
Figure 8. Arbre phylogenique des families de glycosides hydrolases comprenant des chitosanases. Sources : Bac ce HI, Bacillus cereus H-I; Ba WL12, Bacillus circulans WL-12 ; Fus sola, Fusarium solani ; Asp fumi, Aspergillus Jymigatus; Asp oryz, Aspergillus oryzae IAM2660; Str coel, Streptomyces coelicolor A3(2); Bac amyl, Bacillus amyloliquefaciens\ Pse sAOl, Pseudomonas sp. A-01; Str N174, Streptomyces sp.N174; Sph mult, Sphingobacterium multivorum\ Mat chit, Matsuebacter chitosanotabidus 3001; SAV 1850, Streptomyces avermitilis MA-4680; SAV 1288, Streptomyces avermitilis MA-4680; Bac lich, Bacillus licheniformis MD1; Str gris, Streptomyces griseus HUT6037; Asp acul, Aspergillus aculeatus F-50; Hyp jeco, Hypocrea jecorina L27; Pae fuku, Paenibacillus fukuinensis D2; Bac circ, Bacillus circulans MH-K.1; ChV CVK2, Chiorella virus CVK2; Noc N106, Nocardioides sp. N106; Amy Cs02, Amycolatopsis sp. CsO-2; Bac subt, Bacillus subtilis; Bac amyl, Bacillus amyloliquefaciens\ Bac DAU, Bacillus DAU01; Pro, $-Proteobacterium KNU3. Arbre cree a partir d'un alignement des sequences en acides amines Clustal W. Visualisation Treeview 1.6.6.
Tableau 2. Proprietes generates des families de glycosides hydrolases comprenant les chitosanases
Famille Mecanisme GH d'hydrolyse Acides amines catalytiques Type de structure 3D Exemple de structure 3D
Retention Glu/Glu (P/a)s
Cellulase de
Acidothermus cellulolyticus
7 Retention Glu/Glu p -jelly roll
Cellobiohydrolase I / chitosanase de Hypocrea jecorina L27
8 Inversion Asp / Glu (a I a)6
46 Inversion Asp / Glu a+ P
Chitosanase de Bacillus sp. K17
Chitosanase de Streptomyces sp. N174
Tableau 2 (suite). Propriety generates des families de glycosides hydrolases comprenant les chitosanases
Famille GH Mecanisme d'hydrolyse Acides amines catalytiques Type de structure 3D Exemple de structure 3D
75 Inversion Asp / Glu N. D. N. D.
80 Inversion Glu/Glu N. D. N. D.
Les chitosanases des families GH 5 et 7 operent via un mecanisme a retention anomerique alors que celles des families GH 8, 46, 75 et 80 operent via un mecanisme a inversion anomerique (tableau 2). Dans tous les cas etudies a ce jour, la catalyse est assuree par une paire d'acides amines carboxyliques (tableau 2).
Mise a part la repartition des chitosanases en fonction de leur mecanisme d'action et de la similarity de leur contenu en acides amines, la specificite de clivage s'avere une autre caracteristique importante a considered C'est pourquoi, il est possible de classer les chitosanases parmi trois types ou classes (tableau 3) en fonction du genre des osamines situees de part et d'autre du lien a cliver (Fukamizo et al., 1995). II est a noter que cette specificite de clivage varie a l'interieur des chitosanases de meme famille.
Tableau 3. Specifieite de elivage du chitosane chez les chitosanases Type ou
classe Lien hydrolyse
I I
I
Endo II
III
I
J
Legende : GlcN ; o, GlcNAc La fleche ( | ) indique le site de elivage par les divers types de chitosanases. Le residu situe a la droite de cette fleche deviendra, a la suite de l'hydrolyse, l'extremite non reductrice du produit reactionnel. Les donnees comprises dans ce tableau sont tirees de Fukamizo et al., 1995.
Chitosanases de la famille 5 des glycosides hydrolases
La famille 5 des GH renferme un grand nombre d'enzymes (plus de 2000 sequences recensees) detenant une panoplie d'activites enzymatiques differentes. Jusqu'a ce jour, seulement deux chitosanases (Choi et ChoII) dont les activites ont ete confirmees par des etudes biochimiques ont ete repertories en tant que membres de cette famille. Ces chitosanases sont des proteines extracellulaires secretees par l'actinomycete Streptomyces griseus HUT 6037. En 2003, les travaux de Tanabe et al. (2003) sur ChoII, ont demontre que cette enzyme opere via un mecanisme a retention et qu'elle possede une activite transglycosylase, activite peu repandue chez les chitosanases (Tanabe et al., 2003). Ainsi done, ChoII est en mesure de transferer un (GlcN)2 ou un (GlcN)3 a une (GlcNAc)3. II en resulte alors la formation de (GlcN)2-(GlcNAc)3 et de (GlcN)3- (GlcNAc)3. Les memes
observations ont aussi ete faites concernant Choi, une autre chitosanase produite par S. griseus HUT 6037. Les etudes de cette meme equipe sur la specificite de elivage de ChoII relatent plusieurs faits interessants. Effectivement, cette enzyme hydrolyse non seulement le chitosane partiellement acetyle, mais aussi la carboxymethyl-cellulose (CMC), tout comme certaines enzymes de la famille GH 8. II faut toutefois mentionner que l'activite relative de ChoII sur le CMC n'est que 29 % par rapport a son activite sur le chitosane partiellement desacetyle. (Tanabe et al., 2003). Pour ce qui est du elivage du
chitosane, preferablement de haut degre d'acetylation, il s'effectue entre les liens GlcN-GlcN et les liens GlcN-GlcN-GlcN-GlcNAc. Done, en ce qui concerne la specificite de clivage, ChoII appartient a la classe III des chitosanases (Tanabe et al., 2003).
A ce jour, il est demontre que quelques autres enzymes appartenant a la famille GH 5 sont en mesure de degrader a la fois le chitosane et la cellulose. Ces enzymes dites bifonctionnelles sont retrouvees chez Bacillus licheniformis NBL420 et chez Myxobacter sp. AL-1 (Hong et al., 2003; Pedraza-Reyes et Gutierrez-Corona, 1997). Contrairement a ChoII, ces deux endocellulases-chitosanases degradent aussi efficacement tant la cellulose que le chitosane. Dans le cas de l'endocellulase-chitosanase de Myxobacter sp. AL-1, il est clairement etabli que la temperature reactionnelle constitue le principal facteur determinant laquelle des deux activites hydrolytiques sera privilegiee par l'enzyme. Ainsi, l'activite chitosanase sera utilisee preferentiellement a de hautes temperatures, en l'occurrence a des temperatures avoisinant les 70 °C. Inversement, l'enzyme fonctionnera en mode cellulase a des temperatures plus basses, soit des temperatures autour de 40 °C. Un comportement similaire est egalement observe chez l'endocellulase-chitosanase Bacillus licheniformis NBL420 (Hong et al., 2003).
Chitosanases de la famille 7 des glycosides hydrolases
A l'instar de certaines enzymes de la famille GH 5, quelques hydrolases de la famille GH 7 possedent une activite chitosanase et aussi une activite cellulase. En effet, il a ete tout recemment constate qu'une enzyme produite par le champignon filamenteux Trichoderma reesei (l'anamorphe de Hypocrea jecorina) possede une activite chitosanase et une activite cellobiohydrolase (Ike et al., 2007). L'equipe de Ike et al. (2007) a egalement demontre qu'il en etait de meme pour une autre hydrolase fongique secretee par Aspergillus aculeatus. Une analyse des produits reactionnels suggere que ces deux enzymes, nommees respectivement TrCBHI et AaCBHI, agissent sur le chitosane soluble a la maniere d'endo-enzymes. Paradoxalement, ces dernieres clivent la cellulose de fa<?on processive a partir de son extremite reductrice pour generer du cellobiose, ce qui fait de ces cellulases des enzymes dites « exo ». II importe d'ailleurs de rappeler ici que la
topologie de type tunnel du site catalytique de ces enzymes est caracteristique des exo-hydrolases operant de maniere processive (figure 3). La cellulase nommee CCBE, produite par l'ascomycete Trichoderma viride, est une autre chitobiohydrolase-chitosanase bifonctionnelle de la famille GH 7 (Liu et Xia, 2006). L'analyse des produits issus de la degradation du chitosane semble indiquer que CCBE procede, dans un premier temps, en mode endohydrolytique, provoquant ainsi la liberation de chitobiose, de chitotriose, de chitotetraose et de quelques chitooligosaccharides de plus grande taille, pour ensuite operer en mode exohydrolytique liberant alors des monomeres de glucosamines et des chitooligosaccharides de petite taille. Enfin, il est interessant de mentionner que la dualite endo/exohydrolytique de cette cellulase-chitosanase est deja mise a profit en milieu industriel pour la production de chitooligosaccharides (Liu et Xia, 2006).
Chitosanases de la famille 8 des glycosides hydrolases
La majorite des cellulases-chitosanases bifonctionnelles bacteriennes font partie de la famille GH 8. A ce jour, une activite chitosanase a ete mise en evidence chez plus d'une quinzaine d'enzymes de cette famille. Parmi ces enzymes, la cellulase-chitosanase de Paenibacillus fukuinensis D2 est une des plus extensivement etudiees. En ce sens, plusieurs acides amines impliques, de pres ou de loin, dans la catalyse ont deja ete identifies (Xia et al., 2008) et les proprietes physicochimiques sont bien detaillees. La caracteristique la plus distinctive de cette enzyme reside en son poids moleculaire anormalement eleve. En fait, la cellulase-chitosanase de Paenibacillus fukunensis D2 a un poids moleculaire de 85.6 kDa, soit pratiquement le double du poids moleculaire habituellement observe pour les chitosanases. Ceci s'explique par la presence de deux domaines discoidines, domaines pouvant lier le collagene et respectivement de 130 et de 131 residus. De plus, les etudes de troncation de certaines sections de cette hydrolase montrent que seule une region de 42 kDa est necessaire au maintien de 1'activite catalytique de cette enzyme (Kimoto et al., 2002). Depuis la fin des annees 1990, un nombre important de cellulases-chitosanases bifonctionnelles provenant du genre Bacillus a ete identifie en tant que membres de la famille GH 8. En voici quelques-unes a
titre d'exemple : cellulases-chitosanases provenant de Bacillus circulans WL-12, de Bacillus cereus H - l , de Bacillus cereus D - l l et de Bacillus sp. S65 (Mistsutomi et al., 1998 ; Su et al., 2006 ; Gao et al., 2008 ; Xia et al., 2008). Outre les cellulases-chitosanases du genre Bacillus, la cellulase nommee Cel8A de Lysobacter sp. IB-9374 est aussi en mesure d'hydrolyser le chitosane (Ogura et al., 2006). II est a noter que toutes les cellulases-chitosanases de la GH 8 etudiees a ce jour clivent le chitosane dans un mode endo-hydrolytique. En ce qui a trait a leur specificite de clivage, il semble, selon les donnees disponibles, que ces dernieres soient en mesure de reconnaitre le lien dit « homo » entre deux residus GlcN ainsi qu'un des deux liens dits « hetero » unissant un residu GlcN et un residu GlcNAc. Partant de ce fait, ces enzymes bifonctionnelles font partie des chitosanases de type I ou de type III (Xia et al., 2008).
Outre les enzymes deja citees, la famille GH 8 compte aussi des chitosanases ayant une specificite de clivage de type II. La chitosanase de Bacillus sp. No.7-M. et celle de Bacillus sp. K17 en sont des exemples (Adachi et al., 2004). C'est done dire que ces chitosanases sont plus strictes au niveau de la reconnaissance du substrat: seul le lien unissant des residus GlcN peut etre reconnu, puis hydrolyse. La resolution de la structure tridimensionnelle et les etudes sur les residus catalytiques de la chitosanase ChoK de Bacillus sp. K17 ont permis de faire ressortir quelques points interessants concernant les enzymes de la famille GH 8 (Adachi et al., 2004). Une comparaison entre la structure tridimensionnelle de ChoK et celle de CelA (la cellulase de Clostridium thermocellum NCIB 10682/JW20 de la famille GH 8) suggere que la position du residu glutamate dit donneur de protons est conservee (Glu 122 chez ChoK vs Glu95 chez CelA), mais que le residu aspartate correspondant a l'accepteur de protons chez CelA (Asp278) est inactive chez ChoK (via une substitution en asparagine). En comparant plus en detail la structure secondaire de ChoK a celle de CelA, Adachi et al. (2004) ont demontre la presence d'une boucle s u p p l e m e n t a l , composee d'un feuillet p, au coeur du site catalytique de ChoK. Toujours selon Adachi et al. (2004), dans le cas de ChoK, le residu dit accepteur de protons se situe a meme cette boucle. De plus, les etudes de mutagenese dirigee realisees par cette meme equipe attribuent la fonction d'accepteur de protons a un residu glutamate