Étude de l’évolution et de la diversité des poissons
d’eau douce de l’Amérique du Nord par une approche
génétique comparative
Thèse
Julien April
Doctorat en biologie
Philosophiae Doctor (PhD)
Québec, Canada
© Julien April, 2013
Résumé
La variation génétique des poissons d‟eau douce de l‟Amérique du Nord a été analysée aux niveaux intraspécifique et interspécifique afin d‟approfondir les connaissances actuelles sur l‟évolution de la biodiversité et de faciliter la conservation de cette richesse. Premièrement, une banque de données de séquence d‟ADN mitochondrial (codes-barres génétiques) a été générée pour 5674 individus représentant 752 espèces dans le but d‟établir un outil d‟identification moléculaire et de fournir une calibration de l‟incertitude taxonomique. Cette analyse démontre que 90 % des espèces peuvent être délimitées à l‟aide de codes-barres génétiques. De plus, il apparait que la taxonomie actuelle pourrait largement sous-estimer la diversité des poissons d‟eau douce dans son ensemble tout en surestimant la diversité spécifique de certains groupes particuliers. Deuxièmement, les niveaux de divergences génétiques intraspécifique et interspécifique ont été étudiés afin d‟identifier les mécanismes évolutifs responsables des patrons généraux de répartition de la biodiversité. L‟hypothèse suggérant un rôle combiné des cycles glaciaires du Pléistocène et de l‟activité métabolique influençant les taux de mutation est appuyée par les données représentant la presque totalité des poissons d‟eau douce de l‟Amérique du Nord. Troisièmement, les patrons de variation aux niveaux de l‟ADN mitochondrial et de l‟ADN nucléaire (AFLP) ont été analysés parmi plusieurs paires de lignées évolutives codistribuées dans le nord-est de l‟Amérique afin de vérifier l‟importance du processus de spéciation allopatrique. Les résultats indiquent que l‟est du bassin des Grands Lacs représente une zone de suture impliquant plusieurs taxons et que les niveaux de divergence sont variables d‟une paire de lignées à l‟autre. Parmi ces paires de races glaciaires, celles divergeant par plus de 2 % au niveau de l‟ADN mitochondrial présentent d‟importants signes d‟isolement reproducteur. En décrivant les diverses étapes du processus de spéciation allopatrique au sein de différentes espèces d‟une même région, cette étude démontre que le processus de spéciation allopatrique a joué un rôle important parmi les poissons d‟eau douce de l‟Amérique du Nord. L‟ensemble de ces travaux aura donc permis de fournir un nouvel outil d‟identification et d‟appuyer l‟hypothèse stipulant que la spéciation allopatrique est le principal moteur de diversification pour les poissons d‟eau douce.
Abstract
Intraspecific and interspecific genetic variation has been studied among North America‟s freshwater fishes in order to improve our current knowledge on the evolution of biodiversity and to facilitate the conservation of this richness. Firstly, we generated a standard reference library of mitochondrial DNA sequences (DNA barcodes) for 752 North American freshwater fish species to provide an independent calibration of taxonomic uncertainty and to establish a more accessible molecular identification key for its application. This study demonstrates that 90% of known species can be delineated using barcodes. Results further suggest that current North American freshwater fish taxonomy at the species level significantly conceals diversity in some groups, while artificially creating diversity in others. Secondly, we studied intraspecific and interspecific genetic divergence in order to describe and identify the underlying evolutionary causes of general patterns of biodiversity distribution. This study supports a dual role involving both the late Pliocene-Pleistocene climatic fluctuations and metabolic rate in determining latitudinal gradients of genetic divergence. Thirdly, patterns of mitochondrial DNA and nuclear DNA (AFLP) have been studied among different codistributed pairs of glacial lineages in order to verify the generality of allopatric speciation. This study shows that the Eastern Great-Lakes drainage represents a multi-species suture zone for glacial lineages of freshwater fishes with variable levels of genetic divergence. AFLP analyses among four pairs of lineages indicate that lineages with relatively deep levels of mitochondrial DNA sequence divergence (>2 %) developed strong reproductive barriers. By describing different levels of divergence and reproductive isolation in different co-occurring fishes, we offer strong evidence that allopatric speciation has contributed significantly to the diversification of North Eastern American freshwater fishes. This thesis therefore offers a new molecular identification tool for freshwater fish of North America and brings strong evidences that allopatric speciation has played a predominant role in generating biodiversity.
Avant propos
Remerciements
Je tiens premièrement à remercier mon directeur de recherche Louis Bernatchez. Mon aventure dans le laboratoire de Louis a débuté par une histoire de pêche. En effet, c‟est après avoir mentionné à Louis que je venais tout juste de faire une capture inusitée dans la rivière Cap-Rouge, un saumon atlantique juvénile (photo à l‟appui), qu‟il a proposé de m‟engager comme professionnel de recherche afin de développer une collection de poissons d‟eau douce. Ce n‟était que le premier des nombreux projets passionnants qu‟il allait me proposer. Durant mon projet de doctorat, Louis a non seulement été un très précieux conseiller, mais aussi un directeur dont l‟enthousiasme sans limite a rendu le séjour dans son laboratoire extrêmement agréable. Bref, pour avoir cru en moi et pour m‟avoir permis de dépasser mes limites, le tout dans un contexte des plus plaisants, je te remercie sincèrement Louis.
Mon projet de doctorat n‟aurait certainement pas eu la même ampleur sans la collaboration de mon codirecteur Robert Hanner (University of Guelph), lequel m‟a introduit dans l‟équipe du International Barcode of Life Project (iBOL) et m‟a conseillé de façon judicieuse. Paul Hebert (Guelph University), fondateur de ce groupe de recherche, a aussi été d‟un soutien de taille à mon projet. Je me dois de remercier spécialement Richard Mayden (St-Louis University) pour avoir accepté de partager l‟accès à sa collection de poissons, laquelle comprend peut-être le plus grand nombre de poissons d‟eau douce américains dont les tissus ont été conservés de façon à permettre les analyses génétiques. En plus des échantillons amassés durant sa prospère carrière, l‟expertise taxonomique de Richard a aussi été mis à contribution durant les analyses associées au projet du deuxième chapitre.
Je tiens aussi à remercier tous les gens avec qui j‟ai eu le plaisir d‟avoir des discussions scientifiques des plus enrichissantes. Je ne peux passer sous silence le fait que Julie Turgeon, ma directrice de maitrise, m‟a permis d‟acquérir une curiosité et un bagage de connaissances qui m‟ont largement aidé durant mes études doctorales. Julian Dodson et Nicolas Derome font aussi partie des professeurs du département de biologie qui m‟ont fourni de très précieux conseils. Parmi les étudiants et professionnels avec qui j‟ai travaillé, quelques-uns se sont démarqués par la profondeur et la justesse de leurs propos. Je pense en particulier à Vincent Bourret, Guillaume Côté, Pierre-Alexandre Gagnaire, Simon Blanchet, David Paez, Charles Perrier, Arne Nolte et Scott McCairns.
Finalement, je désire remercier affectueusement ma conjointe Cécilia Hernandez, ainsi que ma mère France Simard, qui m‟ont supporté et encouragé durant la rédaction de la thèse. L‟arrivée de mon fils Paul, le 24 mai 2012, a été une belle source d‟inspiration et de motivation.
Organisation de la thèse
Cette thèse est organisée en cinq chapitres. Le premier chapitre consiste en une introduction générale en français. Les chapitres deux, trois et quatre ont tous fait l'objet d'un article de recherche publié ou en voie de l'être. Ils ont été rédigés en anglais sous un format approprié pour la publication scientifique. Le chapitre cinq consiste en une conclusion générale en français. Le matériel supplémentaire mentionné dans les chapitres deux à quatre est inclus en annexe.
Le chapitre 2 est publié sous la référence: Julien April, Richard L. Mayden, Robert Hanner and Louis Bernatchez (2011). Genetic Calibration of Species Diversity among North America's Freshwater Fishes. Proceedings of the National Academy of Sciences of the United States of America. 108, 10602–10607. J.A., R.L.M., R.H. et L.B. ont conçu le projet; J.A., R.L.M., R.H. et L.B. ont réalisé le projet; J.A. et R.L.M. ont analysé les données et J.A. a écrit l‟article.
Le chapitre 3 est soumis sous la référence: Julien April, Richard L. Mayden, Robert Hanner and Louis Bernatchez. Metabolic rate and climatic fluctuations shape continental wide pattern of genetic divergence and biodiversity in fishes. J.A., R.L.M., R.H. et L.B. ont conçu le projet; J.A., R.L.M., R.H. et L.B. ont réalisé le projet; J.A. et R.L.M. ont analysé les données et J.A. a écrit l‟article. Au moment du dépôt initial de cette thèse, cet article était accepté dans la revue PLOS ONE sous condition d‟effectuer des révisions mineures. La version révisée est présentée dans ce document.
Le chapitre 4 est publié sous la référence: Julien April, Robert Hanner, Anne-Marie Dion‐Côté & Louis Bernatchez (2013). Glacial cycles as an allopatric speciation pump in north‐eastern American freshwater fishes. Molecular ecology, 22(2), 409-422. J.A. et L.B. ont conçu le projet; J.A., R.H. et L.B. ont réalisé le projet; J.A. et A.-M.D.-C. ont effectué le travail de laboratoire; J.A. a analysé les données et écrit l‟article; L.B. et R.H. ont révisé et commenté l‟article.
Table des matières
Résumé ... iii
Abstract ... v
Avant propos ... vii
Table des matières ... ix
Liste des tableaux ... xi
Liste des figures ... xiii
Lexique des abréviations utilisées ... xv
Chapitre 1: Introduction générale ... 1
1.1 Biodiversité ... 1
1.2 Évolution de la biodiversité ... 4
1.3 Évolution et diversité des poissons d‟eau douce de l‟Amérique du Nord ... 5
1.4 Lacunes et défis dans l‟étude de la biodiversité et des poissons d‟eau douce ... 8
1.4.1 Identification des espèces ... 8
1.4.2 Patrons de répartition spatiale de la biodiversité ... 10
1.4.3 Spéciation allopatrique et pompe à spéciation ... 12
1.5 Objectifs ... 15
1.6 Marqueurs génétiques ... 16
Chapitre 2: Genetic Calibration of Species Diversity among North America's Freshwater Fishes ... 19
2.1 Résumé ... 19
2.2 Abstract ... 19
2.3 Introduction ... 20
2.4 Materials and methods ... 21
2.5 Results ... 22
2.6 Discussion ... 23
2.7 Conclusion ... 29
2.8 Acknowledgments ... 34
2.9 Data accessibility ... 34
Chapitre 3: Metabolic rate and climatic fluctuations shape continental wide pattern of genetic divergence and biodiversity in fishes ... 35
3.1 Résumé ... 35
3.2 Abstract ... 35
3.3 Introduction ... 36
3.4 Materials and Methods ... 38
3.5 Results ... 41
3.6 Discussion ... 43
3.7 Acknowledgments ... 51
3.8 Data accessibility ... 51
Chapitre 4 : Glacial cycles as an allopatric speciation pump in North Eastern American freshwater fishes ... 53
4.1 Résumé ... 53
4.2 Abstract ... 53
4.3 Introduction ... 54
4.5 Results ... 60
4.6 Discussion ... 63
4.7 Acknowledgments ... 73
4.8 Data accessibility ... 73
Chapitre 5: Conclusion générale ... 75
5.1 Conclusion ... 75
5.2 Implications pratiques et fondamentales ... 78
5.3 Réflexion sur les concepts d‟espèce ... 80
5.4 Perspectives... 82
Bibliographie ... 83
Liste des tableaux
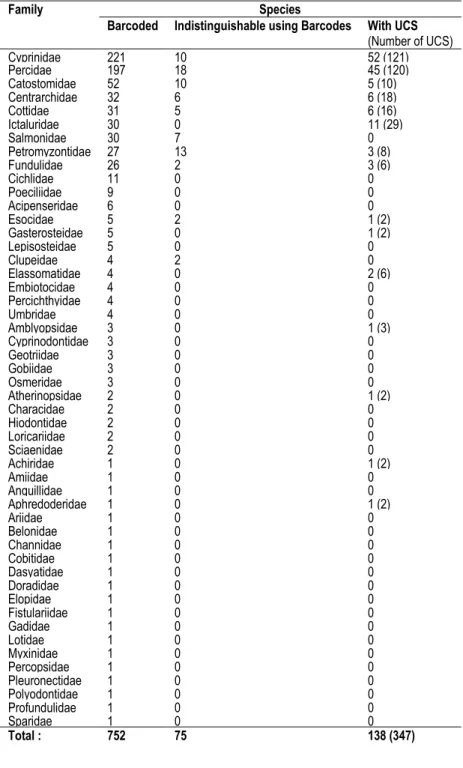
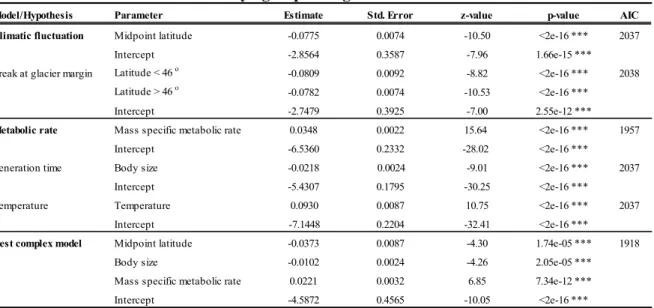
Table 2.1 Summary of the fish taxa analyzed, including the number of indistinguishable species and the number of species with UCS (unconfirmed candidate species; represented by lineages that diverge by over 2 %), along with the total number of UCS (species list in Supporting Text). ... 30 Table 3.1 Results of generalized linear mixed models.. ... 48 Table 4.1 Basic data and assignment results based on AFLP for the secondary contact analyses ... 68 Table 4.2 Divergence between Mississippian and Atlantic glacial lineages from different freshwater fish taxa. 69
Liste des figures
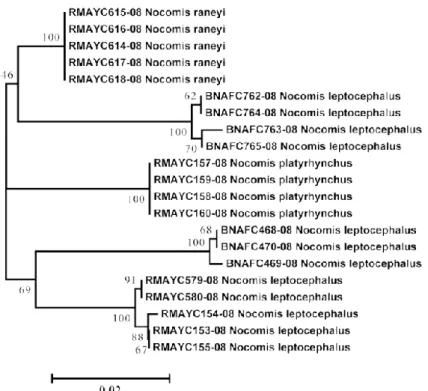
Figure 2.1 Neighbor joining tree that shows the relationships among 615 pb COI sequences from 3 species of the genus Nocomis, producing 5 divergent clusters.. ... 31
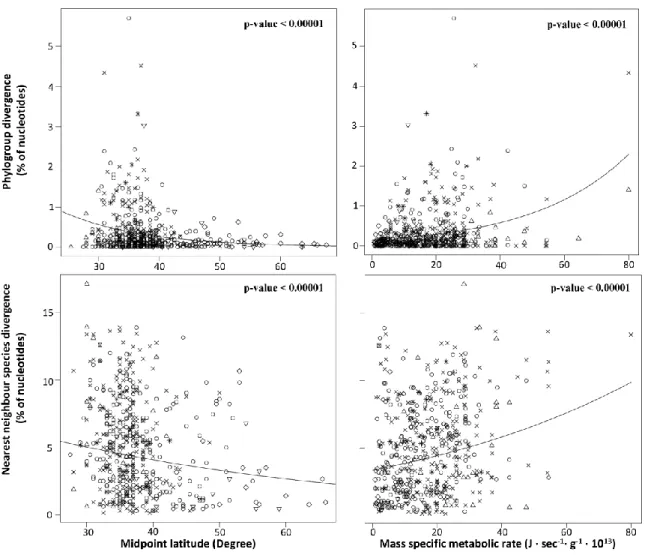
Figure 2.2 Neighbor-joining tree showing the relationships among 620 bp COI sequences from 6 parasitic and 7 non-parasitic species of lampreys, producing 5 clusters ... 33 Figure 3.1 Plots of genetic divergence against midpoint latitude and mass specific metabolic rate using raw data. ... 49
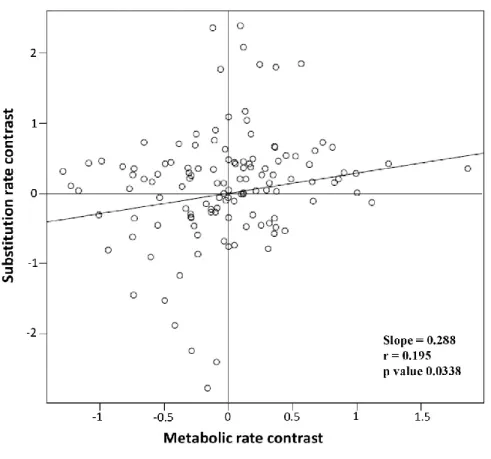
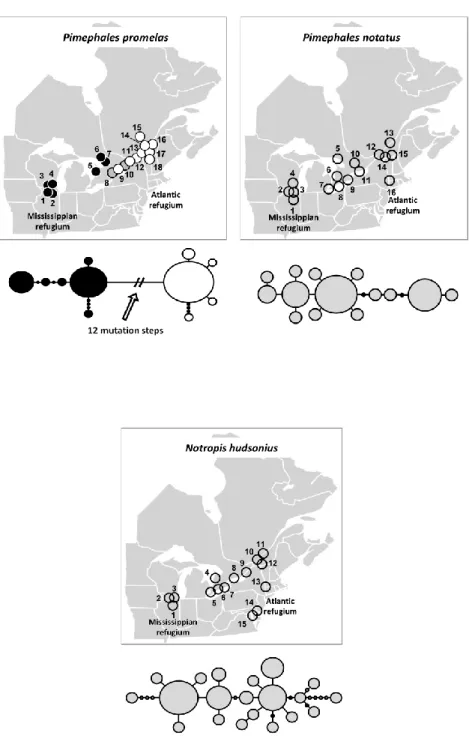
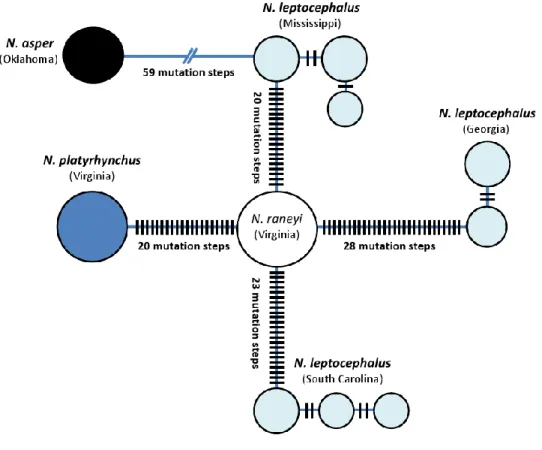
Figure 3.2 Plots of phylogenetically independent sister pair comparisons of mutation rate against mass specific metabolic rate ... 50 Figure 4.1 Map of samples localities (above) and mtDNA haplotype network (below) for each taxon ... 71
Figure 4.2 Bar plot showing the admixture proportion of each specimen based of AFLP data and calculated using the program STRUCTURE ... 72
Lexique des abréviations utilisées
ADN (DNA) :
Acide désoxyribonucléique (deoxyribose nucleic acid)
ADNmt (mtDNA) : ADN mitochondrial (mitochondrial DNA)
ADNnuc (nucDNA) : ADN nucléaire (nuclear DNA)
AFLP :
Polymorphisme de longueur de fragments amplifiés (amplified
fragment length polymorphism)
UES (ESU) :
Unité évolutive significative (evolutionarily significant unit)
PCR :
Réaction de polymerase en chaine (polymerase chain reaction)
p-value :
Valeur de probabilité associée à un test statistique (probability value
Chapitre 1: Introduction générale
1.1 Biodiversité
La diversité du vivant représente une ressource essentielle à la survie de l‟humanité (Ehrlich et Ehrlich 1992, Cardinale et al. 2012) et suscite la fascination depuis des centaines d‟années (ex. : Aristote - Singer 1931). Ce n‟est toutefois qu‟en 1988 qu‟Edward Osborne Wilson introduit le terme « biodiversité » dans la littérature. Selon l‟article 2 de la Convention sur la diversité biologique (conférence de Rio 1992; Antoine et al. 1994), la biodiversité a pour définition la « variabilité des organismes vivants de toute origine y compris, entre autres, les écosystèmes terrestres, marins et autres écosystèmes aquatiques et les complexes écologiques dont ils font partie : cela comprend la diversité au sein des espèces et entre espèces ainsi que celles des écosystèmes ». Ce concept de biodiversité reflète bien la constatation de plus en plus évidente que la diversité du vivant ne peut simplement se résumer qu‟à un nombre d‟espèces (Lomolino et al. 2006). Il existe non seulement plusieurs niveaux d‟organisations biologiques, mais de plus, ces niveaux sont en constante évolution et interagissent les uns avec les autres.
La diversité intraspécifique se définit par la variabilité observée à l‟intérieur des espèces et inclut, par exemple, la diversité génétique et phénotypique au sein d‟une espèce (Union internationale pour la conservation de la nature-UICN 2013). Il existe au moins trois raisons pour conserver la diversité génétique intraspécifique. Premièrement, la perte de diversité génétique réduit la capacité d‟adaptation des organismes face aux changements environnementaux puisque la sélection naturelle agit à partir de la variation déjà existante (Frankham 1995, Avise et Hamrick 1996). Deuxièmement, le maintien de cette partie intégrante de la biodiversité actuelle (UICN 2013) est d‟autant plus important puisque le gain de diversité génétique, ainsi que la formation de lignées évolutives intraspécifiques très divergentes, sont des processus lents puisqu‟ils dépendent des taux de mutation. Troisièmement, plusieurs modèles théoriques et études empiriques démontrent qu‟il existe une relation entre le fitness et l‟hétérozygotie, en partie en raison de l‟expression de la dépression de consanguinité attendue chez les organismes ayant une diversité génétique limitée (Reed et Frankham 2003). Il est donc évident que la diversité intraspécifique doit être conservée afin de permettre la pérennité des processus évolutifs et de diversification.
Plusieurs concepts d‟unité de gestion intraspécifique ont été proposés pour des fins de conservation. Les sous-espèces représentent probablement l‟unité intraspécifique la plus largement reconnue (Nelson et al. 1974, Lomolino et al. 2006, Wilson et Brown 1953). Toutefois, le manque de précision et d‟objectivité dans la désignation des sous-espèces limite grandement l‟utilité de cette unité en conservation (Phillimore et Owens
2006). Ainsi, bien que les sous-espèces soient souvent définies sous la base d‟une variation géographique de caractères phénotypiques (Mayr et Ashlock 1991), le manque de précision de cette définition et l‟emploi de différentes définitions compliquent l‟utilisation de ce concept. De plus, des études démontrent que les sous-espèces définies selon des bases morphologiques ne correspondent pas toujours à des groupes génétiques distincts (revue dans Phillimore et Owens 2006). Le terme « unité évolutive significative » (UES) (en anglais: « evolutionarily significant unit » ou « ESU ») a été introduit par Ryder en 1986 et désigne une sous-unité du groupe plus inclusif que représentent les espèces qui possèdent des attributs significatifs pour le présent et le futur des générations de l‟espèce en question. Depuis Ryder (1986), plusieurs autres définitions ont été proposées (revue dans Fraser et Bernatchez 2001). Toutefois, la plupart de ces définitions semblent converger vers un point; les UES devraient correspondre à des lignées évolutives qui présentent un certain degré d‟isolement reproducteur et qui possèdent une histoire évolutive distincte (Waples 1991, Fraser et Bernatchez 2001). Mentionnons que les analyses génétiques se sont particulièrement révélées utiles afin d‟identifier ces unités (Frankham et al. 2002).
Le niveau hiérarchique de la biodiversité correspondant à l‟espèce est souvent perçu comme occupant une position centrale dans l‟étude de la biodiversité. Ainsi, les processus évolutifs ou écologiques observés à l‟intérieur des espèces sont définis comme microévolutifs et microécologiques alors que les processus observés entre les espèces sont définis comme macroévolutifs et macroécologiques (Reznick et Ricklefs 2009). Depuis le début de la taxonomie moderne (Linnaeus 1735), le niveau spécifique représente l‟unité de base dans la classification taxonomique. Parmi les différentes mesures de biodiversité, la diversité en espèces est la plus utilisée (UICN, Whittaker 1972, Lomolino et al. 2006). Enfin, alors qu‟il est généralement accepté que les groupements taxonomiques inférieurs et supérieurs au niveau de l‟espèce ne soient que des constructions artificielles imaginées par l‟homme et pouvant être subdivisées ou fusionnées de façon arbitraire, il pourrait bien en être différemment des espèces (Ereshefsky 1999, Coyne et Orr 2004). En effet, il a été suggéré que le fait que la grande majorité des scientifiques et des peuples habitant des régions isolées et ayant conservé leur vision traditionnelle de la faune et de la flore s‟accordent à définir les mêmes groupes d‟individus comme des espèces appuie l‟idée que les espèces soient des entités bien réelles (Mayr 1963, Diamond 1966). Considérant les espèces comme des groupes d‟individus pouvant se reproduire les uns avec les autres et demeurant isolés des autres groupes d‟individus, il semble en effet que les espèces correspondent à des groupes discrets chez les animaux sexués. Toutefois, dépendamment des définitions d‟espèces adoptées, la réalité des espèces peut devenir plus discutable (Hendry et al. 2002).
Il existe une multitude de définitions opérationnelles pour les espèces. Parmi les concepts les plus communs, celui d‟espèces biologiques basées sur l‟isolement reproductif est probablement le plus largement connu
(Mayr 1942, Coyne et Orr 2004). Ce concept définit les espèces comme des groupes d‟individus pouvant se reproduire les uns avec les autres et qui demeurent isolés des autres groupes d‟individus. Le concept d‟espèces phénotypiques (Sokal et Crovello 1970), lequel est probablement le plus largement utilisé en taxonomie traditionnelle, décrit une espèce comme un groupe d‟individus morphologiquement distinct des autres groupes et ne présentant que peu ou pas d‟intermédiaires lorsqu‟en contact avec un groupe différent. Un troisième concept d‟espèce, le concept phylogénétique (Cracraft 1983), définit une espèce comme un clade basal d‟individus partageant un ancêtre commun et distinct des autres clades. Ce concept est largement utilisé chez les microorganismes. Un quatrième concept général, celui d‟espèces écologiques (Van Valen 1976), définit plutôt les espèces comme des lignées occupant une niche écologique différente de celles des autres organismes occupant la même région.
La persistance d‟un débat sur les définitions opérationnelles d‟espèces est en partie causée par l‟aspect le plus souvent graduel de la formation des espèces (De Queiroz 2007). Ainsi, puisque les différences morphologiques, génétiques, écologiques, ainsi que l‟isolement reproducteur, ne se développent généralement pas de façon synchronisée, les jeunes espèces peuvent parfois ne répondre qu‟aux critères de quelques-uns des nombreux concepts d‟espèces. À l‟opposé, bon nombre de « vieilles » espèces répondent aux critères de chacun des concepts d‟espèces. Le débat se situe donc assez spécifiquement autour d‟une définition opérationnelle permettant de trancher de façon nette entre les groupes intraspécifiques et les groupes interspécifiques (Wiley et Mayden 1985, De Queiroz 2007). D‟un point de vue pratique, il semble que la meilleure définition opérationnelle puisse varier en fonction du système étudié. D‟un point de vue taxonomique, une approche intégrant divers types d‟information (par exemple morphologique, génétique et écologique), comme préconisée en taxonomie intégrative (Pedial et al. 2010), pourrait constituer le meilleur choix. D‟un point de vue plus fondamental, plusieurs biologistes s‟entendent sur un concept théorique général commun définissant une espèce comme une lignée évolutive composée d‟individus descendant d‟un ancêtre commun et conservant une histoire évolutive indépendante des autres groupes d‟individus (Wiley et Mayden 1985, De Queiroz 2007). Enfin, notons que les définitions d‟espèces basées sur la présence d‟isolement reproducteur complet ont la particularité d‟identifier des espèces ayant atteint un point de non-retour dans le processus de spéciation (Muller 1939, Coyne et Orr 2004).
La diversité interspécifique se définit par la variabilité observée entre les espèces ainsi qu‟à l‟intérieur et entre les groupements hiérarchiques occupant un niveau supérieur aux espèces. Parmi les mesures de biodiversité interspécifique, la plus commune est peut-être la diversité écosystémique (Lomolino et al. 2006). Cette diversité inclut non seulement les différentes espèces d‟un assemblage, mais aussi les composantes abiotiques de l‟habitat de ces organismes. D‟un point de vue taxonomique, la diversité interspécifique réfère à
la diversité en genres, familles et ordres ainsi qu‟aux autres groupes taxonomiques de niveau plus élevé que les espèces. Le concept plutôt récent de diversité fonctionnelle (Walker 1992, Tilman et al. 1997, Tilman 2001) s‟applique aussi le plus souvent à la diversité interspécifique dans le cadre d‟études écologiques et désigne la diversité en fonction des différentes niches écologiques remplies par les organismes d‟un écosystème.
1.2 Évolution de la biodiversité
Les espèces ne sont pas immuables, mais se modifient au cours du temps et peuvent se transformer en de nouvelles espèces (Darwin et Wallace 1858). Le développement de la théorie de l‟évolution a eu un impact majeur sur la société dans les années 1800. Chez les biologistes, il est devenu évident que « rien en biologie n'a de sens, si ce n'est à la lumière de l'évolution » (Dobzhansky 1937). Il apparait donc essentiel d‟inclure des notions de biologie évolutive dans l‟étude de la biodiversité.
Au niveau intraspécifique, les forces évolutives ont largement été étudiées dans un contexte de génétique des populations. Les quatre principales forces agissant à ce niveau sont la mutation, la dérive génétique, le flux génique et la sélection naturelle (Wright 1931). Au niveau interspécifique, les forces évolutives ont beaucoup été étudiées dans un contexte biogéographique. À cette échelle, les principales forces influençant l‟évolution de la biodiversité sont la spéciation, l‟extinction et la migration (Lomolino et al. 2006). Il existe donc certains liens entre les forces évolutives agissant à ces deux niveaux d‟organisation. Ainsi, le flux génique agit au niveau intraspécifique de façon analogue à la migration au niveau interspécifique, ces deux forces permettent ainsi le déplacement de la biodiversité. De même, la mutation et la spéciation sont toutes deux des forces produisant de la nouvelle diversité. Les niveaux d‟organisation intraspécifique et interspécifique sont aussi influencés par la perte de diversité. L‟homogénéisation génétique parfois causée par la dérive génétique et la sélection peut se comparer à l‟extinction.
L‟étude de phénomènes complexes peut bénéficier d‟une approche réductionniste afin d‟identifier des mécanismes de premières importances (ex. : modèle de biogéographie insulaire de MacArthur et Wilson 1967). Par exemple, d‟un point de vue génétique, il a été suggéré que les processus de l‟évolution moléculaire puissent s‟expliquer sans faire intervenir la sélection (théorie neutraliste de l‟évolution moléculaire, sensu Kimura 1985). D‟un point de vue biogéographique, les travaux de Hubbell (2001) suggèrent aussi qu‟il est possible de modéliser les patrons de variation de la biodiversité sans impliquer le rôle des niches écologiques et les différences entre espèces (théorie neutre de la biodiversité, sensu Hubbell 2001). L‟utilisation de tels modèles ne consiste pas nécessairement à nier l‟existence des facteurs qui ne sont pas modélisés, mais
plutôt à étudier un modèle suffisamment simple pour être rigoureusement testé et suffisamment puissant pour expliquer le phénomène en question.
L‟origine des espèces représente un sujet qui suscite l‟imaginaire depuis des centaines d‟années et qui attire beaucoup d‟attention en biologie (Darwin 1859, Coyne et Orr 2004). Aujourd‟hui, les processus menant à la formation de nouvelles espèces sont couramment classifiés de façon géographique (Coyne et Orr 2004). La spéciation allopatrique implique l‟isolement géographique de deux groupes initialement semblables et interféconds. Avec le temps, les groupes isolés se différencient et développent des barrières reproductives pour finalement former deux espèces distinctes. Il s‟agit du mode de spéciation le plus généralement accepté parmi les biologistes et vraisemblablement le plus courant chez les animaux (Coyne et Orr 2004). À l‟autre extrême, la spéciation sympatrique implique que les espèces ont occupé le même territoire et n‟ont donc pas été isolées géographiquement durant le processus de spéciation. Puisque les deux groupes demeurent en contact géographique durant la divergence, la spéciation sympatrique implique souvent un très grand rôle de la sélection naturelle ou sexuelle (Coyne et Orr 2004).
Le développement d‟incompatibilités génétiques permet aux groupes génétiques de conserver une histoire évolutive indépendante et donc d‟achever le processus de spéciation; du moins selon le concept théorique général et le concept opérationnel d‟espèces biologiques. Contrairement aux barrières écologiques ou causées par la sélection sexuelle, lesquelles peuvent disparaitre lors de changements environnementaux, les incompatibilités génétiques représentent des barrières permanentes (Muller 1939, Coyne et Orr 2004, Seehausen 2006, Taylor et al. 2006, Vonlanthen et al. 2012). Les incompatibilités de type Bateson-Dobzhansky-Muller (Dobzhansky 1937, Muller 1942) représentent probablement le modèle d‟incompatibilité génétique le plus largement étudié et possiblement aussi un des types d‟incompatibilité les plus communs en nature. Ce modèle propose que deux lignées puissent se trouver isolées en raison d‟interactions épistatiques délétères entre les allèles de différents locus chez les hybrides (Dobzhansky 1937, Muller 1942).
1.3 Évolution et diversité des poissons d’eau douce de l’Amérique
du Nord
Les poissons représentent une partie importante de la biodiversité. Ils ont une très grande valeur économique et sociale. De plus, avec près de 30 000 espèces vivantes (Moyle et Cech 2004), il s‟agit du groupe de Vertébrés (total = environ 50 000 espèces) qui est de loin le plus diversifié (Hickman et al. 2006). La diversité en poissons d‟eau douce est aussi particulièrement surprenante. Ainsi, bien qu‟il y ait 10 000 fois plus d‟eau salée dans les mers et océans que d‟eau douce dans les lacs et rivières, les poissons d‟eau douce
représentent un peu plus de 40 % de la diversité en espèces de poissons (Moyle et Cech 2004). Cette diversité disproportionnée pourrait s‟expliquer par la très grande fragmentation des habitats d‟eau douce comparée aux eaux salées, ce qui pourrait favoriser la divergence et la spéciation allopatrique.
L‟étude des poissons s‟est avérée riche en découverte pour les évolutionnistes. Par exemple, les travaux de David Starr Jordan portant sur la répartition des espèces de poissons ont mis en évidence l‟importance du processus de spéciation allopatrique (Jordan 1908). Comme le stipule la loi de Jordan (anglais « Jordan‟s law »), les espèces les plus apparentées, c‟est-à-dire les espèces sœurs, ont très peu de possibilité de se retrouver dans la même région, mais ont plutôt tendance à être situées de part et d‟autre d‟une barrière à la dispersion. D‟un autre côté, les poissons ont aussi fourni quelques-uns des exemples de spéciations sympatriques les plus convaincants (Schliewen et al. 1994, Barluenga et al. 2006). Enfin, la faune ichthyologique a offert une des premières démonstrations de spéciation par hybridation chez les animaux (DeMarais et al. 1992).
La faune ichthyenne des eaux douces de l‟Amérique du Nord est la plus diversifiée des zones tempérées (Abell et al. 2000). Avec environ 900 espèces, elle contient plus d‟espèces que l‟Europe (environ 250) et l‟Australie (environ 230) réunies, mais moins que l‟Asie (environ 1500), l‟Afrique (environ 1800) et l‟Amérique du Sud (environ 2200) (Moyle et Cech 2004). L‟Amérique du Nord est occupée par certains groupes de poissons archaïques (Nelson et al. 2004), incluant les lamproies (Petromyzontidae, parfois considérées comme un groupe à part des poissons), les esturgeons (Acipenseridae), le poisson spatule (Polyodontidae), les lépisostées (Lepisosteidae) et les poissons castor (Amiidae). Parmi les groupes de poissons plus récents (Téléostéens), on compte deux familles extrêmement diversifiées, les Percidae et les Cyprinidae. Au sein de ces deux familles, on compte aussi les deux genres les plus diversifiés du continent (Etheostoma et Notropis) (Nelson et al. 2004). L‟Amérique du Nord comporte quelques familles endémiques telles que les Centrarchidae, Ictaluridae, Hiodontidae, Percopsidae, Ambyopsidae et Aphredoderidae (Hocutt et Wiley 1986). Parmi l‟ensemble des quelques 900 espèces du Canada et des États-Unis, la majorité est incluse dans seulement huit genres très diversifiés (Etheostoma et Percina (Famille Percidae), Notropis et Hybopsis (Famille Cyprinidae), Moxostoma (Famille Catostomidae), Cottus (Famille Cottidae), Ictalurus et Noturus (Famille Ictaluridae) et Lepomis (Famille Centrarchidae) (Hocutt et Wiley 1986, Nelson et al. 2004).
Quelques patrons généraux ressortent de l‟examen de la diversité et de la répartition des poissons d‟eau douce de l‟Amérique du Nord. Premièrement, il apparait que les radiations évolutives les plus importantes se sont produites chez des poissons de petites tailles (généralement moins de 20 cm à l‟âge adulte) (Hocutt et Wiley 1986). Deuxièmement, il existe un important gradient latitudinal en diversité spécifique. De fait, même si
le Canada et l‟Alaska représentent un territoire plus vaste que les États américains contigus, seulement 19 % des espèces occupent cette grande région et moins de 1 % des espèces y sont géographiquement restreintes (Hocutt et Wiley 1986). Afin de comprendre l‟origine de ce gradient en espèces, l‟étude géologique du continent apparait incontournable.
Plusieurs paramètres de la géographie de l‟Amérique du Nord ont été affectés par les cycles glaciaires du Pléistocène. Au total, c‟est près d‟une vingtaine de cycles glaciaires, d‟une durée d‟environ 100 000 ans chacun, qui ont eu lieu durant les derniers trois millions d‟années (Martinson et al. 1987). Parmi les plus importants en Amérique du Nord, on compte le Nebraskien, le Kansien, l‟Illinoien et le Wisconsinien. Les cycles glaciaires ont affecté le climat sur le continent entier et l‟avancée d‟immenses glaciers a complètement éliminé la faune aquatique dans les régions nordiques couvertes de glace (Dyke et Prest 1987, Pielou 1992, Hewitt 1996).
Le Wisconsinian est la plus récente des périodes glaciaires (Martinson et al. 1987). L‟avancée des glaciers, qui a atteint son maximum il y a environ 18 000 ans, s‟est étendue vers le sud jusqu‟à environ la limite entre le Canada et les États-Unis (Dyke et Prest 1987). Toutefois, une partie du nord du Canada, de l‟Alaska et de la Béringie est demeurée libre de glace. Il a donc été possible pour les organismes aquatiques de survivre dans ces endroits ainsi qu‟au sud des glaciers (Hocutt et Wiley 1986). La localisation de ces sites, typiquement appelés refuges glaciaires, a principalement été inférée par l‟étude de la répartition des espèces actuelles, des fossiles et des parasites, ainsi que par l‟analyse de la variation spatiale des caractères morphologiques et génétiques (Hocutt et Wiley 1986, Bernatchez et Wilson 1998). Parmi les refuges glaciaires les plus importants pour les poissons, on compte les refuges de la Béringie (région de l‟Alaska), Banff-Jasper (montagnes Rocheuses), Colombien (État de Washington), Missourien (Montana/Dakota du Nord), Mississippien (Missouri/Illinois) et Atlantique (au sud de Long Island sur la plaine Atlantique) (Hocutt et Wiley 1986).
Avec le réchauffement du climat, la recolonisation des territoires rendus libres de glace fut facilitée par une succession d‟immenses cours d‟eau (Hocutt et Wiley 1986, Dyke et Prest 1987). Les eaux provenant en partie de la fonte des glaces ont engendré des connexions entre des bassins versants aujourd‟hui isolés (Dyke et Prest 1987). Par exemple, alors que le bassin des Grands Lacs s‟écoule actuellement vers l‟Atlantique par le fleuve Saint-Laurent, une large portion des cours d‟eau occupant le territoire actuel des Grands Lacs s‟est déversée vers le sud par le Mississippi lors de la fonte des glaciers.
1.4 Lacunes et défis dans l’étude de la biodiversité et des
poissons d’eau douce
1.4.1 Identification des espèces
Les connaissances actuelles sur la biodiversité sont parsemées d‟incertitudes. Par exemple, bien que la diversité en espèces représente une mesure déjà trop simpliste pour décrire la biodiversité, le nombre total d‟espèces sur terre demeure inconnu et seulement une infime partie des espèces sont décrites (May 1988). En effet, on estime qu‟il existe entre 5 et 25 millions d‟espèces et moins de deux millions ont été décrites à ce jour (Lomolino et al. 2006). Considérant le contexte actuel de crise majeure d‟extinction (Vitousek et al. 1997, Sala et al. 2000, Jelks et al. 2008), il est inévitable qu‟une partie de la biodiversité disparaitra avant d‟avoir été étudiée ou même nommée (« Linnean shortfall »; Lomolino et al. 2006). Afin d‟approfondir nos connaissances fondamentales sur la biodiversité ainsi que pour conserver adéquatement cette diversité, il semble indispensable de décrire et d‟être en mesure d‟identifier les composantes de la diversité.
De nouvelles espèces sont décrites pour la première fois à chaque année. Parmi les groupes taxonomiques moins étudiés ou particulièrement diversifiés ainsi que dans les régions reculées, de nouvelles espèces, genres ou même familles sont régulièrement découverts (Stork 1988, Rosenzweig 1995, Bouchet et al. 2002). Plusieurs de ces nouveaux taxons présentent des formes absolument nouvelles. Parmi les régions et taxons ayant plus largement été étudiés par les taxonomistes, mais aussi parmi les autres taxons et régions, plusieurs des nouvelles espèces sont cryptiques et ne présentent pas ou peu de différences morphologiques avec les autres espèces ayant déjà été décrites (Bickford et al. 2007). Il apparait que des espèces répondant à chacun des concepts d‟espèces biologiques, phylogénétiques et écologiques peuvent passer inaperçues à un examen morphologique comme traditionnellement préconisé en taxonomie (Bickford et al. 2007). Néanmoins, certains cas de division peuvent sembler être basés sur des différences trop minimes, augmentant ainsi « artificiellement » la diversité en espèces, ce qui a mené Alroy (2003) à introduire le terme d‟« inflation taxonomique ». Deux écoles de pensées se confrontent sur le sujet; les diviseurs (« splitters ») et les réunisseurs (« lumpers ») d‟espèces. D‟un côté, les diviseurs se basent souvent sur le principe de précaution et affirment que leur approche s‟assure de ne pas mettre de côté des espèces méritant d‟être reconnues et conservées. D‟un autre côté, les réunisseurs peuvent préconiser une taxonomie plus stable et basée sur des caractères morphologiques bien visibles. Puisque les ressources sont limitées en conservation, les réunisseurs stipulent aussi parfois qu‟il est inopportun de reconnaitre une espèce sur des bases plus ou moins solides et qu‟il est plus avantageux d‟investir les efforts de conservation sur les espèces clairement distinctes. Un manque de standardisation dans la taxonomie peut donc affecter l‟efficacité des plans de gestion
environnementale. Une taxonomie non standardisée peut aussi biaiser les études comparatives (Agapow et al. 2004).
Même pour les espèces décrites, l‟identification des espèces peut être très laborieuse. Chez les animaux, la majorité des descriptions et des identifications est basée sur des caractères morphologiques. Bien que cette approche se soit avérée efficace dans plusieurs cas, il existe plusieurs contraintes à l‟emploi de caractères morphologiques afin d‟identifier les espèces. Premièrement, il peut être extrêmement difficile de distinguer des espèces ayant des caractères morphologiques très semblables ou présentant beaucoup de variabilités phénotypiques (ex. : Cottus cognatus et C. bairdi, Scott et Crossman 1998). Il y a souvent un nombre limité de taxonomistes habilités à identifier un taxon particulier. Deuxièmement, plusieurs espèces montrent de profonds changements ontogéniques. Chez les poissons, la majorité des clefs d‟identification n‟est pas applicable aux stades juvéniles et encore moins aux stades larvaires ou embryonnaires. Pour plusieurs lamproies par exemple, il n‟y a toujours pas de clef d‟identification permettant de distinguer les ammocètes (Neave et al. 2007). Chez plusieurs espèces de poissons marins, des chercheurs ont dû tenter d‟élever des larves en bassin, parfois sans succès, afin de pouvoir identifier leurs captures (Leis et Carson-Ewart 2000). Troisièmement, il est courant d‟avoir à effectuer des identifications chez des individus dont les caractères morphologiques sont dégradés ou transformés. Par exemple, il peut être pertinent de vérifier quelles espèces sont déchiquetées dans les turbines d‟une centrale hydroélectrique, quelle est l‟exactitude des étiquettes associées aux produits transformés de poissons tels que les filets ou les sushis (Baker et al. 1996, Wong et Hanner 2008) ou de déterminer l‟alimentation d‟un animal en identifiant le contenu de son estomac ou de ses excréments (Deagle et Tollit 2007). Enfin, plusieurs espèces cryptiques ne sont pas détectables à l‟aide d‟analyses morphologiques (Smith et al. 2006). Il apparait donc que les limites des identifications basées sur la morphologie restreignent l‟acquisition de connaissances dans des domaines aussi variés que l‟étude de la distribution spatiale des stades de vie, les relations écologiques, la spéciation et, bien sûr, la conservation.
Différentes techniques d‟identification ont été développées afin de faire face aux difficultés inhérentes à l‟utilisation exclusive de caractères morphologiques (Burton 1996, Céspedes et al. 1998), mais le développement de nombreux projets indépendants pose toutefois quelques sérieux problèmes. Premièrement, les outils d‟identification développés précédemment couvrent un nombre très limité d‟espèces. Deuxièmement, plusieurs groupes travaillent à l‟aide de marqueurs génétiques différents (Baker et al. 1996, Burton 1996, Céspedes et al. 1998, Palumbi et Cipriano 1998). L‟identification d‟un spécimen peut donc nécessiter de nombreuses analyses différentes, dont plusieurs n‟apportent aucune information pertinente en plus d‟être couteuse en temps et en argent. Troisièmement, il existe une possibilité d‟erreur d‟identification lors du développement de l‟outil. En effet, les spécimens ayant servi à l‟élaboration de l‟outil ne sont que très rarement
conservés dans une collection, ne permettant ainsi aucune vérification. Souvent aussi, et en particulier dans les banques publiques de séquences telles que GenBank, les identifications ne sont pas effectuées par des taxonomistes spécialisés dans le taxon en question (Seabra et al. 2010). Il semble donc qu‟un manque de couverture et de standardisation complique les analyses et laisse parfois planer un doute sur la validité des identifications.
La technique d‟identification d‟espèces appelée « codes-barres génétiques » (DNA barcoding) est basée sur l‟utilisation d‟une ou de quelques courtes séquences de gènes et présente plusieurs avantages sur les techniques traditionnelles basées sur la morphologie, en plus de remédier aux principaux problèmes des techniques d‟identification moléculaire développées précédemment (Hebert et al. 2003). Premièrement, la technique de codes-barres génétiques propose de standardiser l‟emploi d‟un seul segment d‟ADN afin d‟identifier l‟ensemble des animaux. Deuxièmement, la majorité des séquences est associée à un spécimen ayant été examiné par des taxonomistes spécialisés dans le taxon en question, et surtout le spécimen est entreposé dans une collection accessible à des fins de vérification (Ward et al. 2009). Les échantillons sont séquencés de façon bidirectionnelle et les électrophérogrammes sont enregistrés et disponibles sur Internet, toujours à des fins de vérification éventuelle. De plus, la base de données est accessible gratuitement sur Internet dans un format simple d‟utilisation (www.barcodinglife.org). Chez les animaux, l‟utilisation d‟un fragment d‟environ 650 paires de bases de la région 5‟ du gène mitochondrial COI s‟est avérée efficace pour distinguer près de 95 % des espèces testées, en plus de révéler des cas d‟espèces cryptiques (Hebert et al. 2004, Ward et al. 2005, Hebert et Landry 2010). En effet, il semble que pour une vaste partie de la diversité animale, le niveau de divergence génétique observé entre espèces sœurs, telles que reconnues par la taxonomie actuelle, est suffisamment important pour les discriminer. De plus, le pourcentage de divergence génétique observé à l‟interieur des espèces est généralement plus faible que le pourcentage de divergence génétique observé entre espèces définies par des traits morphologiques. Il serait donc possible d‟établir un seuil de divergence d‟ADN mitochondrial qui délimiterait les niveaux taxonomiques intraspécifique et interspécifique (Hebert et al. 2004, Ward et al. 2005). Il semble donc que le développement d‟une banque de données références de codes-barres génétiques pour les poissons d‟eau douce de l‟Amérique du Nord pourrait s‟avérer un outil précieux à bien des égards.
1.4.2 Patrons de répartition spatiale de la biodiversité
Les patrons généraux de répartition de la biodiversité demeurent en bonne partie inexpliqués (Gaston 2000, Mittelback et al. 2007). Par exemple, pour la majorité des organismes vivants, on observe une plus grande diversité en espèces près de l‟équateur comparativement aux régions polaires (Mittelback et al. 2007). Bien
que ce gradient latitudinal de biodiversité représente probablement le patron à grande échelle le plus étonnant et le plus longuement étudié en écologie (Wallace 1878, Hawkins 2001, Gaston 2000, Mittelback et al. 2007), les théories expliquant ce phénomène ne font toujours pas consensus. Parmi les grandes hypothèses générales ayant été proposées, une première stipule que les cycles glaciaires aient pu, de façon récurrente, éliminer la diversité située près des pôles et aient eu un impact graduellement moins important en s‟approchant de l‟équateur (Wallace 1878, Mittelback et al. 2007). Une deuxième hypothèse suggère que l‟activité métabolique, plus élevée à des températures chaudes et donc plus près de l‟équateur, pourrait influencer les taux de mutation et les taux de spéciation (Martin et Palumbi 1993, Allen et al. 2006, Mittelback et al. 2007, Dowle et al. 2013). Plus précisément, une activité métabolique élevée pourrait entrainer une plus grande consommation d‟oxygène et donc une plus grande production de radicaux libres, lesquels entraineraient un taux de mutation plus élevé. Considérant que le développement d‟incompatibilités génétiques de type Dobzanski-Muller représente une étape le plus souvent irréversible de la spéciation, et que l‟on s‟attend à ce que différentes mutations apparaissent dans différentes populations ou écotypes, les modèles mathématiques de spéciation indiquent qu‟un taux de mutation plus élevé devrait se traduire par un taux de spéciation plus élevé (Gavrilets 2003). Ainsi, une meilleure compréhension des mécanismes évolutifs régissant la répartition de la diversité pourrait non seulement aider à la conservation biologique, mais pourrait même aussi permettre une meilleure calibration de l‟horloge moléculaire (Gillooly et al. 2005).
La compréhension des rôles relatifs des cycles glaciaires et de l‟activité métabolique sur l‟évolution du patron de répartition de la biodiversité ne nécessite pas seulement d‟étudier la diversité en espèces, mais aussi la diversité à l‟intérieur des espèces et la diversité entre les espèces. En effet, un patron de variation intraspécifique et interspécifique congruant, en relation avec les patrons de diversité spécifique déjà largement documentés, constituerait une démonstration particulièrement convaincante du rôle de différents facteurs sur l‟évolution de la biodiversité. Jusqu‟à présent, il n‟y a toutefois que peu d‟études analysant simultanément la divergence à ces deux niveaux de divergence bordant le niveau spécifique (mais voir Bernatchez et Wilson 1998, Weir et Schluter 2007). Encore plus rarement, ce type d‟étude a été réalisé à l‟échelle d‟un continent et sur une faune diversifiée. Souvent, les études à très large échelle ont souffert de l‟existence présumée d‟un biais géographique dans la proportion de diversité n‟ayant toujours pas été décrite, comme par exemple les études couvrant des régions tropicales qui n‟ont pas été intensivement étudiées par les taxonomistes (Tobias et al. 2008).
Les poissons d‟eau douce de l‟Amérique du Nord représentent un bon modèle pour étudier les facteurs influençant les patrons de répartition de la biodiversité. En effet, cette faune est diversifiée et la majorité de ses quelques 900 espèces est endémique à ce continent (Nelson et al. 2004, Hocutt et Wiley 1986). Étant
donné la structure des habitats aquatiques d‟eau douce, les poissons de cette faune ont une capacité de dispersion limitée et représentent probablement un des groupes de Vertébrés les plus structurés génétiquement (Ward et al. 1994). Ainsi, la dispersion à partir des tropiques, comme il est suggéré par Jablonski et al. (2006) pour expliquer le gradient latitudinal, pourrait être un facteur beaucoup moins important chez les poissons d‟eau douce que chez les organismes plus mobiles tels que les poissons marins et les oiseaux. De plus, l‟ensemble de cette zone a aussi été largement étudié par de nombreux taxonomistes (Mayden 1992, Scott et Crossman 1998, Hubbs et Lagler 2004). Il n‟y a donc pas de raison évidente de croire qu‟il existe un important biais géographique dans la répartition des espèces cryptiques ou d‟inflation taxonomique, comme il est soupçonné dans le cas d‟analyses incluant des taxons et des régions peu étudiées (Tobias et al. 2008, Bickford et al. 2007, Giam et al. 2011). Enfin, les poissons d‟eau douce de l‟Amérique du Nord sont aussi caractérisés par un important gradient latitudinal. En effet, le nombre d‟espèces varie de 500 dans le sud des États-Unis à une seule espèce dans l‟extrême nord du Canada. Une étude précédente (Bernatchez et Wilson 1998), basée sur 42 espèces, a de plus décrit un gradient latitudinal de diversité génétique intraspécifique et interspécifique parmi cette faune. Enfin, une autre étude (Estabrook et al. 2007), basée sur 54 espèces de poissons nord-américains, suggère qu‟il puisse exister une relation entre l‟activité métabolique et les taux de substitution nucléotidique. L‟analyse des patrons de divergences génétiques intraspécifique et interspécifique de la quasi-totalité des poissons d‟eau douce de l‟Amérique du Nord pourrait donc vraisemblablement permettre de mieux comprendre les facteurs influençant la répartition de la biodiversité.
1.4.3 Spéciation allopatrique et pompe à spéciation
La spéciation allopatrique est considérée comme le mode de diversification le plus commun chez les animaux (Coyne et Orr 2004). Toutefois, les cas de spéciation allopatrique ont le plus souvent été inférés de façon indirecte par la concordance entre les frontières délimitant l‟aire de répartition de deux espèces sœurs et une barrière à la dispersion (Jordan 1908, Coyne et Orr 2004). Le meilleur test de spéciation allopatrique consiste à démontrer que deux groupes ont divergé en allopatrie et demeurent reproductivement isolés lorsqu‟ils se retrouvent en sympatrie dans une zone de contact secondaire (Coyne et Orr 2004). Ce type d‟étude demeure toutefois peu fréquent et n‟est que très rarement effectué sur plusieurs espèces codistribuées. Il est donc difficile de juger de la généralité de la spéciation allopatrique au sein d‟une faune en particulier. Une démonstration particulièrement convaincante de la généralité du processus de spéciation allopatrique consisterait à montrer que plusieurs espèces codistribuées sont rendues à différentes étapes du processus de spéciation allopatrique. En effet, si chacune des étapes du processus est observable au sein de diverses
espèces, il est raisonnable de croire que le processus entier peut se produire chez une seule espèce et que ce phénomène est commun (Jiggins et Mallet 2000, Coyne et Orr 2004).
Les zones de sutures, définies par Remington (1968) par l‟occurrence de plusieurs zones hybrides impliquant différents taxons dans une même région géographique restreinte, représentent un parfait laboratoire pour l‟étude de la spéciation et de l‟introgression. Chacune des zones de contact secondaire représente un test d‟isolement reproducteur. Toutefois, peu de zones de suture ont été répertoriées (Swenson et Howard 2004) et rarement elles ont été étudiées dans un contexte de spéciation (mais voir Moritz et al. 2009, Dasmahapatra et al. 2010). En effet, il semble nécessaire que plusieurs espèces répondent de façon similaire aux mêmes changements environnementaux pour que plusieurs zones hybrides se chevauchent à un endroit donné. Bien que cette situation soit possible (Swenson et Howard 2004), plusieurs études suggèrent plutôt que les espèces répondent souvent de façon individuelle et indépendante face aux mêmes changements (Graham et al. 1996, Soltis et al. 2006, Dasmahapatra et al. 2010). D‟un point de vue fondamental, il est pertinent de chercher à savoir si des espèces confrontées aux mêmes changements environnementaux ont répondu de façon similaire ou indépendante, cette réponse étant reflétée dans la structure phylogéographique (Avise 2000). D‟un point de vue plus pratique, une concordance au niveau de la répartition spatiale des lignées évolutives au sein des espèces faciliterait la mise en place de plans de conservation, par exemple via la protection de régions présentant un niveau d‟endémisme de lignées phylogénétiques particulièrement élevé (Moritz et Faith 2002, Taberlet et Cheddadi 2002, Davis et al. 2008).
Les études phylogéographiques comparées ont permis de répondre à certaines questions concernant l‟histoire évolutive des espèces (Bermingham et Moritz 1998), mais elles ont tout de même certaines lacunes. Premièrement, plusieurs études ne comparent la structure génétique que de deux ou trois espèces (ex. Lejeusne et al. 2011), ce qui est un nombre plutôt faible pour effectuer des généralisations. Deuxièmement, un nombre important d‟études phylogéographiques comparées est basé sur une revue de littérature et compare donc des résultats obtenus à l‟aide de différents marqueurs génétiques (Calsbeek et al. 2003). Toutefois, la comparaison de données provenant des mêmes marqueurs génétiques a le potentiel d‟avoir une meilleure résolution. Troisièmement, il peut être extrêmement intéressant d‟effectuer une analyse phylogéographique comparée dans une optique exploratoire. Cependant, le fait de tester les patrons de variation génétique déjà soupçonnés chez des organismes et dans une région déjà largement étudiée d‟un point de vue biogéographique offre le potentiel de pousser les interprétations à un niveau plus pointu, comme par exemple chez les drosophiles d‟Hawaï (Carson et Kaneshiro 1976, O‟Grady et DeSalle 2008). Enfin, plusieurs projets ont ciblé des espèces d‟une grande importance économique et, en particulier chez les poissons d‟eau douce nordiques du Néarctique, la structure génétique de plusieurs espèces parmi les plus communes et donc parmi
les plus importantes pour l‟écosystème demeure inconnue. Par exemple, plusieurs des premières analyses phylogéographiques effectuées dans la région anciennement glacée du Néarctique ont été réalisées chez les salmonidés, lesquelles ont suggéré que le niveau de concordance soit moindre chez les espèces de régions glacées durant le Pléistocène que dans les régions plus équatoriennes (Bernatchez et Wilson 1998). Il est toutefois probable que les salmonidés puissent souvent posséder d‟importantes capacités de dispersion en plus d‟être physiologiquement très efficaces à de basses températures, ce qui pourrait faire en sorte qu‟ils ne soient que peu représentatifs de l‟ensemble de la faune ichthyenne.
Les cycles glaciaires du Pléistocène pourraient avoir mené plusieurs espèces à répondre aux changements climatiques de façon similaire et avoir favorisé la divergence allopatrique. En effet, il semble possible que l‟avancée des glaciers ait pu mener à l‟isolation de plusieurs groupes d‟individus initialement similaires à diverger en allopatrie et à accumuler des incompatibilités génétiques ou à développer d‟autres mécanismes d‟isolement reproducteur. Un certain nombre d‟études appuie cette hypothèse qui suggère que les cycles glaciaires puissent agir comme une « pompe à spéciation » (sensu Haffer 1969) (Avise et al. 1998, Weir et Schluter 2004, Johnson et Cicero 2004). Toutefois, plusieurs autres études révèlent que la divergence d‟espèces sœurs précède très souvent le Pléistocène, suggérant ainsi que les cycles glaciaires du Pléistocène n‟aient pas eu un impact majeur sur la diversification spécifique (Klicka et Zink 1997, Knapp et Mallet 2003). La situation n‟est donc pas claire.
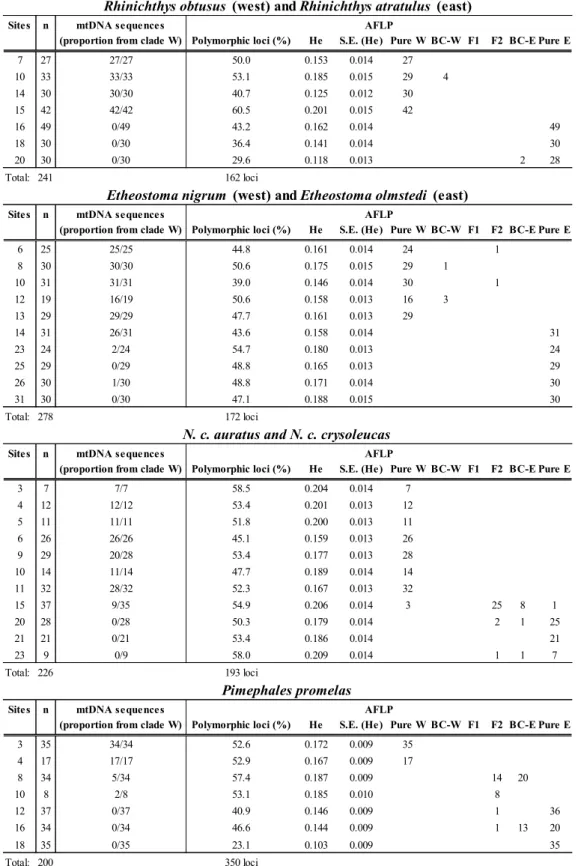
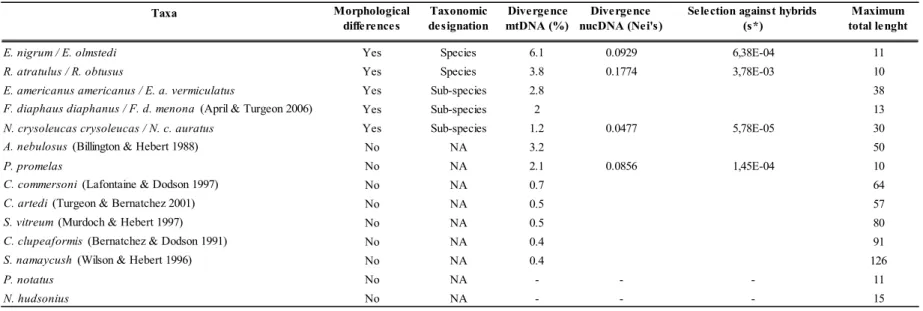
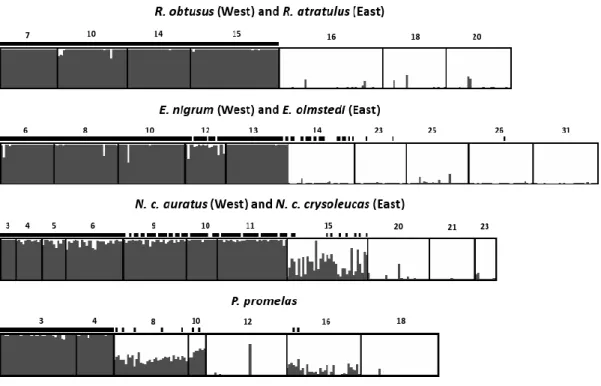
Les poissons d‟eau douce du nord-est américain pourraient constituer un excellent modèle pour tester le modèle de pompe à spéciation allopatrique. En effet, les études zoogéographiques portent à croire que deux refuges glaciaires ont été particulièrement importants pour ces poissons : le refuge mississippien, situé en amont du fleuve Mississippi à l‟ouest des Appalaches, et le refuge atlantique, situé sur la côte atlantique près du Rhode Island, à l‟est des Appalaches (Mandrak et Crossman 1998, Bernatchez et Wilson 1998). Plus de 30 espèces de poissons présentes dans le bassin des Grands Lacs et du fleuve Saint-Laurent pourraient avoir utilisé à la fois les refuges mississippien et atlantique (Hocutt et Wiley 1986). L‟isolement dans ces refuges glaciaires a permis aux groupes de poissons les ayant occupés de diverger (Hocutt et Wiley 1986, Bernatchez et Wilson 1998). Cette divergence est notamment bien représentée au niveau de l‟ADN mitochondrial (ADNmt) chez la majorité des organismes étudiés (Bernatchez et Wilson 1998, April et Turgeon 2006). Dans une moindre mesure, l‟isolation semble avoir permis à certains groupes de poissons de diverger au niveau de traits morphologiques. Certaines lignées glaciaires sont suffisamment différenciées pour être maintenant considérées comme des sous-espèces ou des espèces distinctes (Bailey et Smith 1981, Underhill 1986), telles que Esox americanus americanus/E. a. vermiculatus, Rhinichthys atratulus/R. obtusus, Etheostoma nigrum/E. olmstedi, Fundulus diaphanus diaphanus/F. d. menona.
Plusieurs taxons ayant divergé dans les refuges glaciaires atlantique et mississippien semblent former une zone de contact secondaire permettant de tester la présence d‟isolement reproducteur. Les études phylogéographiques et l‟analyse de la variation des caractères morphologiques montrent que plusieurs poissons ayant utilisé les refuges atlantique et mississippien pourraient former une zone de suture dans le grand bassin des Grands Lacs et du fleuve Saint-Laurent. En effet, plusieurs espèces semblent avoir une structure populationnelle relativement similaire dans cette zone (Murdoch et Hebert 1997, April et Turgeon 2006). Parmi les taxons reconnus pour avoir divergé morphologiquement, seul F. diaphanus a été le sujet d‟une analyse phylogéographique. L‟étude réalisée confirme que les deux sous-espèces sont vraisemblablement issues des refuges atlantique et mississippien et qu‟elles s‟hybrident dans le fleuve Saint-Laurent et dans l‟est des Grands Lacs (April et Turgeon 2006). Chez les autres taxons caractérisés par l‟occurrence de sous-espèces ou espèces associées aux refuges glaciaires, la variation des caractères morphologiques suggère fortement que les lignées se rencontrent aussi dans la région du lac Ontario et du fleuve Saint-Laurent : R. atratulus/R. obtusus, E. a. americanus/E. a. vermiculatus et E. nigrum/E. olmstedi (Lee et al. 1980). Mis à part le complexe E. nigrum/E. olmstedi, pour lequel des analyses suggèrent que les deux espèces demeurent morphologiquement distinctes en sympatrie (Chapleau et Pageau 1985), et bien que peu d‟analyses poussées ne semblent avoir été effectuées pour répondre spécifiquement à cette question, certains auteurs suggèrent que les lignées se mélangent lorsqu‟en sympatrie dans les zones de contact secondaire et qu‟il n‟y ait pas de barrières reproductives entre les lignées (Bailey et Smith 1981, Hocutt et Wiley 1986, Reist et Crossman 1987). Il semble donc que les espèces de poissons ayant utilisé à la fois les refuges glaciaires de l‟Atlantique et du Mississippi représentent un excellent système d‟étude permettant de tester le modèle de pompe à spéciation allopatrique.
1.5 Objectifs
L‟objectif général de cette thèse est d‟approfondir les connaissances fondamentales sur la biodiversité et l‟évolution des poissons d‟eau douce de l‟Amérique du Nord et d‟appliquer ces connaissances à la conservation de cette richesse. Pour ce faire, une approche génétique et comparative sera utilisée afin d‟étudier les niveaux intraspécifique, spécifique et interspécifique de cette faune. Premièrement, une banque de séquences d‟ADNmt représentative de la diversité spécifique des poissons d‟eau douce du Canada et des États-Unis sera développée afin de tester et de mettre en place un outil d‟identification basé sur le principe des codes-barres génétiques. Il sera alors possible de tester l‟hypothèse stipulant que les espèces décrites sur des bases morphologiques correspondent à des clades génétiques distincts. Cette étude permettra donc non seulement de fournir un précieux outil d‟identification, mais l‟analyse des niveaux de divergences génétiques
intraspécifique et interspécifique permettra aussi de mettre en lumière les possibles failles de la classification taxonomique actuelle. Deuxièmement, la collection de séquences d‟ADNmt provenant de la majorité des espèces de poissons de cette région sera utilisée afin de tester l‟effet hypothétique des cycles glaciaires du Pléistocène et de l‟activité métabolique sur les niveaux de divergences génétiques intraspécifique et interspécifique. Cette étude permettra donc de mieux comprendre les mécanismes évolutifs régissant les patrons globaux de répartition de la biodiversité. Troisièmement, la variation spatiale des caractères génétiques mitochondriaux et nucléaires sera analysée chez plusieurs espèces ayant utilisé les mêmes refuges glaciaires afin de vérifier l‟importance du phénomène de spéciation allopatrique au sein des poissons d‟eau douce. Cette approche permettra de valider l‟existence d‟une zone de suture dans le système du fleuve Saint-Laurent et des Grands Lacs ainsi que de tester le modèle de pompe à spéciation causée par les cycles glaciaires.
1.6 Marqueurs génétiques
Les marqueurs génétiques nécessitant relativement peu de travail de développement et étant peu couteux sont particulièrement utiles pour les études effectuées sur un très grand nombre d‟individus et de taxons non modèles. C‟est pourquoi le choix des outils moléculaires s‟est arrêté sur les séquences mitochondriales du gène COI et les marqueurs de type AFLP.
L‟ADNmt s‟avère très utile afin de révéler l‟histoire évolutive des animaux et pour faire le pont entre la microévolution et la macroévolution (Avise 2000). En effet, plusieurs caractéristiques font de cette molécule un outil d‟analyse de choix. Premièrement, les molécules d‟ADNmt sont présentes en grand nombre dans les cellules animales, ce qui en facilite l‟extraction. Deuxièmement, l‟ADNmt évolue relativement rapidement en comparaison avec la majorité des séquences d‟ADNnuc, ce qui rend souvent cette partie du génome variable et donc informative (Brown et al. 1979). En particulier, son taux de mutation permet de différencier adéquatement les lignées évolutives divergeant depuis quelques dizaines de milliers d‟années à quelques millions d‟années (Brown et al. 1979, Avise 2000), ce qui correspond souvent aux niveaux de divergence observés entre les vieilles lignées intraspécifiques et les jeunes espèces (Avise et al. 1998, Hebert et al. 2003). Troisièmement, l‟alignement et l‟analyse des séquences d‟ADNmt sont relativement simples puisque d‟une part, la molécule est uniquement transmise par la mère chez la majorité des animaux et n‟est généralement pas soumise à la recombinaison et, d‟autre part, les insertions et délétions de nucléotides semblent extrêmement rares. Quatrièmement, des amorces permettant l‟amplification de sections d‟ADNmt par PCR ont été développées chez la majorité des animaux et pour l‟ensemble des poissons (ex. : Ivanova et al. 2007). Enfin, Paul Hebert et ses collaborateurs (2003) ont proposé de standardiser l‟utilisation d‟une
séquence mitochondriale du gène COI afin d‟identifier les espèces. Cette standardisation rend le développement d‟un outil d‟identification moléculaire global plus efficace et permet de comparer plus justement les patrons observés dans diverses études évolutives.
Les marqueurs de type AFLP permettent d‟obtenir de l‟information sur une très grande quantité de loci répartis de façon aléatoire dans le génome sans nécessiter de laborieuses étapes de développement (Vos et al. 1995, Bensch et Aakesson 2005). Pour ce faire, l‟ADN génomique est d‟abord digéré à l‟aide de deux enzymes de restriction produisant plusieurs fragments de longueurs différentes. Ensuite, des adaptateurs oligonucléotidiques sont liés à ces fragments. En utilisant des amorces sélectives, il est alors possible d‟amplifier une fraction de ces fragments. Le produit obtenu par PCR migre ensuite au travers d‟un polymère afin que les fragments se séparent en fonction de leurs longueurs. En comparant les profils de migration de différents individus, il devient possible de déterminer la présence ou l‟absence des fragments. Les AFLP sont donc des marqueurs de type dominant qui demeurent anonymes (Vos et al. 1995).
![[PDF] Cours PHP : les Cookies comment ça marche | Cours informatique](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)