Control of larval hematopoiesis in Drosophila; microenvironment, precursors and cell lineage
166
0
0
Texte intégral
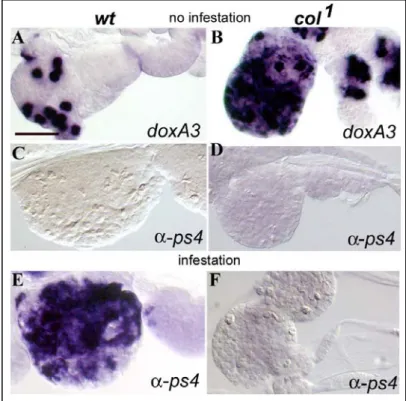
Figure


+7



Documents relatifs