Climat Holocène du Nunatsiavut inféré à partir
des assemblages sédimentaires de pollen et
de kystes de dinoflagellés des fjords
(Labrador, Canada)
Thèse
Thomas RICHEROL
Doctorat en sciences géographiques
Philosophiæ doctor (Ph.D.)
Québec, Canada
iii
Résumé
L’accélération du développement industriel a eu des répercussions importantes sur le climat planétaire, en particulier sur le réchauffement des pôles au cours des dernières décennies. Afin d’anticiper les changements climatiques futurs, il faut comprendre la variabilité naturelle actuelle et passée du climat. Dans le cadre de ma thèse doctorale, je me suis intéressé à l’histoire climatique du Nunatsiavut, une région subarctique peu documentée du Labrador (Canada). L’analyse palynologique des enregistrements géologiques conservés dans les sédiments océaniques permet d’établir un lien entre les paramètres environnementaux modernes et passés caractérisant les conditions de surface océanique (température, salinité, durée du couvert de glace, productivité primaire). Tout d’abord, j’ai décrit les assemblages modernes de kystes de dinoflagellés (= dinokystes) de quatre fjords du Nunatsiavut (Nachvak, Saglek, Okak et Anaktalak) localisés le long d’un gradient latitudinal. L’apparition et la dominance de kystes de dinoflagellés autotrophes caractérisent les fjords du Sud (Okak et Anaktalak). La présence dans le fjord d’Anaktalak de kystes calcaires de
Scrippsiella cf. S. crystallina, indicateur de pollution anthropique, illustre
vraisemblablement l’impact négatif de la mine de nickel récemment implantée dans la région. Ensuite, j’ai analysé de séquences sédimentaires courtes (25-30 cm) prélevées à Nachvak, Saglek et Anaktalak. Les assemblages fossiles de dinokystes et du pollen décrivent l’histoire climatique et océanique de la région au cours des derniers 150-300 ans. Bien que la tendance climatique en Arctique soit au réchauffement, les reconstitutions paléocéanographiques suggèrent une résilience climatique au Labrador, accompagnée d’un léger refroidissement des eaux de surface dans les fjords de Saglek et d’Anaktalak. Les assemblages fossiles du pollen illustrent ce refroidissement sur terre par un déplacement de la limite des arbres vers le sud. Finalement, l’analyse d’une séquence sédimentaire longue (812 cm) prélevée dans le fjord de Nachvak, en utilisant une approche multi-proxy combinant une solide chronologie AMS-14C et les assemblages de
climatique des 7 000 dernières années. Les reconstitutions paléocéanographiques dans le fjord de Nachvak révèlent un climat du début de l’Holocène influencé par l'Atlantique Nord, alors qu’une grande stabilité climatique est observée pour les 1 000 dernières années, probablement due à un effet de « barrière » du courant du Labrador qui charrie des eaux froides de l’Arctique.
v
Table des Matières
Résumé ... iii
Table des Matières ... v
Liste des Tableaux ... ix
Liste des Figures ... xi
Liste des Planches photos ... xvii
Remerciements ... xxi
Avant-propos ... xxv
Chapitre 1. Introduction générale. ... 1
1.1. Introduction ... 1
1.2. Sites d’étude ... 8
1.2.1. Localisation, climat et végétation ... 8
1.2.2. La déglaciation à l’Holocène au Labrador ... 14
1.3. Les kystes de dinoflagellés (= dinokystes) ... 16
1.4. Reconstitutions climatiques au Labrador et dans les régions avoisinantes ... 20
1.5. Références bibliographiques ... 26
Chapitre 2. Modern dinoflagellate cyst assemblages in surface sediments of Nunatsiavut fjords (Labrador, Canada). ... 33
Résumé ... 33 Abstract ... 34 2.1. Introduction ... 35 2.2. Environmental Settings ... 40 2.3. Methodology ... 45 2.3.1. Sampling ... 45 2.3.2. Sieving ... 47 2.3.3. Chemical processing ... 47 2.3.4. Palynomorph counts ... 47 2.3.5. Statistics ... 50 2.3.6. Environmental data ... 50
2.4. Results and Discussion ... 52
2.5. Conclusions ... 65
Acknowledgements ... 67
Chapitre 3. Recent anthropogenic and climatic history of Nunatsiavut fjords (Labrador, Canada). ... 73 Résumé ... 73 Abstract ... 74 3.1. Introduction ... 75 3.2. Environmental Settings ... 77 3.3. Methodology ... 81 3.3.1. Sampling process ... 81 3.3.2. Sedimentological analysis ... 82 3.3.3. Chronological framework ... 85 3.3.4. Palynomorphs preparations ... 86 3.3.5. Statistical analysis ... 87
3.3.6. Diatom laboratory procedure ... 90
3.4. Results ... 92
3.4.1. Chronological framework ... 92
3.4.2. Sedimentology ... 94
3.4.3. Palynomorph fluxes ... 96
3.4.4. Diatoms ... 99
3.4.5. Pollen and Spore Assemblages ... 102
3.4.6. Dinocyst Assemblages ... 104
3.4.7. Sea-surface quantitative reconstructions ... 109
3.5. Discussion ... 112
3.5.1. Environmental and anthropogenic history ... 112
3.5.2. Climatic history ... 115
3.6. Conclusions ... 119
Acknowledgements ... 120
References ... 121
Chapitre 4. Holocene climate history of the Nunatsiavut (northern Labrador, Canada) established from pollen and dinoflagellate cyst assemblages covering the past 7,000 years. ... 127
Résumé ... 127 Abstract ... 128 4.1. Introduction ... 129 4.2. Methodology ... 132 4.2.1. Sampling ... 132 4.2.2. Stratigraphic analysis ... 132 4.2.3. Chronological framework ... 133 4.2.4. Palynomorphs preparation ... 136 4.2.5. Statistical analyses ... 137 4.3. Results ... 139 4.3.1. Chronological framework ... 139 4.3.2. Sedimentology ... 140
vii
4.3.3. Palynomorph fluxes ... 142
4.3.4. Pollen and Spore Assemblages ... 145
4.3.5. Dinocyst Assemblages ... 147
4.3.6. Paleoclimatic reconstructions ... 150
4.4. Discussion ... 156
4.4.1. Holocene Thermal Maximum (HTM) – 5,800-7,000 cal BP ... 158
4.4.2. Neoglacial – 5,800-3,200 cal BP... 159
4.4.3. Roman Warm Period-Medieval Warm Period (RWP-MWP) – 3,200-1,000 cal BP ... 161
4.4.4. Industrial Era (IE) – the last 1,000 cal BP ... 162
4.5. Conclusions ... 163
Acknowledgements ... 164
References ... 165
Chapitre 5. Conclusion générale. ... 171
ix
Liste des Tableaux
Table 2.1 : Geographical coordinates, water depth (in meters), number of cyst counted, cyst concentration (cyst g-1) and hydrographic data for samples shown in Fig. 2A/B/C/D : SST=summer sea surface temperature (°C); S-SSS=summer sea surface salinity; ICE=sea-ice cover duration (months year-1);
PAR=photosynthetically available radiation (Einstein m-2 day-1); NUTRIENTS=average concentration of nitrates, nitrites, silicates and phosphates in the water column for 2006/2007/2009/2010 (µmol L-1);
DIATOMS=average concentration of diatoms in the water column for 2006/2009/2010 (number of cells cm-3). ... 46
Table 2.2 : List of dinoflagellate taxa present in our samples and their code names. ... 48 Table 2.3: Relative abundance (%) of dinoflagellate cyst taxa in our samples. See
Table 2 for the codenames of the dinocyst species and also : Selo=Spiniferites
elongatus; Sfri=Spiniferites frigidus; Imin=Islandinium minutum;
Imic=Islandinium minutum var. cezare; Ibre=Islandinium brevispinosum; Bcar=Brigantedinium cariacoense, Bsim=Brigantedinium simplex; Parc=Polykrikos Arctic Morphotype I; Pqua=Polykrikos Arctic Morphotype II (quadratus). ... 49 Table 3.1 : Details concerning the three studied cores: geographical coordinates,
water depth, length and diameter of each core. ... 81 Table 3.2 : Table showing dry bulk density (g cm-3), cumulative dry mass (g cm-2) and sedimentation rates (g cm-2 year-1 and cm year-1) and their respective standard deviation (σ) for core N602-1. The italic lines represent depths for which sedimentation rates have been extrapolated. ... 83 Table 3.3 : Table showing dry bulk density (g cm-3), cumulative dry mass (g cm-2) and sedimentation rates (g cm-2 year-1 and cm year-1) and their respective
standard deviations (σ) for core S617. The italic lines represent depths for which sedimentation rates have been extrapolated. ... 84 Table 3.4 : Table showing dry bulk density (g cm-3), cumulative dry mass (g cm-2)
and sedimentation rates (g cm-2 year-1 and cm year-1) and their respective
standard deviations (σ) for core V624. The italic lines represent depths for which sedimentation rates have been extrapolated. ... 85 Table 4.1 : Table detailing the statistical data of the calibration of the AMS-14C
dates obtained throughout the core. ... 135 Table 4.2 : Table summarizing the information on the parameters reconstructed
from assemblages of fossil pollen and dinocyst (from Richerol et al., 2014) : parameter denomination and codename, unit, coefficient de correlation (R2)
xi
Liste des Figures
Figure 1.1:Carte représentant le découpage des 4 régions IRIS (tiré de Allard and Lemay, 2012) ... 3 Figure 1.2 : Tendances des températures moyennes annuelles de 1948 à 2003
(Environnement Canada 2007) ... 5 Figure 1.3 : Principales zones de végétation du Labrador. ... 10 Figure 1.4 : Schéma du cycle de vie des dinoflagellés. A) phase haploïde durant
laquelle les individus (1 dinoflagellé = 1 cellule) se divisent via la mitose. Il s’agit du stade de vie végétatif qui produit les événements de floraison ou
bloom. À la suite d’un signal interne et/ou externe, des gamètes se forment et
fusionneront pour produire un zygote diploïde; B) phase planozygote (zygote mobile pélagique) durant laquelle la cellule diploïde perdra éventuellement sa mobilité et développera un kyste qui sera éventuellement libéré dans la colonne d’eau puis sédimentera; C) phase hypnozygote durant laquelle le kyste subira une période de dormance obligatoire contrôlée par une horloge interne qui déclenchera le moment de la libération du dinoflagellé ou exkystement (adapté de Rochon, 2009). ... 16 Figure 2.1: Map of the Nunatsiavut (North Labrador, Canada) illustrating the
location of the four fjords involved in this study: A. Nachvak Fjord; B. Saglek Fjord; C. Okak Fjord and D. Anaktalak Fjord. ... 36 Figure 2.2: Maps of the four fjords showing the location of the surface sediment
samples used in our study. ... 39 Figure 2.3: Latitudinal distribution of the total concentration of dinocysts (cysts g-1)
in the fjords. ... 52 Figure 2.4: Relative abundance of major dinocyst taxa, hydrographic parameters
(temperature, salinity, sea-ice cover, nutrients and diatoms) and statistical assemblages of dinocysts. ... 59
Figure 2.5: Latitudinal distribution of the relative abundance (%) of cysts from autotrophic and heterotrophic dinoflagellates in our samples. ... 60 Figure 2.6: Graph of the Canonical Correspondence Analysis (CCA) between the
dinocyst species and six parameters : S-SST=summer sea-surface temperature; S-SSS=summer sea-surface salinity; ICE=sea-ice cover duration; PAR=photosynthetically available radiation; NUTRIENTS=average concentration of nitrates, nitrites, silicates and phosphates in the water column; DIATOMS=average concentration of diatoms in the water column. See Table 2 for the codenames of the dinocysts species. ... 62 Figure 3.1 : Map of the Nunatsiavut (North Labrador, Canada) illustrating the
location of the three fjords involved in this study: A. Nachvak Fjord; B. Saglek Fjord and C. Anaktalak Fjord and the nearby sea-surface circulation pattern in the Labrador Sea (modified from Richerol et al., 2012)... 76 Figure 3.2 : Detailed maps of the three fjords showing the precise location of the
three studied cores (modified from Richerol et al., 2012). ... 78 Figure 3.3 : Validation test of the ‘N = 1,441’ database for the four reconstructed
parameters : August Surface Temperature (SST, in °C), August Sea-Surface Salinity (SSS), Sea-Ice Cover duration (months year-1) and Annual
Productivity (g C m-2 year-1). RMSEP = Root Mean Square Error of Prediction.
... 90 Figure 3.4 : 210Pb profiles of the three cores used to established sedimentation
rates (g cm-2 year-1) and chronological frameworks (years A.D.). The curves of
the 210Pb total activity (Bq g-1) and natural logarithm of the unsupported 210Pb
activity are represented according to the depth expressed as Cumulative Dry Mass (g cm-2) in order to correct for sediment compaction. The vertical dotted
line on the first graph represents the estimated value of the supported 210Pb
activity. Standard deviations for ages obtained are also depicted. Gray dots for Saglek 617 core represent the two AMS-14C dates obtained at the base of the
xiii Figure 3.5 : Sedimentological analyses of the three cores represented in depth
scale (cm). From left to right: gray-level picture for each core ; grain-size analysis (mean, median and sorting (µm)) and percentages of silt and sand. For V624 core, grain-size analysis is discontinuous at 8.5 cm and 10.5 cm due to flushing and loss of the corresponding samples in the laser grain-size analyzer. ... 95 Figure 3.6 : Fluxes of the five types of palynomorphs counted, according to depth
(cm) and time (years A.D.), for each sediment core : dinocysts (cysts cm -2 year-1), terrestrial palynomorphs (grains cm-2 year-1), Halodinium sp.
(grains cm-2 year-1), foraminifer linings (linings cm-2 year-1) and pre-Quaternary
palynomorphs (grains cm-2 year-1). For each curve, the black thick line is a smooth on three values. Gray areas highlight the major features presented in the result section. ... 98 Figure 3.7 : Different water mass influences in the fjords illustrated through diatom
relative abundances (%) according to depth (cm) and time (years A.D.). ... 100 Figure 3.8 : Pollen and spore zones identified from the relative abundances (%) of
all the species identified and counted in the three cores, according to depth (cm) and time (years A.D.). Different shades of gray represent pollen zones obtained for each core using a cluster analysis. ... 103 Figure 3.9 : Dinocyst zones identified from the relative abundances (%) of major
species identified and counted in the three cores, according to depth (cm) and time (years A.D.). Different shades of gray represent dinocyst zones obtained for each core using a cluster analysis. ... 105 Figure 3.10 : Evolution of dinocyst fluxes (cysts cm-2 year-1) in parallel with the four
reconstructed oceanographic parameters, according to depth (cm) and time (years A.D.) : August SST (°C), August SSS, Sea-Ice Cover duration (months year-1) and Annual Productivity (g C m-2 year-1). For each curve, the
black thick line is a smooth on three values. Horizontal black dotted lines on dinocyst fluxes represent major shift in dinocyst assemblages. Vertical dotted lines represent the confidence interval for each reconstructed parameter at each depth. For each parameter, the black empty diamonds on the x-axis
represent modern values. Gray areas highlight the major features presented in the result section. The arrows in S617 illustrate the trends. ... 110 Figure 4.1 : Map of the Nunatsiavut (North Labrador, Canada) illustrating the
location of the piston core retrieved from Nachvak fjord (modified from Richerol et al., 2012). ... 131 Figure 4.2 : Age-depth model based on the AMS-14C chronology from the piston
core. ... 139 Figure 4.3 : Sedimentological analyses of the core represented on a depth scale
(cm). From left to right: grain-size analysis (mean, median and sorting (µm)), percentages of silt and sand, and volumetric magnetic susceptibility corrected for fractional porosity (SI). ... 141 Figure 4.4 : Fluxes of the five types of palynomorphs counted, according to time
(year cal BP): dinocyst (cysts cm-2 year-1), terrestrial palynomorphs (grains cm -2 year-1), Halodinium sp. (grains cm-2 year-1), foraminifer linings (linings cm -2 year-1) and pre-Quaternary palynomorphs (grains cm-2 year-1). For each
curve, the black thick line is a smooth on three values. ... 144 Figure 4.5 : Pollen and spore zones identified based on the relative abundances
(%) of major species observed in the core, according to time (year cal BP). Different shades of gray represent pollen zones obtained for each core using a cluster analysis. ... 146 Figure 4.6 : Dinocyst zones identified based on the relative abundances (%) of
major species observed in the core, according to time (year cal BP). Different shades of gray represent pollen zones obtained for each core using a cluster analysis. ... 149 Figure 4.7 : Graphic representation, according to the five pollen zones, of the
biomes types and origin of the analogs, and the mean monthly temperature (°C – the bars) and precipitation (mm – the line). The blue up arrow illustrate the cold northern origin of the analogs, and the red down arrow their warmer southern origin. ... 151 Figure 4.8 : Evolution of terrestrial palynomorph fluxes (grains cm-2 year-1) in
xv (year cal BP) : Annual Precipitation (mm) and Mean Temperature of the Coldest and Warmest Month (°C). For each curve, the black thick line is a smooth on three values. Horizontal black dotted lines on terrestrial palynomorphs fluxes represent major shift in pollen zones. Vertical dotted lines represent the confidence interval for each reconstructed parameter over time. For each parameter, the black empty diamonds on the x-axis represent modern values estimated from nearby meteorological stations (Cartwright 57°W and Goose A 60°W (Environment Canada)). HTM = Holocene Thermal Maximum; RWP = Roman Warm Period; MWP = Medieval Warm Period; IE = Industrial Era. ... 153 Figure 4.9 : Evolution of dinocyst fluxes (cysts cm-2 year-1) in parallel with the four
reconstructed oceanographic parameters, according to time (year cal BP) : August Sea-Surface Temperature (°C), August Sea-Surface Salinity, Sea-Ice Cover duration (months year-1) and Annual Productivity (g C m-2 year-1). For each curve, the black thick line is a smooth on three values. Horizontal black dotted lines on dinocyst fluxes represent major shift in dinocyst zones. Vertical dotted lines represent the confidence interval for each reconstructed parameter over time. For each parameter, the black empty diamonds on the x-axis represent modern values. HTM = Holocene Thermal Maximum; RWP = Roman Warm Period; MWP = Medieval Warm Period; IE = Industrial Era. ... 154 Figure 4.10 : Synthesis of the information gathered from fossil pollen and dinocyst
assemblages in relation to time (year cal BP). Left side of the figure: relative abundance (%) of pollen of trees, shrub-like trees, herbs and spores, annual precipitation (mm) and mean temperature of the coldest and warmest months (°C). Right side of the figure: relative abundances (%) of major species of dinocysts observed, August sea-surface temperature (°C) and salinity, and sea-ice cover duration (months year-1). Horizontal black dotted lines on
abundance graphs represent major shifts in pollen and dinocyst assemblage zones. Vertical dotted lines represent the confidence interval for each reconstructed parameter through time. HTM = Holocene Thermal Maximum;
RWP = Roman Warm Period; MWP = Medieval Warm Period; IE = Industrial Era. ... 157
xvii
Liste des Planches photos
Plate 2.1: Each scale bar represents 10 µm. A/C. Scrippsiella cf. S. crystallina station Okak-634, observed after chemical treatment using transmitted-light microscopy at 100x. B/D. Scrippsiella cf. S. crystallina station Okak-634, observed before chemical treatment with an inverse microscope at 40x. E.
Scrippsiella cf. S. crystallina station Anaktalak-624, observed after chemical
treatment using transmitted-light microscopy at 100x. F. Scrippsiella cf. S.
crystallina station Anaktalak-624, observed before chemical treatment with an
inverse microscope at 40x. ... 57 Plate 3.1 : Micrographs of palynomorphs (A and B), and cysts from autotrophic
dinoflagellate species (C to F). Each scale bar represents 10 μm ; transmitted-light microscopy at 100× magnification. A. Halodinium sp., core S617 (11-12 cm). B. Foraminifer lining, core V624 (0-1 cm). C. Cyst of
Pentapharsodinium dalei, core V624 (23-24 cm). D. Spiniferites ramosus, core
V624 (23-24 cm). E. Spiniferites elongatus, core V624 (17-18 cm). F.
Spiniferites frigidus, core V624 (14-15 cm). ... 107
Plate 3.2 : Micrographs of cysts from heterotrophic dinoflagellate species. Each scale bar represents 10 μm ; transmitted-light microscopy at 100× magnification. G. Islandinium minutum, core N602-1 (2-4 cm). H. Islandinium
minutum var. cezare, core N602-1 (10-12 cm). I. Islandinium brevispinosum,
core N602-1 (6-8 cm). J. Brigantedinium sp., core N602-1 (10-12 cm). K.
Brigantedinium simplex, core N602-1 (10-12 cm). L. Brigantedinium cariacoense, core N602-1 (10-12 cm). M. Echinidinium karaense, core N602-1
(24-25 cm). N. Selenopemphix quanta, core V624 (0-1 cm). O. Polykrikos Arctic Morphotype I, core S617 (4-5 cm). P. Polykrikos Arctic Morphotype II (quadratus), core S617 (0-1 cm). ... 108
xix « People assume that time is a strict progression of cause to effect, but actually, from a non-linear non-subjective viewpoint, it's more like a big bowl of wibbly wobbly timey wimey... stuff. »
xxi
Remerciements
Enfin!
La partie facile de la thèse, celle que mes superviseurs n’ont même pas besoin de relire. Tout le reste est rédigé, juste en attente des dernières corrections. Donc aujourd’hui c’est vendredi et je me détends avec mes remerciements... Ah, la vie palpitante d’un étudiant en fin de thèse!
Certains pourraient considérer que 7 ans pour un doctorat c’est long. On pourrait rétorquer qu’en recherche ce qui compte c’est le temps nécessaire à la maturation des idées, pas les années qui passent. Mais sérieusement, même moi j’ai trouvé ça long, très long, tant et si bien que par moment je pensais que ça ne finirait jamais. Depuis la première année, il y a sur mon bureau cette carte postale avec une citation prêtée à Woody Allen : « L’éternité c’est long, surtout vers la fin ». Ce n’est que dans les deux dernières années que j’ai vraiment compris le sens de cette phrase.
Quand on passe autant de temps sur un projet, on rencontre beaucoup de gens, des professeurs mais aussi d’autres étudiants à la maîtrise ou au doctorat, qui travaillent sur des sujets similaires ou complètement différents. On entend parler de certaines histoires, on échange sur comment les choses se passent dans différents laboratoires. On fait des observations sur la dynamique d’une thèse, la relation avec le superviseur, et on peut en retirer certaines satisfactions et reconnaissances.
Je suis reconnaissant de ne pas avoir eu des superviseurs qui fonctionnaient selon un rythme de fonctionnaire en travaillant de 9h à 17h, du lundi au vendredi, et ne consultaient pas leurs courriels les fins de semaine.
Je suis reconnaissant d’avoir eu des superviseurs qui ne prenaient pas des mois pour me renvoyer leurs commentaires sur mes articles et ne se contredisaient pas entre eux.
Je suis reconnaissant d’avoir eu des superviseurs qui prenaient au sérieux leur relecture de mes articles et n’attendaient pas la nième version pour faire des
commentaires fondamentaux.
Je suis reconnaissant de ne pas avoir eu des superviseurs qui considéraient qu’un étudiant au doctorat n’est globalement qu’un employé au rabais au service de leur recherche et qu’en tant que tel il n’a pas vraiment voie au chapitre sur l’orientation des interprétations.
Je suis reconnaissant d’avoir pu développer mes idées et mon style dans mes articles sans que mes superviseurs sortent la carte de l’expérience pour imposer leur vision de la recherche sans discussion possible.
Je suis reconnaissant pour le sujet que m’ont offert mes superviseurs, impliquant la chance d’aller toucher du doigt les paysages magnifiques du Nunatsiavut. Même si mon terrain a été trop court à mon goût, mon bref été à Saglek et ma traversée des fjords sur l’Amundsen restent parmi mes meilleurs souvenirs de ces 7 années.
Je suis reconnaissant pour la patience et la persévérance de mes superviseurs malgré les années qui passaient. Ils ne m’ont jamais abandonné.
Pour toutes ces choses et bien d’autres je remercie mes superviseurs Reinhard Pienitz (Département de Géographie – Université Laval) et André Rochon (ISMER – UQAR) de m’avoir fourni l’opportunité de développer ce projet et de m’avoir si bien accompagné tout du long.
Finalement la fin est là, mais elle aurait pu tarder un peu plus sans la participation des membres de mon comité qui ont accepté de travailler en un temps record sur l’évaluation de ma thèse. Je les remercie d’avoir accepté de jongler avec leurs emplois du temps chargés afin que je puisse soutenir à la date choisie qui arrangeait tout le monde. Pour tous ces efforts et pour leur travail d’évaluation je remercie Anne de Vernal (GEOTOP – UQAM), Fabienne Marret (Département de Géographie – Université de Liverpool) et Warwick Vincent (Département de Biologie – Université Laval). Je tiens aussi à remercier Guillaume Massé
xxiii (Département de Biologie – Université Laval) qui a accepté de réaliser la pré-lecture de ma thèse, également en un temps très court.
Un immense merci aussi à Ghislain Côté. Ghislain a été un excellent assistant de terrain lors de notre été au Nunatsiavut. Ensemble nous avons pu profiter de l’accueil chaleureux des habitants de Nain durant notre séjour prolongé à l’occasion de la saison surprise « fog patches along the coast ». Ghislain a également été un technicien de labo très efficace lorsqu’il a traité les échantillons que nous avions rapportés. Ghislain a offert à un malheureux maudit Français la chaleur de sa famille pour les fêtes de fin d’année durant les 3 premières années de mon doctorat. Ghislain est avant tout devenu un excellent ami. Pour toutes ces raisons il a amplement mérité sa place dans ces remerciements.
Il est une personne très chère à mon coeur qui a également joué une part importante dans la finalisation de ce doctorat : Virginie Galindo. Ma blonde, ma copine, ma conjointe, mon coeur, mon amour, la femme qui est entrée en coup de vent et par suprise dans ma vie un soir de novembre 2009, et qui a accepté de s’y installer pour un bout, pour toujours au coeur de l’été 2010. Je t’ai rencontrée durant ma traversée du désert, alors que j’avais atteint le point le plus bas de ma motivation et que je trainais cette thèse comme un sacerdoce. Tu m’as remonté le moral, tu m’as poussé, houspillé, engueulé pour que je me bouge et que je me reprenne en main. Reinhard et André n’en sont probablement pas conscients, mais sans toi ce doctorat aurait pu durer 10 ans, 15 ans ou même simplement s’écraser sur le bord de la route. Tu me dis parfois que tu as peur qu’inconsciemment je prolonge mon doctorat pour t’attendre et finir en même temps que toi, mais c’est tout le contraire, sans toi j’y serais encore bien après toi. Avant même Reinhard et André, c’est toi qui as eu la primeur de mes écrits. Sans filtre, sans détour car c’est ton style, tu m’as dit exactement ce que tu en pensais, tu m’as donné tes commentaires. Ce qui m’a poussé à retravailler mon texte avant de leur soumettre pour une première lecture. Merci mon amour pour rester à mes côtés malgré moi et pour toujours me pousser à faire ressortir le meilleur de moi-même.
Il y a les amis qui passent, les amis qui restent et ceux qui font des allers-retours. Je veux remercier tous ces ami(e)s pour leur écoute, leur disponibilité, les discussions, dans les bons comme dans les mauvais moments. J’ai une pensée particulière pour Anabelle Baya qui a mis Virginie sur mon chemin, bien que nous vivions proches nous aurions pu ne jamais nous rencontrer sans elle. Je me souviens avec plaisir de nos longues discussions lorsqu’elle venait poser ses valises chez moi. Pour m’avoir accueilli chez elles à Rimouski malgré mes mauvaises manières et pour m’avoir emporté dans leurs valises afin de m’aérer l’esprit une fois par an en écoutant le souffle des baleines à Tadoussac, je remercie Marie Lionard et Marie Parenteau. Pour avoir fait le voyage jusqu’au pays de l’hiver (deux fois pour Caroline) et continué à partager avec moi les histoires de notre petite France malgré la distance un gros merci à Caroline Naquet et Cécile Petitjean-L’Hopitalier.
Le doctorat, on sait quand ça commence, pas quand ça fini. Et pourtant régulièrement la famille restée en France s’obstine à vous poser cette question tant redoutée : « Alors ta thèse, ça avance bien? C’est quand que tu finis? ». Malgré tout il sont restés à me soutenir et à s’intéresser (même quand ils ne comprenaient pas) tout du long. Biensûr, sur la fin, en voyant que ça se prolongeait, et en ne me voyant que quelques jours tous les deux ans, leurs inquiétudes se sont faites plus pressantes : « Est-ce que tu rentres bientôt en
France? », avec une variation sur le thème de la crise : « Qu’est-ce que tu vas faire après avec un doctorat? ». C’est le genre d’amour et d’attachement qui les
pousse à s’inquiéter pour moi quand moi-même je ne m’inquiète pas (assez?).
Pour finir, j’ai une pensée particulière pour mon père qui nous a quitté il y a bientôt 9 ans, alors que je vivais depuis à peine un an au Québec. Souvent je me demande ce qu’il aurait pensé de tout ça. Si je sais que, comme ma mère, il m’aimait et que le plus important pour lui c’était que je sois heureux même si c’était loin, j’espère aussi qu’il aurait été fier du travail accompli.
xxv
Avant-propos
Cette thèse compile l’ensemble des travaux effectués durant ma recherche doctorale et présente les résultats et observations qui en ont découlés. Cette recherche s’est effectuée sous la co-direction du professeur Reinhard Pienitz du Département de Géographie de la Faculté de Foresterie, de Géographie et de Géomatique à l’Université Laval, et d’André Rochon, professeur en géologie marine à l’Institut des Sciences de la Mer de l’Université du Québec à Rimouski.
La thèse est composée de 5 chapitres dont trois présentés sous forme d’articles scientifiques en anglais, précédés d’un résumé en français. Le premier chapitre est une introduction générale du projet, détaillant les problématiques générales et spécifiques de celui-ci, les spécificités géographiques et climatiques de la région et des sites d’étude, et les outils employés pour répondre aux objectifs. Le dernier chapitre est une conclusion générale de la recherche qui rappelle les principales informations découlant de l’étude et propose des voies exploratoires et des perspectives pour les travaux futures.
Je suis premier auteur sur chacun des articles issus de cette thèse. Reinhard Pienitz est responsable de l’échantillonnage réalisé en 2006 des séquences sédimentaires décrites dans le chapitre 3, je suis co-responsable de la planification du terrain et de l’échantillonnage réalisés en 2009 et décrits dans les chapitres 2 et 4. Je suis également responsable de la plupart des analyses en laboratoire ainsi que du processus d’interprétation des données et de la rédaction. Chapitre 1 : Introduction générale
Chapitre 2 : Richerol, T., Pienitz, R. and Rochon, A., 2012. Modern dinoflagellate
cyst assemblages in surface sediments of Nunatsiavut fjords (Labrador, Canada).
Chapitre 3 : Richerol, T., Pienitz, R. and Rochon, A., 2014. Recent anthropogenic and climatic history of Nunatsiavut fjords (Labrador, Canada). Paleoceanography, DOI : 10.1002/2014PA002624.
Chapitre 4 : Holocene climate history of the Nunatsiavut (north Labrador, Canada) established from pollen and dinoflagellate cyst assemblages covering the past 7,000 years. Ce chapitre sera soumis à la revue The Holocene.
Chapitre 5 : Conclusion générale
Au cours de la thèse, Les résultats et les interprétations des Chapitres 2, 3 et 4 ont été partiellement présentés lors de conférences internationales, sous la forme de :
affiches :
Assemblée Générale Annuelle d’ArcticNet en 2007 (Collingwood, ON) et en 2008 (Arctic Change - Québec, QC).
Conférence Internationale sur les Dinoflagellés Modernes et Fossiles (DINO) en 2008 (DINO8 – GEOTOP, Montréal, QC).
Arctic Frontier 2009 (Trømso, Norvège)
Arctic Workshop en 2009 (Bates College, Lewiston (Maine), USA)
Conférence Internationale en Paléocéanographie (ICP) en 2010 (ICP10 – Université de Californie à San Diego, USA).
présentations orales :
Symposium de Paléolimnologie des Étudiants de l’Ontario et du Québec (PALS) en 2008 (Université Laval, Québec, QC) et en 2010 (Queen’s University, Kingston, ON).
xxvii Conférence Internationale sur les Dinoflagellés Modernes et Fossiles (DINO) en 2011 (DINO9 – University of Liverpool, UK).
1
Chapitre 1. Introduction générale.
1.1. Introduction
Les changements climatiques, et en particulier le réchauffement des conditions climatiques dans les régions de hautes latitudes, sont une des préoccupations majeures de la communauté scientifique, des décideurs politiques et du grand public en général. Afin d’améliorer notre compréhension et notre capacité de prédiction de l’évolution future de notre climat, il a été nécessaire de développer des outils pour identifier et comprendre les rouages des cycles climatiques passés et présents de notre planète. Cette nécessité a émergé du consensus scientifique établi autour des changements climatiques suite aux observations des 20 à 30 dernières années (ACIA, 2005 ; IPCC, 2007 ; AMAP, 2011). Des outils de mesure indirecte de certains paramètres environnementaux (température de l’air ou de l’eau, salinité, pH, couvert de glace, productivité des écosystèmes, etc.), également appelés proxies, ont ainsi été développés. Ces derniers permettent d’estimer l’état du climat passé en fonction de situations analogues connues du climat moderne. Ces proxies paléo-environnementaux représentent des enregistrements passés du climat régional et/ou global.
Mon projet de recherche doctoral s’inscrit dans le cadre du regroupement multidisciplinaire canadien de recherche du Réseau des Centres d’Excellence et du programme ArcticNet (Arctic Network). L’objectif est de comprendre et documenter les changements induits par l’amplification anticipée du réchauffement climatique dans les régions arctiques et subarctiques. En parallèle à ces changements, l’Arctique canadien subit également une « modernisation ». Une partie de l’étude a donc pour objectif de mesurer les impacts de cette modernisation sur les écosystèmes arctiques et subarctiques. Pour répondre à cet objectif, le réseau ArcticNet rassemble des chercheurs de tous les horizons et travaillant sur des problématiques diverses telles que : les changements
climatiques et leurs impacts sur les différents écosystèmes arctiques et subarctiques terrestres et marins, l’impact de la fonte du pergélisol sur les infrastructures humaines, les comportements migratoires des espèces animales nordiques, la santé des populations humaines arctiques en relation avec leur mode de nutrition (chasse et pêche versus aliments transformés venant du sud), la qualité et la quantité d’eau potable disponible, etc. La culture des Inuits canadiens, leur relation particulière à l’environnement arctique et leur approche de l’économie globale sont également pris en compte par les chercheurs d’ArcticNet afin d’intégrer science et savoir Inuit. Afin de tenir compte des différences et des similarités rencontrées sur l’ensemble du territoire, le réseau ArcticNet a découpé cette étendue en quatre régions, chacune possédant un « Integrated Regional Impact Study » (Étude d’Impact Régionale Intégrée) ou IRIS : l’Ouest et le Centre de l’Arctique, L’Est de l’Arctique, la baie d’Hudson, et finalement l’Est subarctique (Figure 1.1). Chaque IRIS produit un rapport pour une des régions déterminées, dans lequel sont compilées toutes les données récoltées par ArcticNet afin de les rendre disponibles pour les organisations gouvernementales et privées mais également pour les communautés Inuits. Mon projet doctoral fait partie du IRIS 4 qui couvre le Nord du Québec appelé Nunavik et le Nord du Labrador appelé Nunatsiavut.
3
Figure 1.1:Carte représentant le découpage des 4 régions IRIS (tiré de Allard and Lemay, 2012)
Mon projet met l’accent sur les écosystèmes côtiers des fjords faisant partie de cette dernière région. Il s’agit plus précisément d’un projet intitulé : « Nunatsiavut Nuluak : comprendre et réagir aux impacts des changements climatiques et de la modernisation sur l’environnement nordique », codirigé par le Gouvernement du Nunatsiavut (Terre-Neuve et Labrador, Canada). Le Nunatsiavut (« notre beau pays »), est la région septentrionale du Labrador se trouvant sous gouvernement Inuit depuis 2005. Les Inuits du Labrador sont dépendants de la mer et de la glace de mer pour la chasse, la pêche mais aussi pour leurs déplacements. Ils sont donc extrêmement préoccupés par l’intégrité écologique du milieu marin du nord du Labrador, plus particulièrement du point de vue des changements climatiques, de l’industrialisation et des contaminants. La sensibilité de la région aux changements climatiques est plus évidente depuis une vingtaine
d’années. Un réchauffement abrupt s’est fait ressentir pour la première fois vers 1993-1995 entraînant de nombreux changements environnementaux tels que la fonte du pergélisol et la formation de mares de thermokarst au Nunavik, et une diminution de 20% de l’étendue des glaciers dans le massif des Monts Torngats entre le Nunavik et le Nunatsiavut de 2005 à 2007 (Brown et al., 2012a). La rareté des stations d’observation et la brève période couverte par les rares enregistrements instrumentales dans ces régions ne permettent pas de documenter plus précisément ce réchauffement. De plus, il existe une réelle différence climatique entre le Nunavik, situé plus à l’intérieur des terres et sous l’influence de la baie d’Hudson, et le Nunatsiavut, situé sur la côte Est et influencé par les apports d’eaux arctiques du courant du Labrador de la mer éponyme (Brown et al., 2012a). En effet, les fluctuations climatiques majeures des 1 000 dernières années, telles que le Petit Âge Glaciaire, sont visibles dans les enregistrements du Nunavik mais imperceptibles dans ceux du Nunatsiavut (Chapitre 4). Environnement Canada a compilé des données sur 55 ans afin d’illustrer la tendance des températures moyennes annuelles pour le pays. De 1948 à 2003, le Centre et l’Ouest du Canada montrent une tendance claire au réchauffement alors que l’Est reste stable avec quelques régions qui montrent une tendance au refroidissement (Figure 1.2).
5
Figure 1.2 : Tendances des températures moyennes annuelles de 1948 à 2003 (Environnement
Canada 2007)
En utilisant le « Canadian Regional Climate Model » (Caya et al., 1995 ; Caya et Laprise, 1999 ; Plummer et al., 2006 ; Music et Caya, 2007) le consortium OURANOS a travaillé sur des scénarios climatiques régionaux pour la zone IRIS 4 (Brown et al., 2012a). Ils ont comparé le climat actuel, pour une période moyenne de 30 ans entre 1971 et 2000, avec le scénario couvrant la période 2041-2070 afin d’estimer l’amplitude des changements projetés. Cette « future » période comprend le cadre temporel de l’année 2050 projeté dans le scénario SRES (« Special Report on Emissions Scenarios ») A2 pour les futures émissions de gaz à effet de serre (Nakicenovic et al., 2000). Le groupe de scénarios A2 définis par le GIEC (Groupe d’experts Intergouvernemental sur l’Évolution du Climat) décrit un monde très hétérogène, caractérisé par l’autosuffisance, le renforcement et la préservation des identités culturelles locales et des valeurs familiales. La population mondiale croît constamment jusqu’à un niveau élevé. Le développement économique et le progrès technologique sont moins rapides
(Villeneuve et Richard, 2005). Les changements projetés sont pour l’essentiel dans la continuité des tendances actuelles observées, soit un réchauffement plus marqué pour le Nunavik et plus lent pour le Nunatsiavut. La plus importante augmentation de la température de l’air et des précipitations est projetée au-dessus du Nord de la péninsule d’Ungava. Globalement les changements climatiques de cette région pour les 30 à 50 prochaines années seront dominés par les influences locales (i.e. grandes masses d’eau comme la baie d’Hudson et la mer du Labrador) plus que par une tendance globale/planétaire (Brown et al., 2012a).
L’un des principaux objectifs de ce projet est d’établir un partenariat intégrant les Inuits et leurs connaissances dans le cadre d’un programme de surveillance à long terme du milieu marin du nord du Labrador. Ce projet est à la base d’un programme qui contribuera à plus long terme aux recherches, à la surveillance et à l’évaluation de l’état de l’ensemble des écosystèmes marins arctiques. Cette étude permettra également de développer les aptitudes des Inuits à comprendre et évaluer les changements occasionnés dans leur milieu dans l’avenir. Plus particulièrement, mon projet doctoral intègre les domaines de recherche de la paléoécologie et de la paléocéanographie et porte sur des séquences sédimentaires océaniques prélevées dans quatre fjords du Nunatsiavut lors de missions océanographiques : Nachvak, Saglek, Okak et Anaktalak. Les
proxies paléo-environnementaux utilisés sont principalement les kystes de
dinoflagellés (= dinokystes), les grains de pollen, les spores ainsi que les diatomées. Le premier objectif de mon projet doctoral est de documenter les assemblages modernes de dinokystes dans les quatre fjords et de faire le lien avec certains paramètres écologiques et physiques caractérisant ces milieux marins côtiers particuliers (température, salinité, durée du couvert de glace, quantité de lumière, nutriments, quantité de diatomées). Il s’agit aussi de déterminer si la récente installation d’une mine dans le fjord d’Anaktalak est enregistrée par ce proxy (Chapitre 2 : Richerol et al., 2012). Le second objectif porte sur des séquences sédimentaires courtes (~25-30 cm) prélevées dans trois fjords (Nachvak, Saglek et Anaktalak) et a pour but la caractérisation des
7 assemblages fossiles de dinokystes, de grains de pollens et de diatomées ainsi que la reconstitution des conditions paléocéanographiques des 150-300 dernières années dans les fjords à partir des dinokystes. Il s’agit essentiellement de déterminer si l’histoire climatique récente du Nunatsiavut, telle qu’enregistrée par ces proxies, correspond à ce qui est déjà connu pour la région et si les récents changements climatiques et l’industrialisation du nord sont perceptibles dans ces enregistrements (Chapitre 3). Le troisième objectif porte sur une séquence sédimentaire longue (~8 m) prélevée dans le fjord de Nachvak et a pour but la reconstitution des conditions paléocéanographiques dans le nord du Nunatsiavut durant l’Holocène (les 10 000 dernières années), à partir des assemblages fossiles de dinokystes et du pollen. Il s’agit de déterminer si la stabilité climatique observée pour la région dans les 100 dernières années a débuté après la dernière grande déglaciation ou est plus récente (Chapitre 4). Les réponses à ces objectifs fourniront de précieuses informations sur l’histoire climatique passée de cette région subarctique peu connue, permettant ainsi de renforcer les modèles d’évolution climatique pour l’Arctique canadien.
1.2. Sites d’étude
1.2.1. Localisation, climat et végétation
Le territoire du Labrador est la partie continentale de la province de « Terre-Neuve et Labrador » sur la côte Atlantique du Canada. Cette province représente environ ~4.2% de la superficie du Canada. Le Labrador couvre une superficie de 294 330 km2, soit un peu plus des 2/3 de la superficie de la province (De Blij,
2005). Ce territoire s’étend entre les 50ème et 60ème parallèles, le long de la mer
éponyme, et repose sur les roches précambriennes du bouclier canadien (Chapitre 2, Figure 2.1).
Une grande partie de la région repose à des altitudes élevées, entre 450 et 750 m au-dessus du niveau de la mer. Au Nord-Est, les chaînes de montagnes des Torngat et des Kaumajet s’élèvent jusqu’à 1600 et 1800 m d’altitude, respectivement (Short and Nichols, 1977). L’Ouest et le centre du Labrador sont caractérisés par de vastes plaines et plateaux de till, couvertes de lacs et de tourbières, tandis que le paysage côtier est dominé par des collines découpées par d’importantes rivières et de majestueux fjords au Nord (Engstrom and Hansen, 1985). Ces fjords subissent à la fois l’influence des masses d’eaux atlantique et arctique mais également des apports d’eau douce, de nutriments et de sédiments des glaciers et des rivières. Les fjords les plus nordiques ont de profonds bassins sédimentaires séparés par des seuils rocheux et sont flanqués de hautes parois abruptes (Nachvak et Saglek). Les fjords dans la partie centrale du Labrador, bien qu’également d’origine glaciaire, sont au contraire peu profonds, de formes irrégulières et avec des parois en pente douce et de grandes zones intertidales (Okak et Anaktalak) (Brown et al., 2012b).
9 Le climat régional est caractérisé par de longs hivers froids et de courts étés chauds et humides. Le régime des températures et de l’humidité est régi par des transports Nord-Sud d’air arctique froid et sec, et d’air atlantique chaud et humide (Hare and Hay, 1974). De plus, le courant marin froid du Labrador, qui s’écoule depuis l’île de Baffin, a une très grande influence sur le climat local (Engstrom and Hansen, 1985), de sorte qu’un important contraste est noté entre le climat de l’intérieur des terres et celui de la côte. En effet, les températures en juillet et août à l’intérieur des terres peuvent atteindre en moyenne 16°C, avec des maximums à 38°C occasionnellement, tandis que le long de la côte les températures moyennes avoisinent les 10°C. Ces conditions s’inversent en hiver quand les températures moyennes à l’intérieur des terres sont relativement plus froides alors qu’elles avoisinent les 3°C le long de la côte (Short and Nichols, 1977; De Blij, 2005). Les précipitations moyennes annuelles au Labrador varient entre 750 mm au nord et 960 mm au sud. Les précipitations sont également distribuées au cours de l’année, avec des quantités légèrement supérieures en été et au début de l’automne. Les chutes de neige sont relativement importantes avec des quantités annuelles comprises entre 390 et 480 cm (Ullah et al., 1992; De Blij, 2005).
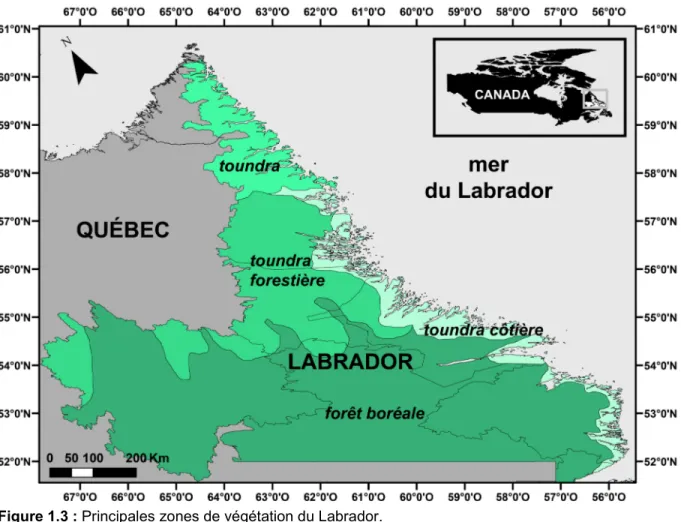
La végétation du Labrador peut être divisée en quatre zones biogéographiques (Hare, 1959; Lamb, 1984; Engstrom and Hansen, 1985; Ecoregions Working Group, 1989; Ullah et al., 1992; Richard, 1995; Williams et al., 2000; Jessen et al., 2011) : la toundra, la toundra forestière, la toundra côtière et la forêt boréale (Figure 1.3).
Figure 1.3 : Principales zones de végétation du Labrador.
Cette subdivision végétale suit un gradient latitudinal Nord-Sud. Au Labrador, la toundra s’étend plus au Sud par rapport aux régions au centre et à l’Ouest du Canada. L’intégralité de la frange côtière, qui est exposée à un climat océanique rude, ne supporte également qu’une végétation de toundra (Engstrom and Hansen, 1985). La toundra se retrouve essentiellement au Nord du 58ème
parallèle, bien que des extensions australes existent, comme sur le plateau de la région de Nain. Elle est principalement composée de Carex et d’arbustes dans les habitats humides, et d’un couvert de lichens dans les zones élevées exposées. Cyperaceae, Ericaceae, Salix spp. (saules)., Betula spp. (bouleaux), Alnus sp. (aulnes), et des herbacées diverses sont les éléments les plus importants de la végétation. La présence d’une végétation caractéristique d’un climat arctique au Sud du Labrador est principalement due au refroidissement provoqué par le
11 courant du Labrador, mais aussi à la persistance et à la distribution de la banquise hivernale (Short and Nichols, 1977).
La toundra forestière est caractérisée par un faible habitat forestier boréal, composé d’épinettes noires (Picea mariana) et blanches (Picea glauca), de mélèze d’Amérique (Larix laricina) et de sapin baumier (Abies balsamea) dans les vallées fluviales, et d’une toundra dans les régions entourées de fleuves (Short and Nichols, 1977).
Enfin, la forêt boréale est composée de peuplements continus de forêts denses avec un couvert végétal au sol riche en mousses. L’épinette noire est l’espèce dominante, accompagnée de quelques peupliers baumiers (Populus
balsamifera) et faux-tremble (Populus tremuloides), de bouleau blanc (Betula papyrifera), de sorbier des montagnes (Sorbus decora) et d’aulnes (Ullah et al.,
1992; Richard, 1995; Williams et al., 2000; Jessen et al., 2011).
Le Nunatsiavut est la région du Labrador située au Nord du 56ème parallèle. Du Nord au Sud, les quatre fjords explorés dans ce projet sont Nachvak, Saglek, Okak et Anaktalak (Chapitre 2, Figures 2.1 et 2.2). La stratification des eaux de surface dans les fjords est contrôlée par les apports d’eaux douces provenant des rivières et de la fonte du couvert de glace, soit une circulation de type estuarienne. Durant l’été, une forte pycnocline s’installe avec des eaux douces de surface recouvrant des eaux intermédiaires plus salées. Cette pycnocline est plus prononcée dans les fjords les plus septentrionaux. Au contraire, durant l’automne, la colonne d’eau est beaucoup plus homogène et non stratifiée (Richerol et al., 2012). Les fjords de Nachvak et de Saglek ainsi que la baie d’Anaktalak enregistrent les plus faibles salinités durant l’été (respectivement 29, 30,4 et 27,9). Contrairement à la plupart des fjords de la côte Ouest canadienne dont les eaux sont anoxiques (Kumar and Patterson, 2002; Hay et al., 2003; Patterson et al., 2011), les eaux des fjords du Nunatsiavut sont riches en oxygène, démontrant une très bonne circulation océanique et un renouvellement continuel des masses d'eau de fond (Richerol et al., 2012).
Dans cette étude, le fjord de Nachvak est le plus au Nord, localisé dans le Parc National des Monts Torngat. Il représente l’écosystème le plus isolé et le mieux préservé de l’influence des activités humaines. Dans le cadre de ce projet, Nachvak sert de site de référence afin de pouvoir estimer la variabilité climatique et environnementale naturelle des fjords du Nunatsiavut (Chapitre 2, Figure 2.2A). Le fjord de Nachvak est une dépression d’origine glaciaire de 45 km de longueur qui coupe à travers les Monts Torngats, présentant des parois de 1 km de hauteur et des bassins pouvant atteindre 200 m de profondeur. Le fjord est d'une largeur de 2 à 4 km et s’élargit vers l’Est et la baie de Nachvak qui s’ouvre sur la mer du Labrador (Bell and Josenhans, 1997). De la tête du fjord jusqu’à son embouchure, la profondeur des bassins augmente avec une profondeur de la colonne d’eau allant de 90 m à 210 m. Les bassins sont séparés par des seuils situés entre 10 et 180 m sous le niveau de la mer et composés soit de matériel rocheux soit de dépôts glaciaires (Bell and Josenhans, 1997). Le fjord de Nachvak reçoit l’essentiel de ses apports sédimentaires de la rivière Ivitak alimentée par un glacier et drainant les parties Sud et centrale du bassin versant du fjord (Kahlmeyer, 2009).
La baie de Saglek, à l’entrée du fjord éponyme, a été le site d’une station radar militaire faisant partie de la ligne de détection lointaine avancée (DEW-Line : Distant Early Warning) durant la guerre froide (Chapitre 2, Figure 2.2B). Après l’abandon du site en 1971, une mauvaise gestion des déchets provenant des huiles et lubrifiants utilisés pour le bon fonctionnement de la station a entrainé une contamination des sols aux biphényles polychlorés (BPC). Autour de la baie de Saglek, des concentrations élevées atteignant 250 µg de BPC par gramme de sol ont été mesurées (Pier et al., 2003). Ces sols contaminés ont à leur tour été source de contamination pour les lacs des alentours ainsi que les eaux et les sédiments de la baie. Plusieurs études portant sur des microalgues d’eau douce (Paterson et al., 2003) et différentes espèces animales marines (Kuzyk et al., 2003, 2005a) ont apporté la preuve d’une bioaccumulation et d’une bioamplification des contaminants à travers la chaîne alimentaire locale (Kuzyk et al., 2005b). Le fjord de Saglek est un vaste système de baies et de fjords reliés
13 entre eux et long de 65 km (Fitzhugh, 1980). Le fjord s’ouvre à l’Est dans la mer du Labrador par la baie de Saglek, large de plus de 14 km et parsemée de petites et grosses îles. De manière générale, les parois du fjord sont abruptes et hautes de plus de 800 m à partir du niveau de la mer. Les différents bassins du fjord sont de plus en plus profonds en s’approchant de l’embouchure, s’étendant de 80 à 256 m de profondeur sous le niveau de la mer. Des seuils situés entre 45 et 96 m sous le niveau de la mer séparent ces différents bassins. Le fjord de Saglek reçoit l’essentiel de ses apports sédimentaires de la rivière Nachvak, alimentée par la fonte de la neige et drainant la partie Nord et centrale du bassin versant du fjord (Kahlmeyer, 2009).
Le fjord d’Okak était le site d’une communauté Moravienne à partir de 1775. La pandémie de grippe espagnole de 1918 a dévasté cette communauté laissant le fjord inoccupé depuis (Rompkey, 2003), à l’exception des expéditions occasionnelles de chasse et de pêche des Inuits de la communauté de Nain située plus au Sud (Chapitre 2, Figure 2.2C). La baie d’Okak forme une crique de forme irrégulière de 50 km de longueur, avec des entrées Nord et Sud de part et d’autre de l’île d’Okak. Le bassin versant environnant est vallonné et de faible altitude avec quelques sommets au-dessus de 400 m. La profondeur moyenne est comprise entre 40 et 80 m, avec des bassins à fonds plats séparés par des seuils au relief limité. La tête de la baie est relativement peu profonde (45-50 m) avec deux affluents majeurs: les rivières Saputit et Nord. Le bassin le plus profond (200 m) est situé du côté Nord-Est de l’île d’Okak le long de l’entrée Nord de la baie. L’entrée sud de la baie est étroite et peu profonde (Brown et al., 2012b).
Mon site d’étude le plus au Sud se situe dans le fjord d’Anaktalak. Ce fjord est régulièrement utilisé par les Inuits pour la chasse, la pêche et leurs déplacements. Entre 1997 et 2002, une mine de nickel s’est établie dans la baie de Voisey (Hulett and Dwyer, 2003; Noble and Bronson, 2005). Depuis le début des activités de la mine de nickel Vale Inco, le fjord et l’environnement marin ont reçu d’importantes quantités d’effluents de cette mine. Les communautés du
Nunatsiavut ont exprimé leurs inquiétudes quant à la nature de ces rejets et l’impact des activités de la mine et du transport maritime sur l’intégrité écologique du fjord d’Anaktalak. Un des objectifs du projet est de développer des indicateurs appropriés pour la surveillance à long terme des conditions écologiques et environnementales dans l’écosystème de ce fjord (Chapitre 2, Figure 2.2D). La baie d’Anaktalak, aussi appelée baie de Voisey, est un fjord long et étroit de 65 km et d'une largeur de 2 km en majeure partie. Les parois du fjord sont basses et discontinues, composées de nombreuses îles de tailles variables. L’essentiel de la baie forme un large bassin profond de 100-200 m qui remonte au niveau du seuil à 85 m dans la partie externe de la baie (Kahlmeyer, 2009).
1.2.2. La déglaciation à l’Holocène au Labrador
Lors de la dernière glaciation du Wisconsinien (de 85 000 à 10 000 BP (BP = Before Present)), le Labrador était complètement recouvert par l’Inlandsis Laurentidien (glacier continental). Les seules exceptions étaient quelques nunataks (des hauts pics libres de glace car s’élevant au-dessus du dôme de glace) dans les monts Torngat. Un réchauffement s’est produit il y a environ 11 000 BP (le Maximum Thermique Holocène) libérant des glaces le sud du Labrador (Dyke and Prest, 1987; Dyke et al., 2002; Dyke, 2004). Cependant, dépendamment du patron de déglaciation de l’Inlandsis Laurentidien et de l’influence des cycles solaires à travers l’Amérique du Nord et le Canada, le rythme et l’importance du réchauffement ont varié substantiellement entre l’Ouest et l’Est du Canada (Renssen et al., 2009). Entre 11 000 et 6 000 BP, le retrait de l’Inlandsis a libéré le Nord du Labrador tandis que des restes de glace ont persisté sur le haut plateau central de la région du Labrador-Ungava jusqu’à environ 6 000 BP (Lamb, 1980; Engstrom and Hansen, 1985; Richard, 1995). Une phase rapide de déglaciation du centre de l’Inlandsis s’est produite entre 9 000 et 8 400 BP au-dessus de la baie d’Hudson, suivant la vidange catastrophique du lac proglaciaire Agassiz via le Détroit d’Hudson vers 8 200 BP (Barber et al., 1999; Dyke, 2004; Carlson et al., 2007; Lajeunesse et St-Onge, 2008). Les
15 enregistrements paléoclimatiques montrent un décalage du Maximum Thermique Holocène vers 7 000-6 000 BP au centre et au Nord-Est de l’Amérique du Nord et dans le Sud du Groenland (Kaufman et al., 2004; Kaplan and Wolfe, 2006; Carlson et al., 2007). À Terre-Neuve (Sud du Labrador), la déglaciation a débuté autour de 10 000 BP. D’après les enregistrements palynologiques, la végétation a colonisé l’île après 9 300 BP (Macpherson, 1982; Ritchie, 1987). Au Nord, vers 7 000 BP, les conditions climatiques étaient plus froides au niveau des régions directement influencées par la présence de l’Inlandsis (Renssen et al., 2009). Des études ont suggéré la disparition de l’Inlandsis autour de 7 000 BP, avec cependant la persistance de plus petits glaciers isolés dans le Nord du Québec (région de Schefferville) et du Labrador jusqu’à 6 000 BP (Richard, 1995; Peltier, 2004; Carlson et al., 2008; Renssen et al., 2009).
1.3. Les kystes de dinoflagellés (= dinokystes)
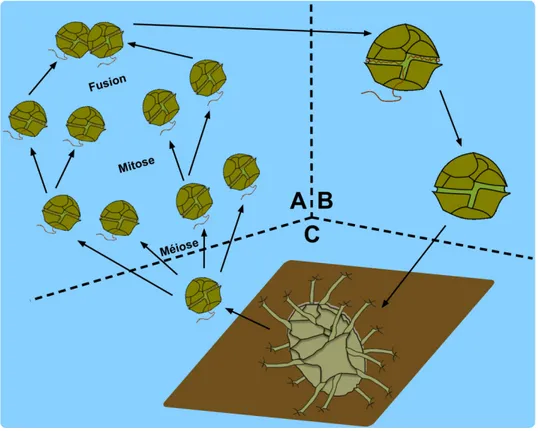
Les dinoflagellés sont des organismes unicellulaires microscopiques biflagellés faisant partie du phytoplancton marin (Taylor et al., 2008). Ils peuvent occasionnellement former des floraisons massives appelées marées rouges. Certaines espèces peuvent être toxiques. Le cycle de vie de certaines espèces de dinoflagellés comporte une phase sexuée après laquelle un kyste résistant est formé (Figure 1.4).
Figure 1.4 : Schéma du cycle de vie des dinoflagellés. A) phase haploïde durant laquelle les
individus (1 dinoflagellé = 1 cellule) se divisent via la mitose. Il s’agit du stade de vie végétatif qui produit les événements de floraison ou bloom. À la suite d’un signal interne et/ou externe, des gamètes se forment et fusionneront pour produire un zygote diploïde; B) phase planozygote (zygote mobile pélagique) durant laquelle la cellule diploïde perdra éventuellement sa mobilité et développera un kyste qui sera éventuellement libéré dans la colonne d’eau puis sédimentera; C) phase hypnozygote durant laquelle le kyste subira une période de dormance obligatoire contrôlée par une horloge interne qui déclenchera le moment de la libération du dinoflagellé ou exkystement (adapté de Rochon, 2009).
17 Ce dinokyste est composé de dinosporine, un polymère très résistant, similaire à la sporopollénine impliquée dans la construction de la paroi des grains de pollen et des spores. Le dinokyste est ainsi préservé dans les sédiments jusqu’à la venue des conditions nécessaires à la floraison (Rochon, 2009). La forme et l’ornementation du kyste sont caractéristiques pour chaque espèce et permettent leur identification.
Les dinokystes ont été utilisés dans de nombreuses études en tant que
proxies paléocéanographiques de certains paramètres environnementaux
caractéristiques du développement de chaque espèce (salinité et température des eaux de surface, durée du couvert de glace, productivité). La méthode de reconstitution quantitative employée utilise des fonctions de transfert qui comparent les assemblages fossiles des séquences sédimentaires marines aux assemblages modernes de dinokystes des sédiments de surface afin d’identifier les meilleurs analogues modernes et les conditions environnementales qui leurs sont liées. (Rochon et al., 1999; Devillers and de Vernal, 2000; Boessenkool et al., 2001; de Vernal et al., 2001, 2005, 2006; Grøsfjeld and Harland, 2001; Kunz-Pirrung, 2001; Mudie and Rochon, 2001; Radi et al., 2001; Voronina et al., 2001; Radi and de Vernal, 2008; Richerol et al., 2008a,b; Bonnet et al., 2012).
Les dinokystes ont déjà été employés dans des études précédentes portant sur des fjords de régions arctiques et subarctiques. Dans les fjords norvégiens, la dynamique de la formation, de la sédimentation et la préservation des dinokystes a été étudiée à Trondheimsfjord (Dale, 1976), tandis que les dinokystes ont été employés comme proxy pour la pollution et le climat passé à Nordåsvannet et Grimstadfjord (Thorsen et Dale, 1997) et comme proxy pour l’eutrophisation d’origine humaine à Oslofjord (Dale et al., 1999). Les dinokystes ont également été employés dans une étude multi-proxies sur le climat Holocène dans le fjord subarctique de Malangen (Hald et al., 2003). Dans le fjord de Gullmar (Suède), la relation entre l'abondance des dinoflagellés planctoniques, l'abondance de dinokystes récupérés dans des pièges à sédiments et les facteurs
environnementaux a également été étudiée (Godhe et al., 2001; Harland et al., 2006). Dans le fjord d’Igaliku (Sud du Groenland), les dinokystes ont été employés pour documenter les palynofaciès de la fin de l'Holocène en relation avec la paléohydrographie du fjord (Roncaglia et Kuijpers, 2004). Les assemblages de dinokystes des fjords du Svalbard ont été documentés en relation avec les conditions océanographiques modernes (Kongsfjorden, Krossfjorden, Sassenfjorden, Van Mijenfjorden et Storfjorden) (Grøsfjeld et al., 2009) et l'influence de la circulation atlantique (Kongsfjorden et Rijpfjorden) (Howe et al., 2010). Sur la côte Ouest du Canada, le fjord anoxique d’Effingham (Île de Vancouver, Colombie-Britannique.) a été étudié afin de caractériser les assemblages modernes de dinokystes (Kumar et Patterson, 2002) et les conditions paléocéanographiques (Patterson et al., 2011).
La glace de mer est un paramètre complexe, difficile à reconstituer à partir d’observations indirectes. Alors que les sciences du climat considèrent la glace de mer comme un paramètre purement physique, les géosciences reconstituent les conditions passées de glace de mer en assumant qu’elle joue un rôle central dans la biogéochimie de l’eau de mer et donc dans la production primaire et la structure trophique des populations planctoniques (Meier et al., 2011). La plupart des
proxies pour la glace de mer employés dans les reconstitutions proviennent de
restes d’organismes vivants récupérés dans les sédiments marins (i.e. diatomées, foraminifères, dinokystes, ostracodes). Comme les zones de productivité dans les environnements de glace de mer se retrouvent essentiellement proche de la limite de la glace au printemps et à l’été, la plupart des proxies sont reliés à la saisonnalité de la glace de mer (de Vernal et al., 2013). Les dinokystes en particulier fournissent de l’information sur l’étendue annuelle du couvert de glace de mer pour l’hémisphère Nord (de Vernal et al., 2008). Cependant, leur relation avec le couvert de glace est souvent indirecte, dépendante plus directement de la quantité de lumière et/ou de nourriture disponibles en fonction du couvert de glace (Richerol et al., 2012). De plus, l’hétérogénéité taxonomique des populations dans l’espace peut être liée à l’endémisme ou au développement de différents
19 génotypes pour différentes affinités écologiques. Ainsi, les dinokystes sont essentiellement utiles à une échelle régionale pour reconstituer le couvert de glace et doivent être comparés à d’autres indicateurs pour une interprétation plus complète des conditions paléoenvironnementales (de Vernal et al., 2013).
1.4. Reconstitutions climatiques au Labrador et dans les régions
avoisinantes
À l’heure actuelle, il existe peu de reconstitutions paléoclimatiques qui portent uniquement sur la région du Labrador. La plupart englobent de plus grandes régions dont le Labrador ne représente qu’une petite partie (par exemple, le Nord-Est du Canada, la région boréale du Canada, l’Arctique et le Subarctique canadien ou la mer du Labrador). Une étude de Fallu et al. (2002) utilise comme
proxy les assemblages modernes de diatomées provenant des sédiments de
surface de 64 lacs du Québec-Labrador. Les analyses statistiques de ces assemblages ont montré une nette corrélation entre la répartition des espèces et la couleur de l’eau et l’alcalinité. Ces assemblages de diatomées permettent de démontrer le potentiel de cette base de données pour déceler des changements plus subtils du couvert végétal du bassin versant des lacs. Les reconstitutions climatiques réalisées sur une séquence sédimentaire lacustre du Nord du Québec, en utilisant cette base de donnée de diatomées, ont révélé une tendance régionale au refroidissement sur les derniers 1 500 ans et l’absence de signe du réchauffement récent (Fallu et al., 2005). En règle générale, les études portant sur une reconstitution climatique du Labrador et du Nord du Québec à partir de diatomées montre une stabilité climatique au cours des 200 dernières années (Laing et al., 2002 ; Paterson et al., 2003 ; Smol et al., 2005), en décalage avec le reste de l’Arctique qui montre un réchauffement plus ou moins rapide selon les régions (ACIA, 2005).
Les reconstitutions climatiques pour le Labrador, basées sur le pollen, portent sur les ~10 000 dernières années, selon des échelles de temps millénaires (orbitales) afin de mieux appréhender le réchauffement climatique actuel (Viau et Gajewski, 2009). De nombreux sites au Labrador enregistrent une longue période de végétation de type toundra après la déglaciation (Short et Nichols, 1977 ; Lamb, 1980) ce qui conduit à une reconstitution de températures froides pour le début de
21 l’Holocène. La transition d’une toundra herbacée vers une toundra arbustive dans le Nord du Labrador (Short et Nichols, 1977) et la diminution du sapin et de l’épinette blanche favorisant l’épinette noire dans le Sud du Labrador (Engstrom et Hansen, 1985) vers 6 500 BP, témoignent d’une augmentation rapide des températures hivernales. Un maximum des températures estivales entre 4 000 et 2 500 BP correspond au maximum d’abondance relative de l’épinette et de l’accumulation pollinique dans le Sud-Est et le centre du Labrador (Short et Nichols, 1977 ; Lamb, 1980). Cette reconstitution faite à partir des assemblages polliniques n’a cependant pas la résolution suffisante pour faire ressortir l’impact du réchauffement climatique actuel sur la végétation locale (Viau et Gajewski, 2009).
Les reconstitutions reposant sur les assemblages de dinokystes sont principalement concentrées dans la mer du Labrador, le Nord-Ouest de l’océan Atlantique ou Nord-Est de l’Arctique canadien et la baie de Baffin (Hillaire-Marcel et al., 1994 ; Levac et de Vernal, 1997 ; Sawada et al., 1999 ; de Vernal et al., 2002 ; de Vernal et Hillaire-Marcel, 2006 ; Radi et de Vernal, 2008a). Il existe une base de données des assemblages modernes de dinokystes pour l’hémisphère Nord (n=1 171), mais les sites spécifiques aux écosystèmes du Labrador en sont absents en-dehors de quelques-uns dans la mer du Labrador (Radi et de Vernal, 2008b). Les reconstitutions climatiques réalisées à partir des dinokystes couvrent le domaine marin et sont complémentées par des reconstitutions réalisées à partir du pollen de la végétation terrestre afin d’établir une corrélation entre la palynostratigraphie continentale et la palynostratigraphie océanique (Levac et de Vernal, 1997 ; Sawada et al., 1999).
Des analyses statistiques ont montré, qu’au niveau régional, il existe une corrélation entre les deux types de reconstitutions car les deux proxies répondent au même signal climatique. L’étude palynologique d’une séquence sédimentaire prélevée le long de la côte du Labrador, a permis de reconstituer les conditions marines de surface grâce aux dinokystes et de les corréler avec l’histoire de la végétation des terres adjacentes sur la base des assemblages polliniques fossiles