Utilisation de l'espace par les grands herbivores dans
un environnement hétérogène et dynamique :
méthodologie et applications
Thèse
Marie-Caroline Prima
Doctorat en biologie
Philosophiæ doctor (Ph. D.)
Québec, Canada
Utilisation de l’espace par les grands herbivores
dans un environnement hétérogène et dynamique :
méthodologie et applications
Thèse
Doctorat en biologie
Philosophiae Doctor (Ph.D.)
Marie-Caroline Prima
Sous la direction de :
Daniel Fortin, directeur de recherche
Thierry Duchesne, codirecteur de recherche
Résumé
L’objectif général de cette thèse est de développer des modèles mécanistes de l’utilisation de l’espace qui sont basés sur les déplacements des animaux, afin de comprendre et d’anticiper la répartition spatiale des populations animales dans des environnements hétérogènes et dynamiques. J’utilise et je développe des méthodologies qui intègrent autant la modélisation mathématique de la dynamique spatio-temporelle des déplacements que des analyses statistiques de données simulées et empiriques de déplacement. Dans mon premier chapitre, j’effectue une série de simulations afin de clarifier combien de grappes sont nécessaires lors d’une estimation par équations d’estimation généralisées, pour correctement tenir compte de la corrélation dans les données de déplacement et obtenir des inférences robustes sur la sélection de l’habitat. Mes simulations révèlent que 30 individus indépendants, chacun étant assigné à une grappe, suffisent pour éviter la mesure biaisée de l’incertitude sur la sélection de l’habitat lors des déplacements dans un environnement hétérogène. Dans le cas où moins de 30 individus sont disponibles, il est possible d’augmenter le nombre de grappes en divisant les données des individus, cependant il faut s’assurer de la présence d’une autocorrélation temporelle et d’une faible hétérogénéité interindividuelle dans les données. Dans mon deuxième chapitre, je développe un modèle statistique de déplacement permettant d’identifier différentes phases comportementales par lesquelles passent les individus (p. ex., alimentation, déplacements entre les parcelles de ressource) et de révéler la sélection de l’habitat spécifique à chaque phase, pour l’ensemble de la population et à partir de données collectées irrégulièrement. L’analyse de données simulées et empiriques de déplacement de trois grands herbivores dont le bison des prairies (Bison bison bison), le cerf à queue noire (Odocoileus hemionus) et le zèbre des plaines (Equus quagga) démontrent la robustesse et la bonne capacité de prévision du modèle. Cet outil statistique est également flexible puisque j’évalue différents processus écologiques à partir de ces données tels que l’alimentation, la migration ou encore les réponses comportementales face à un prédateur. De plus, je montre la nécessité de tenir compte des phases comportementales pour correctement caractériser la sélection de l’habitat lors des déplacements des animaux. Le développement mathématique que j’ai effectué dans mon troisième chapitre permet de coupler les déplacements des individus au sein d’un réseau de parcelles de ressources et le temps de résidence dans les parcelles afin de prévoir, de façon
mécaniste, la répartition spatiale d’une population dans un environnement hétérogène. De plus, j’illustre une méthodologie pour identifier et prévoir le réseau théorique le plus représentatif de l’espèce étudiée. Je démontre à partir de l’application du modèle aux données de bisons des prairies, que la topologie du réseau théorique est cruciale pour correctement anticiper l’utilisation de l’espace d’une population, ainsi que pour implémenter des plans de gestion ou de conservation les plus réalistes possibles. Dans mon chapitre 4, je teste empiriquement la robustesse d’un réseau de parcelles de ressources lorsque celui-ci est perturbé par une fragmentation anthropique du paysage. Les résultats révèlent que les caribous des bois (Rangifer tarandus caribou) reconnectent certaines parcelles favorisant ainsi la robustesse du réseau. Cependant, les prévisions de la répartition spatiale des individus obtenues en utilisant le modèle mécaniste développé dans le chapitre 3 démontrent que, malgré la reconnexion, l’utilisation des parcelles de ressources par les caribous change suite à la perturbation. De plus, ce changement est plus soutenu lorsque ce sont les parcelles les plus connectées (c.-à-d., les pôles) qui sont fragmentées. Ma thèse apporte une contribution méthodologique pour mieux tenir compte de la corrélation dans les données de déplacement et intégrer les phases comportementales lors de l’analyse de la sélection de l’habitat dans des paysages hétérogènes. Mon travail permet aussi de faire le lien entre la théorie des réseaux et l’utilisation de l’espace pour prévoir d’une façon mécaniste la répartition spatiale des populations animales dans des environnements hétérogènes et dynamiques. Mon doctorat donne également lieu à une évaluation du contexte dans lequel la théorie des réseaux peut s’appliquer à l’écologie spatiale. Finalement, ma thèse vient améliorer notre compréhension mécaniste des déplacements de quatre espèces de grands herbivores.
Abstract
In my thesis, I develop mechanistic models of space use based on animal movement, to understand and to predict population distribution in heterogeneous and dynamic landscapes. Used and developed methodologies couple mathematical modelling of the spatio-temporal dynamics of animal movement together with statistical analysis of simulated and empirical movement datasets. In my first chapter, I proceed in a series of simulations to clarify how many clusters are needed when using generalized estimating equations to correctly account for the correlation in movement data and to obtain robust inference on habitat selection. My simulations reveal that 30 independent individuals, each assigned to a cluster, are sufficient to avoid biased evaluation of the uncertainty on habitat selection along movement in heterogeneous environments. When less than 30 individuals are available, destructive sampling can be used but solely when temporal correlation is present and inter-individual heterogeneity is low in the data. In my second chapter, I develop a statistical movement model that allows to identify successive behavioral phases (e.g., foraging phase, inter-patch movement) together with behavior-specific habitat selection parameters, over the whole population and using temporally irregular data. Analysis of simulated and empirical movement data from three large herbivores including plains bison (Bison bison bison), mule deer (Odocoileus hemionus) and plains zebra (Equus quagga) show the robustness and the high predictive capacity of the model. This statistical tool is also flexible since I assess multiple ecological processes from those datasets such as foraging behavior, migratory behavior or prey-predator interactions. In addition, I show how accounting for behavioral phases in habitat selection analysis is crucial to correctly characterize habitat selection along animal movement. In my third chapter, I develop a mathematical framework to couple movement of individuals among a network of resource patches with residency time in patches to mechanistically predict space use in heterogeneous landscapes. In addition, I illustrate a methodology to identify and predict the most representative theoretical network for the target species. I show from model application on data of plains bison that the theoretical network topology is crucial to correctly infer population space use and implement realistic management and conservation planning. In my chapter 4, I empirically assess the robustness of a network of resource patches following landscape fragmentation from anthropogenic source. The analysis shows that woodland caribou (Rangifer tarandus caribou) reconnect
some patches, thus causing robustness in their spatial networks. However, predictions on space use from the mechanistic model developed in chapter 3 reveal that, despite the rewiring, patch use change following the fragmentation. Moreover, this change is stronger when the most connected patches (i.e., the hubs) are impacted. My thesis provides a methodological contribution to better account for correlation in movement data and integrate behavioral phases in habitat selection analysis in heterogeneous landscapes. Besides, my work links network theory and space use to mechanistically predict population distribution in heterogeneous and dynamic environments. My research also assesses the context in which network theory can be applied to spatial ecology. Finally, my thesis improves our mechanistic understanding of animal movement in four species of large herbivores.
Table des matières
Résumé ... ii
Abstract ... iv
Table des matières ... vi
Liste des figures ... ix
Liste des tableaux ... xii
Remerciements ... xiii
Avant-propos ... xv
Introduction générale ... 1
1. La répartition spatiale des populations animales : un thème majeur en écologie ... 1
2. Anticiper la répartition spatiale d’une population dans un environnement dynamique .... 3
3. L’approche mécaniste de la répartition spatiale des populations : une approche basée sur les déplacements ... 5
3.1 Les différentes composantes mécanistes du déplacement ... 5
3.2 Les facteurs influençant les déplacements ... 6
3.3 Identification des facteurs influençant les déplacements ... 6
4. Les données de déplacement: opportunités et défis associés à l’essor des technologies GPS ... 7
4.1 La corrélation dans les données de déplacement ... 7
4.2 L’identification des phases comportementales à partir des données de déplacement ... 9
5. Les déplacements et la théorie des graphes ... 11
5.1 Qu’est-ce que la théorie des graphes ? ... 11
5.2 Prévision de la connectivité du paysage à partir de la théorie des graphes ... 12
5.3 Relier la théorie des graphes à la répartition spatiale des populations ... 14
5.4 La théorie des graphes dans un contexte de perturbations ... 15
6. Objectif général et organisation de la thèse ... 16
7. Modèles d’études : les grands herbivores ... 19
Chapitre 1. Robust inference from conditional logistic regression applied to movement and habitat selection analysis ... 22
1.1 Résumé ... 23
1.2 Abstract ... 24
1.3 Introduction ... 25
1.4 Materials and methods ... 27
1.4.1 Notation and model ... 27
1.4.2 Robust estimates of variance using GEE ... 28
1.4.3 Dataset simulation ... 29
1.4.5 Statistical analysis ... 32
1.5 Results ... 33
1.5.1 Naive estimate of variance ... 33
1.5.2 Inter-individual heterogeneity ... 33
1.5.3 Temporal autocorrelation with or without inter-individual heterogeneity ... 34
1.5.4 Coefficient estimates ... 34
1.6 Discussion ... 35
Chapitre 2. Determinants of multi-mode movement decisions across widely ranging behavioral processes ... 41
2.1 Résumé ... 42
2.2 Abstract ... 43
2.3 Introduction ... 44
2.4 Materials and methods ... 48
2.4.1 HMM-SSF overview ... 48
2.4.2 Dealing with irregular time intervals and missing data ... 49
2.4.3 Population level estimates ... 49
2.4.4 Evaluation of model predictive capability ... 50
2.4.5 Simulation study ... 51
2.4.5 Case studies ... 54
2.5 Results ... 60
2.5.1 Testing the robustness of the HMM-SSF using simulations ... 60
2.5.2 Case studies ... 60
2.6 Discussion ... 62
Chapitre 3. Combining network theory and reaction-advection-diffusion modelling for predicting animal distribution in dynamic environments ... 75
3.1 Résumé ... 76
3.2 Abstract ... 78
3.3 Introduction ... 79
3.4 Methods ... 81
3.4.1 Development of reaction-advection-diffusion model ... 81
3.4.2 Identification of spatial network topology ... 83
3.4.3 Predicting links in complex networks ... 84
3.4.4 Case study ... 84
3.5 Results ... 90
3.5.1 Properties of the empirical network ... 90
3.5.2 Network based on complex topology ... 90
3.5.3 Effect of habitat attributes on residency time and patch choice ... 91
3.5.4 Effect of network structure on space use pattern ... 91
3.5.5 Effect of an intervention on space use pattern ... 91
Chapitre 4. A landscape experiment of network robustness and space use
reorganization following habitat fragmentation ... 99
4.1 Résumé ... 100
4.2 Abstract ... 102
4.3 Introduction ... 103
4.4 Materials & methods ... 105
4.4.1 Study area ... 105
4.4.2 Spatial data ... 106
4.4.3 Telemetry data ... 106
4.4.4 Design of the experiment ... 106
4.4.5 Spatial network and space-use predictions ... 108
4.4.6 Test of spatial network and space use predictions ... 110
4.4.7 Evaluation of spatial network and space-use reorganisation depending on the type of disturbed node ... 111
4.5 Results ... 113
4.6 Discussion ... 114
Conclusion générale ... 122
1. Contribution dans le développement de modèles statistiques et mathématiques en écologie spatiale ... 122
2. Tenir compte de la corrélation pour analyser la sélection de l’habitat lors des déplacements ... 125
3. Acquérir une compréhension mécaniste des déplacements ... 126
3.1 Compréhension de la dynamique spatiale des déplacements ... 126
3.2 Compréhension de la dynamique temporelle des déplacements ... 128
4. Application de la théorie des réseaux à des systèmes réels ... 130
5. Perspectives futures ... 131
Bibliographie ... 135
Annexe A. Supporting material of Chapitre 1. Robust inference from conditional logistic regression applied to movement and habitat selection analysis ... 157
Annexe B. Supporting material of Chapitre 2. Determinants of multi-mode movement decisions across widely ranging behavioral processes ... 196
Annexe C. Supporting material of Chapitre 3. Combining network theory and reaction-advection-diffusion modelling for predicting animal distribution in dynamic environments ... 203
Annexe D. Supporting material of Chapitre 4. A landscape experiment of network robustness and space use reorganization following habitat fragmentation ... 217
Liste des figures
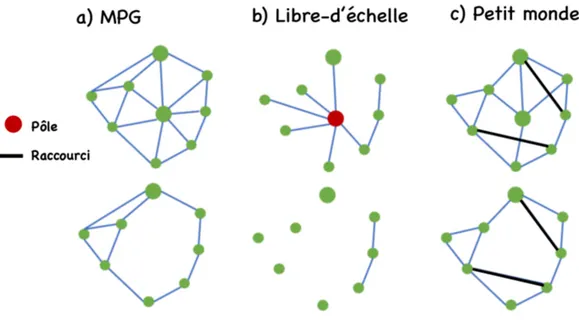
Figure 1. Illustration de topologies d’un réseau simple, le graphe planaire minimum (MPG) et de deux réseaux complexes, le graphe libre-d’échelle et le graphe du petit monde, ainsi que de leur changement potentiel de connectivité suite au retrait complet d’un nœud. Dans un réseau spatial, chaque nœud représente une parcelle de ressources pour l’espèce étudiée et les liens représentent des déplacements inter-parcelles observés ou potentiels. Un pôle (nœud rouge) est un nœud très connecté dans un réseau libre-d’échelle, comparativement aux autres nœuds du réseau qui sont, quant à eux, peu connectés. Un raccourci (lien bleu foncé) est un lien direct entre deux parcelles éloignées, qui croise d’autres liens et qui permet de traverser le réseau en passant par un faible nombre de liens intermédiaires. Les raccourcis sont inhérents au réseau du petit monde. ... 14 Figure 1.1. Details on how to resample datasets using destructive sampling. S represents the
number of strata from one individual. LK represents the lag i.e., the number of strata to remove to meet the assumption of temporal independence. ... 38 Figure 1.2. Comparison of average ratios between robust estimates of variance (VR/VT),
dashed lines) or naive estimates of variance over true variance (VN/VT,dotted lines) of coefficient β1 when P = 10 for different number of clusters (K), as a function of temporal autocorrelation (ρ) and inter-individual heterogeneity (σH2 on the right side of the panels), as well as different data processing: a) Balanced, each cluster has the same number of strata (S = N/K); b) Weakly Unbalanced, K/3 clusters have S/2 strata and 2K/3 clusters have S strata; c) Strongly Unbalanced, K/3 clusters have S/4 strata, K/3 clusters have S/2 strata and K/3 clusters have S strata; d) Destructive sampling, each initial cluster of S strata has been split into 2 clusters, a variable number of strata had been dropped in between to meet the assumption of independence between clusters. Robust or naive estimates of variance are unbiased when ratios are not significantly different from 1 at the 5% level (solid line). ... 40
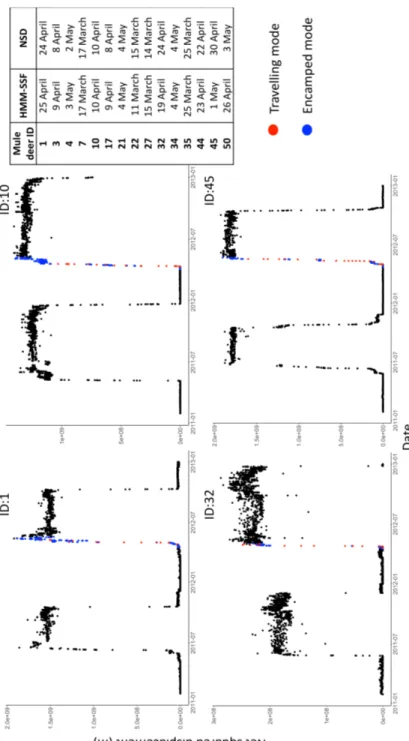
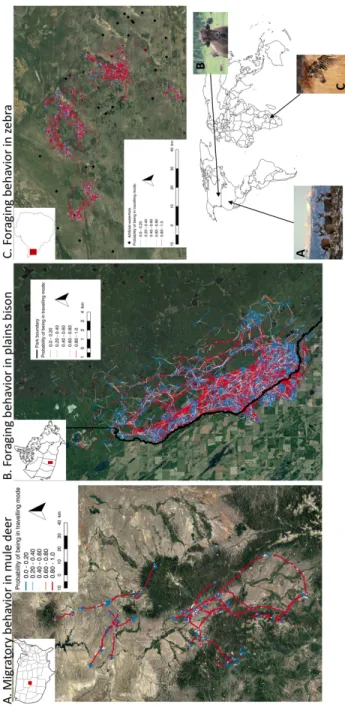
Figure 2.1. Net squared displacement over time for 4 mule deer geolocalised every 3 hours between 2011 and 2012 in Medicine National Bow Forest (WY-CO, USA). Net squared displacement is measured as the square of the Euclidean distance between location at a specific time t and start location (i.e., at time 1). Estimated mode of movement (red for travelling and blue for encamped) during the 2012 migration was based on the conditional probabilities of being in encamped and travelling state at each step, as estimated from the HMM-SSF, and dichotomized to 0-1 using a 0.50 threshold. ... 71 Figure 2.2. Trajectories of 42 plains bison geolocalised every hour between 2005 and 2016,
during summer season in Prince Albert National Park (SK, Canada), 15 mule deer geolocalised every 3 hours during 2012 spring migration in and around Medicine Bow National Forest (WY-CO, USA) and 17 zebras geolocalised every half hour between 2012 and 2014, during dry hot season in Hwange National Park (Zimbabwe). The conditional probability of being in travelling mode of movement at each step, estimated from the HMM-SSF, is represented using a blue (low) to red (high) gradient. ... 72
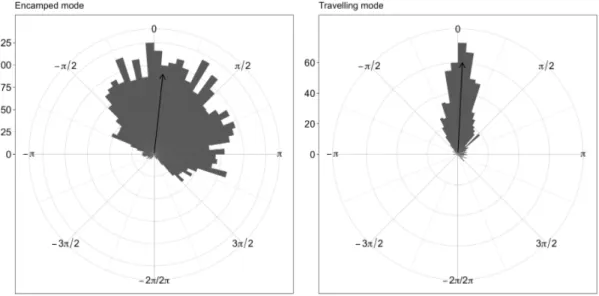
Figure 2.3. Distribution of directional bias (in radians) towards summer range visited in 2011 by mule deer in Medicine Bow National Forest, while they were migrating during spring 2012, according to the mode of movement estimated by the HMM-SSF at each step (either encamped or travelling). Estimated mode of movement was based on the conditional probabilities of being in encamped and travelling state at each step, truncated to 0-1 using a 0.50 threshold. The arrows indicate the average directional bias. ... 73 Figure 2.4. Estimated probability of switching from encamped to travelling mode of
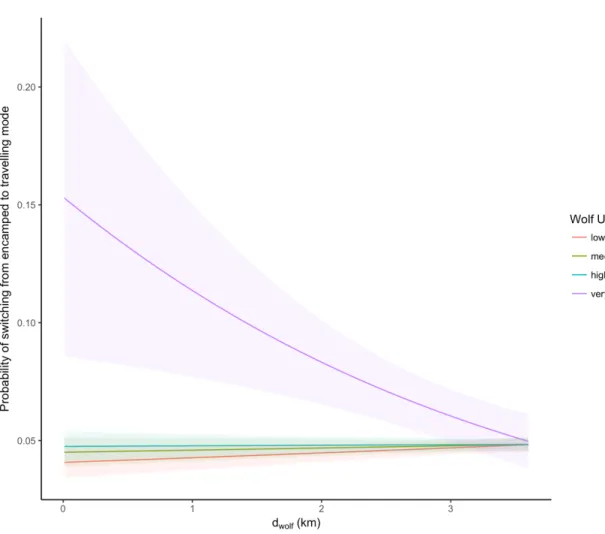
movement and 95% confidence interval, in plains bison during summer, according to wolf distance and local intensity of wolf utilization distribution (wolf UD). Coefficient estimates and standard errors were obtained from the fit of a mixed-effects generalized linear model with binomial distribution on 27,101 geolocations from 32 plains bison followed between 2007 and 2016 in Prince Albert National Park (SK, Canada).
Probabilities were calculated here, by setting period of day and meadow variables to day and 1, respectively (see Table 2.2). ... 74
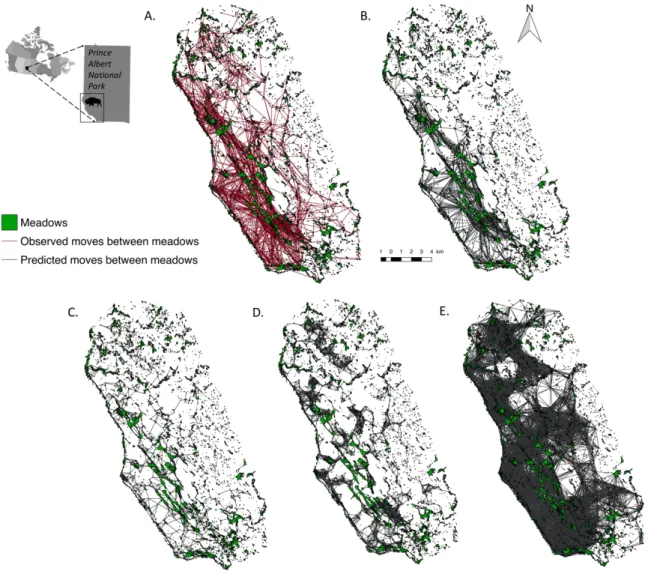
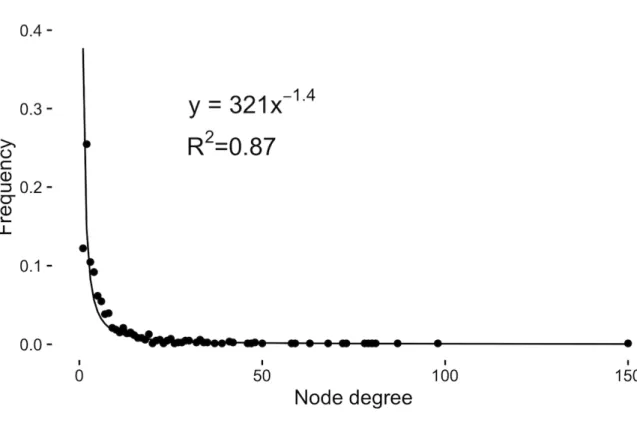
Figure 3.1. Graphical representation of different network structures to model landscape functional connectivity of the population of plains bison in Prince Albert National Park during summer. A) Empirical graph based on bison GPS trajectories. B) Ultra-small world scale-free network. C) Minimum planar graph. D) 700-m distance threshold network. E) 1800-m distance threshold network. Bison occupy the southwestern corner of the park during summer time (black rectangle on the map of the park). ... 95 Figure 3.2. Degree distribution (black dots) of the empirical spatial network of plains bison in Prince Albert National Park based upon the movements of 43 GPS-collared individuals in the summers 2005-2015. The distribution decays as a power function, i.e., y ∝ x-a, with
a ± s. e = 1.4 (±0.073) (solid line), as expected for scale-free networks. ... 96 Figure 3.3. Relative intensity of meadow use estimates for plains bison in Prince Albert
National Park: (A) based upon geolocations of 22 bison during summer over 8 years of monitoring (2005, 2007, 2010-2015); predicted by the reaction-advection-diffusion model while considering different network structures (B) ultra-small world scale-free network, (C) minimum planar graph, (D) 700-m distance threshold network, (E) 1800-m distance threshold network. Goodness-of-fit of (B), (C), (D), and (E) to (A) are respectively pseudo-RB2 = 0.74, pseudo-RC2= 0.21, pseudo-RD2= 0.21 and pseudo-RE2 = 0.37. ... 97
Figure 3.4. Change in relative intensity of meadow use estimates for plains bison in Prince Albert National Park: predicted by the reaction-advection-diffusion model while considering different network structures for implementing the removal of the 10 most connected meadows (yellow stars) (A) ultra-small world scale-free network, (B) minimum planar graph, (C) 700-m distance threshold network and (D) 1800-m distance threshold network. The value represents the change in the relative intensity of meadow use in
comparison to model prediction before meadow removal (Fig. 3. 3). ... 98
Figure 4.1. Relative intensity of patch use estimates for woodland caribou, in the Côte-Nord region of Quebec for both before (left panels) and after (right panels) patch disturbance
in one experimental site, based on geolocations of one caribou during winter 2009
(bottom panels) or predicted by the reaction-advection-diffusion model (top panels). ... 119 Figure 4.2. Changes in four indices of connectivity (i.e., link density, number of components,
diameter of the network and characteristic path length) in 10 experimental sites categorized either as hub (n=4) or non-hub (n=6) disturbed network. Network was categorized as hub disturbed when at least one disturbed node was a hub (i.e., its degree exceeded 465 links). (*) indicates that the change in connectivity index significantly differs from 0 according to a linear mixed effect model. Observed data are represented using grey triangles behind each boxplot. ... 120 Figure 4.3. Predicted (left) and observed (right) change in caribou space use during winter in
10 experimental sites categorized either as hub (n=4) or non-hub (n=6) disturbed network. Network was categorized as hub disturbed when at least one disturbed node was a hub (i.e., its degree exceeded 465 links). The index of change in caribou space-use varies between 0 and 1, and indicates whether caribou returned to all previously visited patches (change = 0) or did not revisit any of them (change = 1). Changes in space use were
significantly different between hub and non-hub disturbed networks (*), according to a linear mixed effect model. Observed data are represented using grey triangles behind each boxplot. ... 121
Liste des tableaux
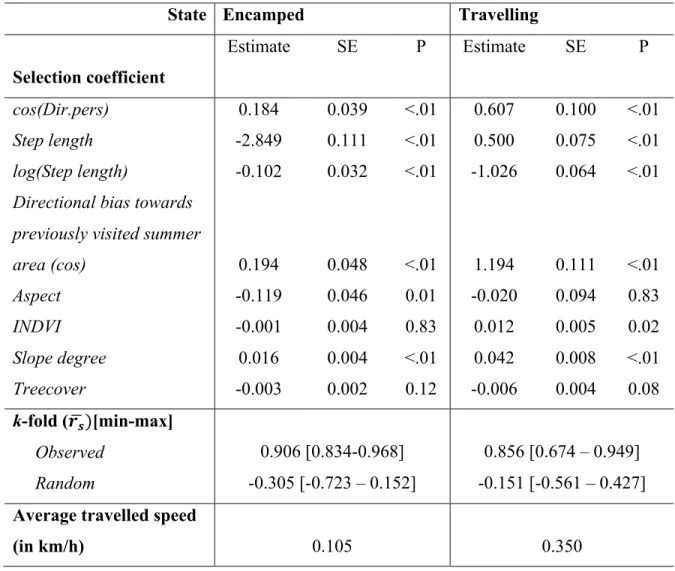
Table 2.1. Assessment of the HMM-SSF robustness from simulations to predict habitat selection along encamped and travelling movement in three ecological scenarios, using k-fold validation. Means, minimums and maximums ([min - max]) of 100 Spearman rank correlation are reported for each behavioural mode of movement and both observed and random steps. ... 66 Table 2.2. State-specific selection coefficient estimates along with their standard-error (SE)
and associated P-value (P) of the HMM-SSF used to predict animal movements according to encamped or travelling mode, in three ecological systems. A. Habitat selection during the different phases of the migratory behavior, B. and C. Habitat selection during the different phases of the foraging behavior. Analysis were performed using 13,444, 94,686 and 72,730 geolocations from 15 mule deer, 42 plains bison and 18 zebras over 2 (2011-2012), 11 (2005-2016) and 3 (2012-2014) years of monitoring, respectively. The predictive capability of HMM-SSF was assessed using k-fold validation repeated 100 times, means and ranges of spearman correlation (rs) are reported for each behavioral mode of movement and both observed and
random steps. Average travelled distances were calculated using step length and
log-transformed step length coefficient estimates (see Appendix 2.1). ... 67 Table 2.3. Coefficient estimates along with their standard-error (SE) and associated P-value
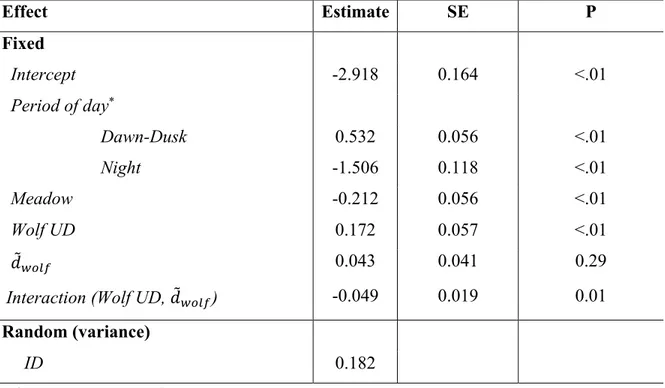
(P) of the mixed-effects generalized linear model with binomial distribution to predict probability that a bison switched from encamped to travelling mode during its travel in summer. We used a probability threshold of 0.50 (Pthreshold) to determine transition and non-transition from the conditional probabilities of being in encamped or travelling state, as estimated from the HMM-SSF. Mixed-effects generalized linear model was fitted using 27,101 geolocations from 32 GPS-collared plains bison in Prince Albert National Park (SK, Canada). dwolf was set to dwolf when dwolf ≤ 3.6 km and 3.6 km, otherwise. ... 70 Table 4.1. Hypothesis testing and predictions supported to evaluate both the robustness of a
scale-free spatial network (A) and space-use reorganisation towards disturbance of hub or non-hub resource patch (B). Node degree = number of links connected to a node; Link density = number of existing links divided by the number of possible links; Number of components = number of sub-graphs in the network; Diameter of the network = maximum distance, in number of links, between any connected pair of nodes in the network;
Characteristic path length (CPL) = average distance between any connected pair of nodes in the network (Rayfield et al. 2011). Predictions are made considering that a disturbance does not remove entirely a node but instead splits it up into several smaller nodes. ... 117
Remerciements
Cette thèse est l’aboutissement de quatre années de travail avec l’aide et la collaboration de plusieurs personnes. Je tiens à exprimer ma gratitude à l’ensemble d’entre elles.
Je remercie tout d’abord mes directeurs de thèse, Daniel Fortin (directeur principal) et Thierry Duchesne (co-directeur) pour m’avoir permis de faire mon doctorat à leurs côtés et d’avoir toujours été présents pour m’aider et répondre à mes questions. Nos discussions et votre intérêt pour mon travail ont fait progresser mes capacités de réflexion et d’écriture. Un grand merci !
Je remercie également André Fortin pour avoir été dans mon comité d’encadrement et m’avoir aidée dans le développement du modèle mathématique utilisé dans plusieurs chapitres de ma thèse. Cette aide a rendu la tâche plus facile qu’attendue. Je le remercie également pour m’avoir donné la possibilité de présenter une partie de mes travaux de doctorat à ses élèves au baccalauréat.
Je remercie Jean-Pierre Tremblay, le quatrième membre de mon comité d’encadrement, pour m’avoir suivie tout au long de ces années et m’avoir donné des commentaires extérieurs constructifs lors de nos rencontres. Je remercie Ladd Johnson, Gilles Gauthier et Steeve Côté pour avoir été sur mon comité d’examen doctoral, ainsi que Louis Dubé et Jean Gabriel Young avec qui j’ai eu des discussions fructueuses sur mes analyses réseaux.
I would like to thank all my co-authors: Simon Chamaillé-Jammes, Pierre Drapeau, Matthew Kauffman, Jerod Merkle, Louis-Paul Rivest and Martin-Hugues St-Laurent for their constructive and profound comments that improved my manuscripts. I also thank you for sharing your data.
Le Fonds de recherche nature et technologie du Québec, le Conseil de recherches en sciences naturelles et en génie du Canada, la Chaire industrielle de recherche de l’Université Laval en sylviculture et faune de la forêt boréale, le Centre d’étude de la forêt, et les Fonds Richard Bernard et de la Société Provancher ont été des soutiens financiers tout au long de mon doctorat, sans quoi ma thèse n’aurait pas eu lieu. Merci à ces partenaires financiers. Je remercie aussi Parcs Canada pour m’avoir autorisée à faire du terrain dans le parc national de Prince Albert, en Saskatchewan, ainsi que Norm Stolle pour avoir partagé les localisations des bisons et des loups.
Ma gratitude va également à Fréderic Maps pour m’avoir offert la chance d’être auxiliaire d’enseignement pour son cours de biostatistiques. Cette expérience fut divertissante et enrichissante.
Je remercie les membres du secrétariat du Département de biologie de l’Université Laval : Louise Lapointe, Josée Verret, Marie-Soleil Desjardins ainsi que Marie-Claude Martin, technicienne du Département de biologie, pour leur aide technique et administrative tout au long de ces quatre années.
Je voudrais remercier tous les membres actuels et anciens du laboratoire de Daniel Fortin, avec qui j’ai passé de très bons moments et qui ont rendu mon doctorat plus agréable : Florian Barnier, Audrey-Jade Bérubé, Orphé Bichet, Léa Harvey, Guillemette Labadie, Tom Mason, Jerod Merkle, Ricardo Simon, Alexandre Terrigeol, Virginie Vanlandeghem. Ainsi que ceux avec qui j’ai eu de brèves discussions (mais intéressantes et utiles!): Mathieu Basille, Nicolas Courbin, Karine Dancose et Marie Sigaud. Je tiens aussi à remercier Aurélien Nicosia, ancien étudiant au doctorat dans le laboratoire de Thierry Duchesne, avec qui j’ai eu des discussions scientifiques très intéressantes et de très bonnes discussions amicales.
Une grande pensée va à mes colocataires de bureau et d’appartement : Antoine Boudreau Leblanc, Morgan Gazzola, Laurent Hany, Nicolas Houde, Gaëtan Le-bel et Ricardo Simon. Vous avez égaillé ma vie de tous les jours donc merci !
Je remercie du fond du cœur ma famille proche pour leur soutien : mes parents, mon frère, sa conjointe et leur petit garçon. Je sais que la distance n’a pas été facile tous les jours mais vous étiez toujours dans mon cœur pendant ces quatre années.
Finalement je remercie Ricardo, qui de collègue est devenu mon conjoint. Je n’aurais jamais imaginé que notre mois de terrain aurait fini sur cette note ! Le doctorat est une étape de vie pleine de surprises et celle-ci en fût une très bonne. Je te suis très reconnaissante pour tous les conseils que tu me donnes et les discussions scientifiques (ou pas) que nous pouvons avoir. Sans oublier toute la bonne humeur que tu me procures. Une page se tourne avec cette thèse mais une autre va bientôt s’ouvrir pour de nouvelles aventures tous les deux, j’ai hâte !
Avant-propos
Ce doctorat est présenté sous la forme d’une thèse comprenant quatre articles scientifiques. La thèse inclut également une introduction générale et une conclusion générale qui lient l’ensemble des articles, bien que chacun puisse être lu indépendamment.
En tant qu’auteure principale des quatre articles, j’ai élaboré les objectifs de recherche, j’ai effectué le travail de modélisation et les analyses statistiques, et rédigé les manuscrits. Mon directeur de thèse, Daniel Fortin, a largement participé au développement des objectifs et de révision des manuscrits. Mon co-directeur, Thierry Duchesne, a fortement contribué à la révision des analyses statistiques et également à la rédaction des manuscrits. Je dois également souligner la collaboration d’André Fortin, qui a participé à l’élaboration du modèle de réaction-advection-diffusion utilisé dans les chapitres 3 et 4.
Mon chapitre 1 est publié dans PLOS One, avec Thierry Duchesne et Daniel Fortin comme coauteurs. Le chapitre 2 est en cours de révision dans Ecological Monographs avec Thierry Duchesne, Jerod Merkle, Matthew Kauffman, Simon Chamaillé-Jammes et Daniel Fortin comme coauteurs. Le chapitre 3 est publié dans Methods in Ecology and Evolution, avec Thierry Duchesne, André Fortin, Louis-Paul Rivest et Daniel Fortin comme coauteurs. Le chapitre 4 est en cours de révision dans Functional Ecology avec Thierry Duchesne, André Fortin, Louis-Paul Rivest, Pierre Drapeau, Martin-Hugues St-Laurent et Daniel Fortin comme coauteurs.
J’ai également contribué à deux autres articles scientifiques en dehors de mon travail de thèse. J’ai participé aux analyses statistiques et à la révision du manuscrit de l’article de Fortin et coll. (2017), qui a été publié dans Scientific Reports, pour lequel je suis septième auteure. J’ai également contribué à la préparation et à l’extraction de données spatiales, ainsi qu’à leurs analyses, pour le projet de maîtrise de Virginie Vanlandeghem. Un article est actuellement en cours de rédaction dans lequel je suis une des coauteurs.
Introduction générale
1. La répartition spatiale des populations animales : un thème majeur en
écologie
L’écologie est une science qui vise à comprendre les fluctuations d’abondance et de répartition spatiale des espèces dans différents contextes (Elton 1927). La répartition spatiale des populations est donc un thème central à l’écologie animale et elle est étudiée depuis longtemps (Andrewartha & Birch 1954; Macfadyen 1963). Par exemple, Lack (1933) a illustré l’effet d’un déboisement sur la répartition spatiale de plusieurs espèces d’oiseaux dans la région de Breckland, en Angleterre. Ou encore, Platt et al. (1957) ont expliqué en partie l’aire de répartition en milieu naturel d’une espèce de moustique (Anopheles quadrimaculatus) par un ensemble de conditions physiques (c.-à-d., température, humidité et luminosité) préféré par ces insectes, à l’aide d’expériences en laboratoire.
L’intérêt scientifique pour l’étude de l’utilisation de l’espace par les populations animales provient du fait que la répartition spatiale et l’abondance des individus sont couplées. En effet, les conditions écologiques rencontrées par les individus dans l’espace influencent leurs chances de survivre et de se reproduire, ce qui ultimement détermine l’abondance de la population (Andrewartha & Birch 1954; Dussault et al. 2012; Losier et al. 2015; Lafontaine et al. 2017; Sigaud et al. 2017). La recherche fondamentale sur l’écologie spatiale des espèces peut également bénéficier à la conservation de populations en déclin ou à la gestion de conflits humain-faune (Festa-Bianchet & Apollonio 2003; Lesmerises et al. 2013; Marley et al. 2017). Prenons, par exemple, le cas des tortues caouanne (Caretta caretta), qui sont listées vulnérables par l’union internationale pour la conservation de la nature (IUCN 2015). La géolocalisation de plusieurs individus a permis d’identifier des corridors de migration et ainsi de proposer des mesures visant à réduire et à ajuster les activités de pêche dans ces zones, afin de minimiser l’impact négatif des captures involontaires sur l’abondance de l’espèce (Morreale et al. 1996; Polovina et al. 2000; Luschi 2003).
L’utilisation de l’espace par les animaux fait intervenir l’hétérogénéité de l’habitat (Klopfer 1969). Un paysage est considéré comme hétérogène lorsque celui-ci est perçu par les individus comme un ensemble de parcelles, chacune comparativement uniforme dans ses caractéristiques écologiques et séparée notablement de ses voisines (Elton 1966; Klopfer 1969). Chez la plupart des espèces, un animal ne peut pas rester éternellement au même endroit, d’une part sa nourriture finirait par manquer et d’autre part, ses besoins concordent souvent avec des milieux différents (p. ex., refuge pour échapper à la prédation versus alimentation, Fortin et al. 2005). Par conséquent, la répartition spatiale des populations animales est généralement influencée par l’organisation spatiale des différentes parcelles d’habitat, qui sont nécessaires pour subvenir aux besoins des individus (Dunning et al. 1992; Taylor et al. 1993; Courbin et al. 2014; Owen-Smith & Traill 2017).
Le développement des outils télémétriques, mathématiques et statistiques nous permet dorénavant de comprendre plus en détails la répartition spatiale des populations animales, notamment en mettant en évidence les mécanismes sous-jacents associés à l’utilisation de l’espace (Moorcroft 2012). Par exemple, Merkle et al. (2014) ont démontré, à partir d’un modèle statistique de choix des parcelles, que la mémoire des individus influence les déplacements des bisons des prairies (Bison bison bison), résultant en une utilisation de l’espace restreinte aboutissant à la formation d’un domaine vital (Merkle et al. 2016b). Cette compréhension est utile pour anticiper les changements de répartition spatiale lorsqu’une modification s’opère dans le paysage (Moorcroft et al. 2006), ou pour identifier par quels mécanismes l’utilisation de l’espace influence les paramètres démographiques d’une population (Losier et al. 2015; Fortin et al. 2017). Ainsi, la méthodologie prend une place importante en écologie, puisque le développement de notre compréhension mécaniste de la répartition spatiale des populations animales repose en partie sur la disponibilité des modèles statistiques et mathématiques (Moorcroft & Lewis 2006).
2. Anticiper la répartition spatiale d’une population dans un
environnement dynamique
Qu’elles soient d’origine naturelle (p. ex., feu, épidémie, inondation) ou anthropique (p. ex., agriculture, urbanisation, exploitation forestière), les perturbations entraînent des changements de composition et de configuration du paysage (Delcourt et al. 1982; Urban et al. 1987; Collinge 2009). Ces modifications peuvent impacter la répartition spatiale des populations animales, par exemples en remaniant la taille des parcelles disponibles aux individus ou en créant des barrières aux déplacements dans le paysage (Andreassen & Ims 1998; Courtois et al. 2003, 2007; Bartzke et al. 2015). L’évaluation de la répartition spatiale des caribous des bois (Rangifer tarandus), un an après l’occurrence d’une perturbation anthropique (c.-à-d., coupe forestière et construction de chemins forestiers), a montré que les individus se relocalisaient à une distance de 4 km de la perturbation, résultant en une augmentation locale de la densité de caribous à la périphérie des zones perturbées (Fortin et al. 2013). Cette réorganisation spatiale des caribous peut notamment modifier les interactions de cette proie avec son prédateur, le loup gris (Canis lupus), puisque l’augmentation locale de la densité de caribous peut fournir des zones ciblées de chasse (Fortin et al. 2013). La modification de la structure et de la composition du paysage suite à une perturbation peut, de ce fait, avoir des conséquences indirectes sur les interactions trophiques et ultimement sur l’abondance des espèces et sur le fonctionnement d’un écosystème (Wittmer et al. 2005; Fahrig 2007; Collinge 2009; Doherty & Driscoll 2018). Dans la mesure où les perturbations naturelles et anthropiques sont en augmentation (Diffenbaugh & Field 2013; Müller et al. 2018), il est de plus en plus urgent d’obtenir des outils de modélisation robustes et efficaces pour correctement anticiper les changements de répartition spatiale des populations animales dans des environnements dynamiques (Elith & Leathwick 2009; Moorcroft 2012; Ehrlén & Morris 2015).
Les modèles prédictifs de l’utilisation de l’espace peuvent également s’avérer utiles pour la gestion ou la conservation des espèces. En effet, les modèles spatialement explicites peuvent assister les gestionnaires dans leur prise de décisions, afin d’opter pour la stratégie la plus adéquate pour atteindre l’objectif de gestion ou de conservation (Risenhoover et al. 1997; Marley et al. 2017). Par exemple, un simulateur de gestion des cervidés fût développé
pour tester l’efficacité de plusieurs scénarios de gestion de populations de cervidés en surabondance (Risenhoover et al. 1997). Notamment, le modèle projette la répartition spatiale d’une population selon une intervention de gestion (p. ex., mise en place de barrières), et permet ainsi d’identifier les zones potentielles de forte et faible utilisation par les cervidés selon l’alternative testée. Plus précisément, le modèle simule la dispersion des cervidés dans un paysage hétérogène en modélisant les déplacements des individus en réponse aux caractéristiques du paysage. Ce modèle acquiert sa capacité de prévision de l’utilisation de l’espace, lorsque le paysage change, grâce à l’utilisation d’une approche mécaniste, c’est-à-dire qu’il intègre une règle de déplacement sous-jacente à l’utilisation de l’espace par les cervidés (Moorcroft et al. 2006). Similairement, Moorcroft et al. (2006) ont développé un modèle mécaniste de l’utilisation de l’espace par les coyotes (Canis latrans), basé sur une marche aléatoire corrélée et biaisée, qui intègre deux règles de déplacement suivies par les individus. Les coyotes se déplaçaient vers un point central de leur domaine vital lorsqu’ils flairaient une odeur étrangère, et ils biaisaient également leurs déplacements vers les zones riches en proies (Moorcroft et al. 2006). Les auteurs ont particulièrement démontré l’utilité du modèle pour correctement prévoir le changement de répartition spatiale des coyotes suite à une perturbation (Moorcroft et al. 2006). Malgré son succès à prévoir proprement l’utilisation de l’espace par les coyotes, l’application de ce modèle à des espèces qui s’alimentent dans des parcelles spatialement fixes (p. ex., les herbivores) peut être limitée, puisqu’il ne considère pas l’intensité de l’utilisation des parcelles (p. ex., le temps de résidence dans une parcelle) (Bailey et al. 1996; Owen-Smith & Traill 2017). Afin d’acquérir des modèles de prévision les plus réalistes possible, il est nécessaire d’orienter leurs développements vers des outils intégratifs pour tenir compte des différents processus sous-jacents à l’utilisation de l’espace par les populations animales.
Les modèles basés sur l’individu s’avèrent également utiles pour construire des modèles mécanistes de l’utilisation de l’espace (Latombe et al. 2014b; Marley et al. 2017; Tardy et al. 2018). Des règles de déplacement sont utilisées pour générer des trajectoires individuelles (Tardy et al. 2018) et la répartition spatiale de la population peut être obtenue à partir de ces multiples trajectoires simulées (Giuggioli et al. 2011). Cependant, les modèles basés sur l’individu qui intègrent des règles de déplacement complexes sont souvent longs à mettre en œuvre et demandent d’importantes ressources de calcul (Avgar et al. 2016; Tardy
et al. 2018), ce qui peut décourager les utilisateurs à développer des modèles réalistes, de par leur complexité, ou limiter leur utilisation par les gestionnaires. Dans le but d’obtenir une utilisation la plus répandue possible, le développement d’outils de modélisation devrait également promouvoir des modèles flexibles, efficaces, simples d’implémentation et d’utilisation.
3. L’approche mécaniste de la répartition spatiale des populations : une
approche basée sur les déplacements
3.1 Les différentes composantes mécanistes du déplacement
Les décisions prises par les animaux lors de leurs déplacements reflètent une interaction complexe entre leurs besoins de se déplacer pour accéder à leurs ressources (p. ex., alimentation, refuge, Morales et al. 2004; Nathan et al. 2008), la connaissance qu’ont les individus de leur environnement et leur capacité de navigation (p. ex., le déplacement vers une parcelle connue et riche en nourriture, Nathan et al. 2008; Merkle et al. 2014), la mobilité des individus (p. ex., la distance parcourable par unité de temps, Nathan et al. 2008) et les facteurs externes associés à l’hétérogénéité du paysage (p. ex., la présence d’une barrière ou d’un corridor, Bartzke et al. 2015). L’ensemble de ces décisions peut être vu comme la réponse à quatre questions : où, quand, comment et pourquoi se déplacer (Pyke 1983; Nathan et al. 2008) ? Savoir répondre à ces questions fondamentales du déplacement animal nous offre ainsi une compréhension mécaniste des processus sous-jacents à l’utilisation de l’espace, qui est nécessaire à l’obtention de modèles prédictifs réalistes de répartition spatiale des populations animales dans des environnements hétérogènes (Nathan et al. 2008; Moorcroft 2012). Spécifiquement, la réponse à la question pourquoi se déplacer couvrent les bénéfices proximaux (p. ex., aller dans une parcelle de ressource pour se nourrir afin d’atteindre la satiété) et ultimes (p. ex., aller dans une parcelle de ressource qui permet de maximiser son gain énergétique et donc sa valeur adaptative) au déplacement de telle sorte à ce que cette approche soit bien intègre des mécanismes fondamentaux à l’utilisation de l’espace (Nathan et al. 2008).
3.2 Les facteurs influençant les déplacements
Les animaux se déplacent d’une place à une autre pour une variété de raisons dont la recherche de nourriture (Pyke 1983; Bailey et al. 1996; Owen-Smith et al. 2010; Viswanathan et al. 2011). Cependant, il a été observé chez certaines populations une répartition qui n’est pas parfaitement liée avec la répartition de leurs ressources, signifiant que d’autres facteurs sont en cause (Laundré 2010; Courbin et al. 2014). Le risque de prédation semble notamment être un facteur majeur influençant les déplacements des individus (Lima & Dill 1990; Fortin et al. 2005; Harvey & Fortin 2013). En effet, les individus peuvent employer des stratégies proactives (p. ex., éviter les zones risquées, Fortin et al. 2005; Creel et al. 2014; Latombe et al. 2014a; Padié et al. 2015) et réactives (p. ex., fuir lorsqu’un prédateur est proche, Wirsing et al. 2010; Creel et al. 2014; Courbin et al. 2016), afin d’éviter la prédation. De même, l’agencement spatial du paysage (Courbin et al. 2014), la présence de congénères (Fortin & Fortin 2009) ou encore l’expérience passée (Merkle et al. 2014) peuvent également influencer la répartition spatiale des populations animales.
3.3 Identification des facteurs influençant les déplacements
Les analyses de sélection des ressources sont utiles pour identifier les parcelles de l’habitat et les aires qui sont utilisées disproportionnellement par rapport à leur disponibilité (Manly et al. 2002; Fortin et al. 2005). Ces analyses statistiques permettent de comprendre les liens entre l’utilisation relative de l’espace par les individus et différentes formes d’hétérogénéité spatiale (p. ex., type d’habitat, risque de prédation, disponibilité des ressources, Fortin et al. 2009; Merkle et al. 2014; McMahon et al. 2017). Particulièrement, les fonctions de sélection de pas (« step selection functions », SSF sensu Fortin et al. 2005) permettent d’identifier la sélection relative des attributs du paysage au cours des déplacements des individus. À partir de la trajectoire des individus, une SSF compare les attributs du paysage associés aux pas observés (c.-à-d., segments entre deux occurrences successives d’un individu), aux attributs du paysage associés à plusieurs pas aléatoires que les individus auraient pu choisir lors de leurs déplacements (Fortin et al. 2005; Thurfjell et
al. 2014). Les caractéristiques des pas (p. ex., tortuosité et longueur) peuvent aussi être intégrées dans une SSF, fournissant ainsi un cadre mécaniste pour comprendre où, comment et pourquoi les individus se déplacent. Cependant, l’analyse de la dynamique temporelle des déplacements (p. ex., quand quitter une parcelle) est habituellement découplée des analyses de sélection des ressources. Notamment, les mécanismes associés à la dynamique temporelle des déplacements sont généralement identifiés par des analyses statistiques de temps de résidence dans les parcelles (p. ex., à partir d'un modèle de Cox, Courbin et al. 2014; ou d'une régression de Poisson, McMahon et al. 2017).
4. Les données de déplacement: opportunités et défis associés à l’essor des
technologies GPS
La radio-télémétrie est un outil essentiel en écologie animale puisqu’elle permet de suivre des espèces sur une longue période de temps (p. ex., plusieurs années), même lorsque les individus se déplacent sur de grandes distances (Kojola et al. 2006; Lesmerises et al. 2013; Wilson et al. 2014). L’augmentation de la capacité des batteries des colliers équipés d’un système de positionnement global (GPS) favorise la collecte des données à haute fréquence temporelle alors que la miniaturisation des colliers permet maintenant de suivre des espèces de petite taille (Stevenson et al. 2013). La diversité et l’augmentation de la fiabilité (c.-à-d., moins d’erreurs de localisation) des données obtenues avec les colliers GPS font qu’ils sont considérés comme une des meilleures technologies disponible pour déterminer les patrons de déplacement et de sélection des ressources par les animaux (Millspaugh & Marzluff 2001).
4.1 La corrélation dans les données de déplacement
Malgré leur importance capitale pour comprendre l’utilisation de l’espace à fine échelle par les populations animales, les données de déplacement récoltées à haute fréquence et sur plusieurs individus présentent plusieurs défis associés à leurs analyses. C’est le cas,
notamment, de la gestion de la corrélation qui peut être présente dans les données longitudinales, c’est-à-dire collectées sur un même individu au cours du temps (Fieberg et al. 2010; Fitzmaurice et al. 2011). L’autocorrélation temporelle, souvent inhérente aux jeux de données longitudinales, résulte en une ressemblance des données collectées près dans le temps et qui s’amenuise puis disparait au fur et à mesure que l’on compare des données de plus en plus éloignées temporellement (Fieberg et al. 2010). De plus, les individus peuvent réagir différemment à l’hétérogénéité spatiale pour des raisons d’âge, de sexe, de statut social ou encore de condition physique (Stamps & Swaisgood 2007; Bonnot et al. 2015; Marchand et al. 2015; Rossman et al. 2015). De ce fait, les données d’un même individu peuvent être plus semblables entre elles que si comparées aux données d’un autre individu. Ces deux sources de corrélation peuvent entraîner une sur- ou sous-estimation des variances des estimateurs des coefficients de sélection si elles ne sont pas correctement prises en compte (Gillies et al. 2006). Les estimations des variances des estimateurs des coefficients sont particulièrement utilisées pour calculer les valeurs de P, qui informent sur les facteurs influençant la variable réponse considérée (Rencher & Schaalje 2008). Ainsi, ne pas considérer la corrélation dans les données de déplacement pourrait nous amener à des conclusions erronées sur les mécanismes associés à la répartition spatiale des individus dans un environnement hétérogène.
L’estimation par équations d’estimation généralisées (GEE) est une approche statistique qui permet d’estimer d’une manière robuste les variances des estimateurs des coefficients d’un modèle de régression, de telle sorte que les estimations des variances soient corrigées pour tenir compte de la corrélation dans les données empiriques (Hardin 2005). Pour ce faire, les GEE utilisent des « grappes » pour estimer la structure de la corrélation empirique (Hardin 2005). Chaque grappe représente un ensemble de données corrélées, mais les grappes doivent être indépendantes entre elles.
Afin de tenir compte de la corrélation dans les données de déplacement, l’inférence sur les paramètres d’une fonction de sélection de pas peut être effectuée par la méthode des équations d’estimation généralisées (Craiu et al. 2008). Dans une SSF, le pas observé et les pas aléatoires associés sont regroupés en strates (Fortin et al. 2005). Cette stratification des données permet notamment d’identifier les attributs du paysage qui influencent les
déplacements et l’utilisation de l’espace à fine échelle, à partir d’une régression logistique conditionnelle (Fortin et al. 2005). Lorsque la méthode des GEE est utilisée pour inférer les paramètres de la régression logistique conditionnelle, les strates successives d’un même individu sont soit assignées à une même grappe, soit séparées en plusieurs grappes (Fortin et al. 2005; Zimmermann et al. 2014). La séparation en plusieurs grappes des strates d’un même individu nécessite l’élimination de strates intermédiaires jusqu’à obtenir l’indépendance temporelle entre les strates appartenant à des grappes séparées. Ceci résulte en une forte variation sur le nombre total de grappes (p. ex., 14 à 450) utilisé pour estimer la corrélation empirique, et une grande perte de données lorsque la division des strates d’un même individu est effectuée intensément (Babin et al. 2011; Courbin et al. 2014; Zimmermann et al. 2014). Le partitionnement des strates en grappes est une étape importante puisqu’elle détermine l’efficacité des GEE à corriger le biais statistique des erreurs standards associé à la corrélation dans les données de déplacement (Hardin & Hilbe 2002). Cependant, les règles de partitionnement sont très variables dans la littérature, sans connaître leurs conséquences sur le succès des GEE à correctement estimer la sélection de l’habitat lors des déplacements des individus (Forester et al. 2009; Courbin et al. 2014; Zimmermann et al. 2014; Lewis et al. 2015). Il est donc important d’identifier quelles règles de partitionnement des données corrélées de déplacement en grappes permettent aux GEE de corriger le biais statistique dans les estimations des variances des estimateurs des coefficients de sélection, sans perdre une trop grande quantité d’information biologique.
4.2 L’identification des phases comportementales à partir des données de déplacement
L’augmentation de la disponibilité des données de déplacement, via l’essor des technologies GPS, fournit également des opportunités pour mieux comprendre plusieurs processus écologiques associés aux déplacements (Cagnacci et al. 2010). En effet, les données de déplacement peuvent nous informer sur différentes phases comportementales par lesquelles passent les individus à partir de la forme de leur trajectoire (Johnson et al. 2002; Fauchald & Tveraa 2003; Franke et al. 2004). Par exemple, les individus optent souvent pour des déplacements sinueux et courts lorsqu’ils s’alimentent (c.-à-d., mode de déplacement
campé), alors qu’ils vont parcourir de plus grandes distances en ligne droite lorsqu’ils se déplacent d’une parcelle de ressources à une autre (c.-à-d., mode de déplacement exploratoire) (Bailey et al. 1996; Morales et al. 2004). L’intégration de ces phases comportementales dans les analyses de sélection de l’habitat peut nous aider à identifier les utilisations spécifiques de certains types d’habitat (Roever et al. 2014; Marchand et al. 2015; Abrahms et al. 2016). Par exemple, Roever et al. (2014) démontrent que certains éléphants (Loxodonta Africana) sélectionnent les points d’eau en mode campé mais les évitent en mode exploratoire. Tenir compte de ces phases comportementales est donc important pour correctement quantifier la sélection de l’habitat, améliorer notre compréhension des mécanismes associés à l’utilisation de l’espace et produire des modèles réalistes de répartition spatiale des populations animales (Nathan et al. 2008; Roever et al. 2014). De plus, cela permet d’identifier les habitats spécifiques à certains comportements, qui peuvent être d’intérêt pour un objectif de conservation ou de gestion (p. ex., type d'habitat associé à des longs déplacements pour identifier des corridors, Chetkiewicz et al. 2006; Roever et al. 2013). Cependant, les outils disponibles pour identifier ces phases de déplacement présentent plusieurs limites, telles que le manque d’inclusion d’information spatiale (p. ex., les attributs du paysage) pour identifier les modes de déplacement (Fauchald & Tveraa 2003; Gurarie et al. 2009) ou encore la complexité d’implémentation et de calcul, limitant ainsi le nombre de paramètres que l’on peut mettre dans le modèle (McClintock et al. 2012; Nicosia et al. 2017a; Patterson et al. 2017). Par conséquence, nous avons besoin d’un outil de modélisation flexible, robuste et simple d’utilisation pour identifier ces différentes phases comportementales.
Récemment, les fonctions de sélection de pas multi-états (HMM-SSF) ont été développées (Nicosia et al. 2017b). Celles-ci permettent d’identifier simultanément les différentes phases comportementales et le comportement de sélection associé à chacune de ces phases, fournissant ainsi une analyse plus consistante que les outils actuellement disponibles (Nicosia et al. 2017b). Cependant, plusieurs limites pourraient freiner l’utilisation de ce modèle dans les études écologiques. Premièrement, les données d’un seul individu à la fois peuvent être utilisées limitant ainsi la généralité des conclusions au niveau de la population. Deuxièmement, la trajectoire d’un individu doit être construite à partir de données recueillies à pas de temps régulier, sans données manquantes. Troisièmement, il n’y
a pas d’approche systématique qui permet de tester la validité du modèle. L’amélioration du HMM-SSF sur ces trois points permettrait notamment d’obtenir un outil flexible et facile d’emploi pour identifier les différentes phases comportementales lors des déplacements des animaux, et évaluer la sélection de l’habitat associée à ces phases comportementales.
5. Les déplacements et la théorie des graphes
Dans un paysage hétérogène, certains attributs du paysage peuvent entraver le déplacement des individus entre deux parcelles de ressources (p. ex. les routes, Shepard et al. 2008), alors que d’autres attributs peuvent faciliter les déplacements entre les parcelles (p. ex. les corridors, Haas 1995). La connectivité du paysage se mesure par le degré auquel le paysage entrave ou facilite le déplacement des individus entre leurs parcelles de ressources (Taylor et al. 1993). La connectivité structurelle du paysage se distingue de la connectivité fonctionnelle du paysage (Crooks & Sanjayan 2006). La première concerne l’organisation spatiale des différentes parcelles de ressources alors que la seconde réfère à la réponse comportementale des individus, en termes de déplacement, à cet agencement spatial des parcelles (Crooks & Sanjayan 2006). Évaluer les changements de connectivité du paysage suite à la perte d’habitat et à la fragmentation du paysage offre ainsi un moyen de comprendre et prévoir comment les déplacements des individus et leur répartition spatiale peuvent être modifiés dans des environnements altérés (Tigas et al. 2002; Collinge 2009; Courbin et al. 2014). Alors que les fonctions de sélection de pas offrent un outil pour identifier quels attributs du paysage facilitent ou entravent les déplacements des individus dans un environnement hétérogène (Fortin et al. 2005; Thurfjell et al. 2014), la théorie des graphes offre, quant à elle, un outil complémentaire pour quantifier et prévoir la connectivité au niveau du paysage tout entier dans des environnements dynamiques (Urban & Keitt 2001; Fall et al. 2007).
5.1 Qu’est-ce que la théorie des graphes ?
La théorie des graphes est un outil mathématique qui permet de représenter un système sous la forme d’un réseau, c’est-à-dire par un ensemble d’entités (c.-à-d., les nœuds)
reliées par des liens représentant une interaction, un flux ou une connexion (Harary 1969; Newman 2003). En écologie spatiale, les nœuds représentent les parcelles de ressources pour l’espèce considérée et les liens correspondent aux déplacements, potentiels ou observés, entre ces parcelles (Rhodes et al. 2006; Fall et al. 2007; Dale & Fortin 2010). De nombreux indices de connectivité peuvent être extraits d’un graphe, permettant ainsi une meilleure compréhension à la fois de la connectivité globale du paysage (c.-à-d., indices au niveau du réseau entier) et du rôle de certains nœuds ou groupes de nœuds dans la connectivité du réseau (c.-à-d., indices au niveau du nœud ou d’un groupe de nœuds connectés) (Rayfield et al. 2011). Par exemple, le degré d’un nœud correspond à son nombre de liens et il permet d’identifier les parcelles les plus connectées dans le réseau (Minor & Urban 2008; Rayfield et al. 2011). La théorie des réseaux est largement employée par les gestionnaires pour identifier les parcelles critiques à la connectivité du paysage, comparer la connectivité du paysage entre différentes alternatives de gestion ou entre différentes espèces, et évaluer des scénarios de compensation ou de restauration écologique (Lookingbill et al. 2008; Ziółkowska et al. 2012; Bergsten & Zetterberg 2013). Des applications plus théoriques de la théorie des graphes en écologie spatiale ont aussi permis de mieux comprendre le jeu spatial prédateur-proie (Courbin et al. 2014) et la propagation de pathogènes (Ferrari et al. 2014), d’identifier des patrons d’utilisation de l’espace spécifique au sexe (Jacoby et al. 2012; Stehfest et al. 2015), ou encore de tester la robustesse d’un réseau au retrait de ses nœuds ou de ses liens (Bunn et al. 2000; O’Brien et al. 2006; Treml et al. 2008; Lookingbill et al. 2010).
5.2 Prévision de la connectivité du paysage à partir de la théorie des graphes
Différentes règles de déplacement peuvent être utilisées pour décrire et prévoir la connectivité des parcelles dans un réseau (Fall et al. 2007; Minor & Urban 2008). Par exemple, le graphe planaire minimum (« minimum planar graph », MPG) considère que les parcelles de ressources sont des tremplins, de sorte que tous les nœuds sont connectés par un ensemble de liens courts sans croisement, et les liens sont ajoutés jusqu’à ce que l’on ne puisse plus en ajouter sans créer de croisement (Fig. 1) (Fall et al. 2007; Minor & Urban 2008; Courbin et al. 2014). Un MPG ne contient alors aucun raccourci, c’est-à-dire aucun
lien direct entre deux parcelles éloignées, qui croise d’autres liens et qui permet de traverser le réseau en passant par un faible nombre de liens intermédiaires. Les graphes basés sur la distance euclidienne ou fonctionnelle (c.-à-d., modélisation des moindres coûts) sont aussi employés pour prévoir la connectivité du paysage (O’Brien et al. 2006; Minor & Urban 2008; Lookingbill et al. 2010). Dans ce cas, les parcelles distantes d’une valeur qui est en-dessous d’un certain seuil sont connectées alors que celles plus éloignées ne le sont pas.
Les patrons de connectivité identifiés dans la nature sont cependant généralement plus complexes que ceux d’un MPG ou d’un réseau basé sur une distance seuil (Rhodes et al. 2006; Almpanidou et al. 2014; Fox & Bellwood 2014). Par exemple, Fox & Bellwood (2014) ont montré que les réseaux spatiaux de plusieurs espèces de poissons présentaient à la fois des raccourcis et une connectivité fortement hétérogène entre les nœuds. Certains nœuds étaient très fortement connectés (c.-à-d., les pôles) alors que la plupart des autres nœuds avaient peu de liens (Fox & Bellwood 2014). Ces patrons de connectivité peuvent être représentés par des réseaux théoriques complexes (Fig. 1), tels que le réseau du petit monde (c.-à-d., présence de raccourcis) et le réseau libre-d’échelle (c.-à-d., distribution des degrés très hétérogène) (Watts & Strogatz 1998; Barabasi & Albert 1999; Newman 2003). Malgré la fréquente occurrence de patrons de connectivité complexes dans la nature, les réseaux simples sont souvent employés pour prédire la connectivité du paysage dans des systèmes réels (O’Brien et al. 2006; Treml et al. 2008; Lookingbill et al. 2010; Courbin et al. 2014), car peu d’outils sont disponibles pour construire des réseaux spatiaux complexes.
La structure d’un réseau a cependant des conséquences sur sa robustesse vis-à-vis des perturbations, telles que le retrait de nœuds ou de liens (Fig. 1) (Newman 2003; Reunanen et al. 2012). Par exemple, un réseau libre-d’échelle est robuste au retrait aléatoire de ses nœuds mais sensible au retrait ciblé de ses pôles, alors que le MPG est robuste au retrait de ses nœuds puisque de nombreux chemins alternatifs sont présents dans le réseau (Fig. 1) (Albert et al. 2000; Reunanen et al. 2012). Afin de pouvoir anticiper correctement l’effet d’une perturbation sur la connectivité du paysage, il est donc important d’identifier quel modèle de graphe reflète le mieux le patron de connectivité de l’espèce étudiée et d’utiliser ce modèle pour prévoir les déplacements potentiels des individus entre les parcelles de ressources.
Figure 1. Illustration de topologies d’un réseau simple, le graphe planaire minimum (MPG) et de deux réseaux complexes, le graphe libre-d’échelle et le graphe du petit monde, ainsi que de leur changement potentiel de connectivité suite au retrait complet d’un nœud. Dans un réseau spatial, chaque nœud représente une parcelle de ressources pour
l’espèce étudiée et les liens représentent des déplacements inter-parcelles observés ou potentiels. Un pôle (nœud rouge) est un nœud très connecté dans un réseau libre-d’échelle, comparativement aux autres nœuds du réseau qui sont, quant à eux, peu connectés. Un raccourci (lien bleu foncé) est un lien direct entre deux parcelles éloignées, qui croise d’autres liens et qui permet de traverser le réseau en passant par un faible nombre de liens intermédiaires. Les raccourcis sont inhérents au réseau du petit monde.
5.3 Relier la théorie des graphes à la répartition spatiale des populations
Prévoir la répartition spatiale d’une population à partir seulement d’un réseau peut cependant être risqué. En effet, les inférences sur l’utilisation de l’espace à partir d’une analyse de réseau seule peuvent être faussées, puisque la dynamique temporelle des déplacements (c.-à-d., associée au temps de résidence dans les parcelles) est absente avec une telle approche (Stehfest et al. 2015). Par exemple, Stehfest et al. (2015) ont démontré qu’un réseau seul ne permettait pas d’identifier correctement les aires préférées par les requins plat-nez (Notorynchus cepedianus), car la dynamique temporelle des déplacements