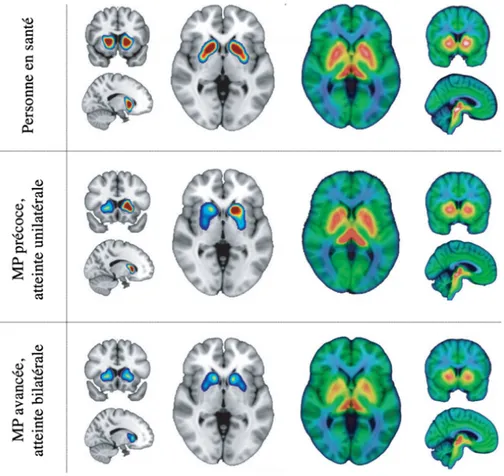
Études spectroscopiques de la structure, de l'auto-association et de la topologie membranaire du peptide amyloïde a-synucléine 71-82
281
0
0
Texte intégral
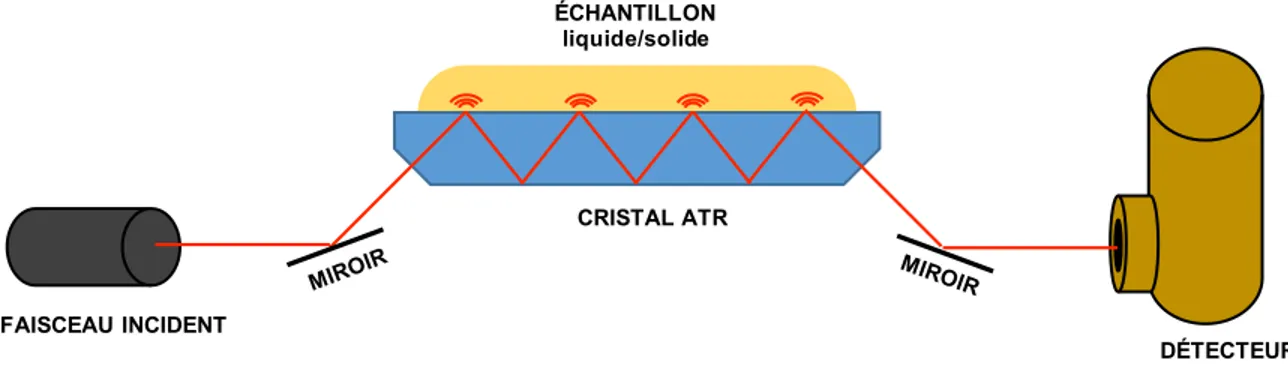
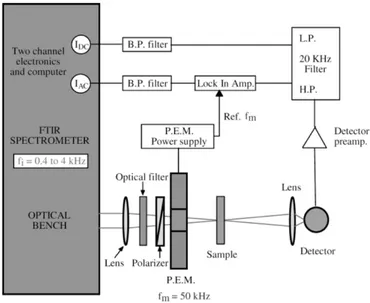
Figure



+7

Documents relatifs