Senga Patrick MUTONKOLE
Caractérisation chimique de bois de tiges et de
branches de Trema orientalis (L.) Blume et de
Leucaena leucocephala (Lam.) de Wit.
Mémoire présenté
à la Faculté des études supérieures et postdoctorales de l’Université Laval dans le cadre du programme de maîtrise en sciences forestières
pour l’obtention du grade de Maître ès sciences (M. Sc.)
DÉPARTEMENT DES SCIENCES DU BOIS ET DE LA FORET FACULTÉ DE FORESTERIE, DE GÉOGRAPHIE ET DE GÉOMATIQUE
UNIVERSITÉ LAVAL QUÉBEC
2013 © Senga Patrick MUTONKOLE, 2013
RÉSUMÉ
La caractérisation chimique des bois raméaux fragmentés (BRF) et du bois de tronc de Trema orientalis et de Leucaena leucocephala a révélé un plus fort degré de lignification dans les branches de Trema que dans le tronc, alors que le résultat inverse a été observé pour le Leucaena. De même des plus forts taux en cendres ont été trouvés dans les branches que dans le tronc, avec des valeurs globalement plus élevées pour le Trema. Par ailleurs, le ratio C/N déterminé par analyse élémentaire, a révélé des valeurs plus élevées pour le bois de tronc que pour les BRF des deux essences. En revanche, le ratio lignine/N des branches, naturellement plus faible, a conduit à la prédiction selon laquelle la décomposition de la matière ligno-cellulosique des branches serait plus rapide que celle du bois de tronc.De plus, les meilleurs ratios C/N et lignine/N ont été trouvés pour Leucaena. Enfin plusieurs composés identiques, dérivés de la lignine, ont été majoritairement identifiés par pyrolyse-GC-MS dans le bois de tronc comme dans les BRF.
Mots clés : Bois raméal fragmenté, lignine, Trema orientalis, Leucaena leucocephala, ratio C : N, ratio lignine : N, composition chimique, composition élémentaire, pyrolyse-GC-MS.
ABSTRACT
The chemical characterization of ramial chipped wood (RCW) and of the T. orientalis and the L. leucocephala stem woods has shown a higher degree of lignification in the branches of Trema than in the stem woods whereas reverse results have been found for the Leucaena.
Besides, higher rates in ashes have been found in the branches than in the trunk, with values globally higher for Trema. The C/N ratio, determined by elementary analysis, has revealed higher values for stem wood than for the RCW of the two species.
In contrast, the lignin/N ratio of the branches, naturally low, for the two species, has led to the prediction stating that the decomposition of the lignin-cellulosic matter of the branches would be faster than that of the stem wood. Besides, best ratio C/N and lignin/N have been found for Leucaena. At last, several identical compounds, derivatives of the lignin, have been mostly identified in the stem wood as in the RCW.
Keywords: Ramial chipped wood, lignin, Trema orientalis, Leucaena leucocephala, ratio C: N, ratio lignin: N, chemical composition, elementary composition, pyrolyse-GC-MS.
AVANT-PROPOS
Je tiens de prime à bord à remercier ma directrice et mon co-directeur, Mme Tatjana Stevanovic J. et M. Damase Khasa, pour leur encadrement et pour les conseils judicieux qu’ils m’ont prodigués tout au long de ma Maitrise.
J’adresse mes remerciements à Monsieur Yves Bédard, pour son assistance technique sans laquelle la justesse des analyses chimiques serait compromise.
Mes remerciements s’adressent également à Mlle Maria Prado pour avoir réalisé une partie des analyses pendant mon absence de Québec.
J’exprime mes sincères remerciements à Monsieur Ronald Dumarsais pour sa disponibilité et son assistance pendant les laboratoires.
La soumission pour publication du seul article de ce travail n’interviendra qu’après évaluation de ce dernier et pourrait subir quelques modifications en vue de l’adapter aux règles du journal. De plus, notre participation en qualité d’auteur principal dans la préparation de cette étude a consisté à réaliser les opérations suivantes : laboratoires, traitement et analyse des données et rédaction.
REMERCIEMENTS
Je tiens à remercier toute l’équipe du projet FOGRN pour avoir conçu et mis en œuvre le programme de formation des gestionnaires et spécialistes en foresterie dans le bassin du Congo. Mais aussi pour leurs conseils combien précieux tout au long de ma Maitrise.
J’adresse également mes remerciements au gouvernement du Canada à travers l’Agence Canadienne de Développement International (ACDI) pour son concours financier et pour toutes les facilités administratives afin de rendre effective ma formation.
Enfin, je remercie ma femme et mes deux filles pour le sacrifice consacré tout au long de ma Maitrise.
TABLE DES MATIERES
RÉSUMÉ ...i
ABSTRACT ... ii
AVANT-PROPOS ... iii
REMERCIEMENTS ...iv
TABLE DES MATIERES ... v
LISTE DES TABLEAUX ... vii
LISTE DES FIGURES ... viii
LISTE DES ANNEXES ...ix
LISTE DES ABREVIATIONS ... x
Chapitre I. Introduction générale ... 1
I.1 Mise en contexte ... 2
I.2. Revue de la littérature ... 7
I.2.1.1. T. orientalis var orientalis (Ulmaceae) ... 7
I.2.1.2. L. leucocephala (Mimosaceae) ... 8
I.2.2. Présentation de BRF ... 9
I.2.3. Historique de BRF ... 9
I.2.4. Composition de BRF ... 10
I.2.4.1. Nutriments ... 10
I.2.4.1.1. Concentrations en nutriments ... 10
I.2.4.1.2. Source de variation des teneurs en nutriments ... 111
I.2.4.2. Polyphénols ... 13
I.2.4.2.1. Définition et structure ... 13
I.2.4.2.2. Sources de variation de la concentration en polyphénols ... 14
I.2.4.3. Lignine ... 15
I.2.4.3.1. Introduction ... 15
I.2.4.3.2. Source de variation de la concentration en lignine ... 18
I.2.5. Expérimentations antérieures relatives à l’application des BRF ... 19
I.3. But et objectifs ... 22
Chapitre II. Caracyérisation chimique du bois de tiges et de branches de Trema or ... 23
RÉSUMÉ ... 24
II.1. Introduction ... 25
II.2. Matériel et méthodes ... 27
II.2.1. Description du site d’étude ... 27
II.2.2. Production du broyat ... 28
II.2.3. Extraction du broyat ... 29
II.2.4. Détermination de la lignine ... 30
II.2.5. Détermination en composants organiques par Py-GC-MS ... 31
II.2.6. Détermination de la cellulose ... 32
II.2.7. Détermination des pentosanes ... 32
II.2.8. Détermination de la composition élémentaire ... 33
II. 2.8.2 Composition élémentaire des échantillons des bois et des branches ... 34
II.2.9. Détermination des cendres ... 34
II.3. Résultats ... 35
II.3.1. Humidité du bois ... 35
II.3.2. Composition chimique : Teneurs en lignine, pentosanes, cellulose, extraits et cendres ... 35
II.3.3. Composition chimique élémentaire du bois de tiges et de branches de T. orientalis et L. leucocephala ... 38
II.3.4. Ratio lignine: N ... 40
II.3.5. Produits de pyrolyse-GC-MS ... 40
II.4. Discussion ... 43
II.4.1. Humidité ... 43
II.4.2. Teneurs en lignine ... 43
II.4.3. teneurs en pentosanes ... 45
II.4.4. Teneurs en cellulose ... 46
II.4.5. Composition chimique élémentaire ... 47
II.4.6. Ratio C:N ... 51
II.3.7. Ratio lignine :N ... 52
II.4.8.Teneurs en extraits ... 53
II.4.9. Teneurs en cendres ... 54
II.4.10. Pyrolyse-GC-MS du bois de tiges et de branches ... 55
Conclusion ... 56
Liste des tableaux
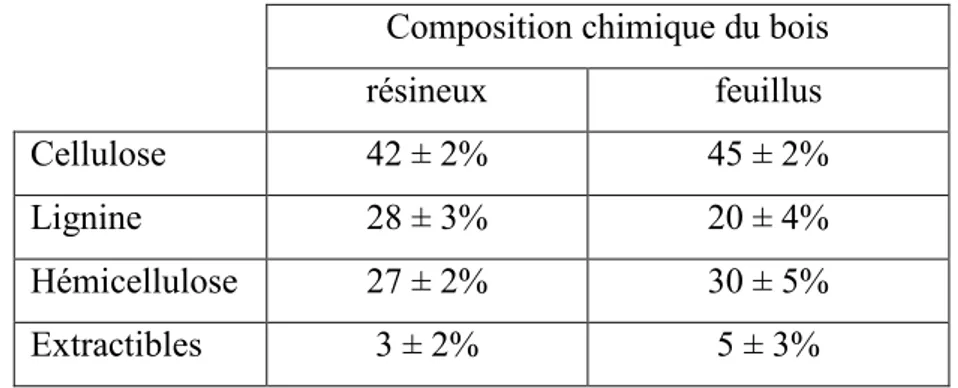
Tableau 1 : Composition chimique du bois (page 4)
Tableau 2 : Humidité du broyat extrait et non extrait (page 35)
Tableau 3 : Indices chimiques sur T. orientalis et L. leucocephala (page 36)
Tableau 4 : Teneurs du bois de tiges (T1-T5 et L1-L5) et de branches (TB et LB) en C, N, H, P, K, Ca, Mg, Na, Fe et Mn, et les ratios C/N du Trema et du
Leucaena (page 38)
Tableau 5 : Ratio lignine : N sur T. orientalis et L. leucocephala (page 40)
Tableau 6 : Composés organiques issus de la pyrolyse-GC-MS de bois de tiges et de branches de T. orientalis (page 41)
Tableau 7 : Composés organiques issus de la pyrolyse-GC-MS de bois de tiges et de branches L. leucocephala (page 42)
Liste des figures
Fig.1. : Planche du T. orientalis (page 7) Fig.2. : Planche du L. leucocephala (page 8)
Fig.3. : Structure probable de la lignine des feuillus (page 15) Fig.4. : Unités phénylpropane précurseurs de la lignine (page 16)
Liste des annexes
Fig.5. : Programme (Py-GC-MS) de T. orientalis (page 68) Fig.6. : Programme (Py-GC-MS) de L. leucocephala (page 68)
Liste des abréviations
ACDI : Agence Canadienne de Développement International BRFs : Bois Raméaux Fragmentés
C : Carbone
C/N : Rapport Carbone - Azote Ca : Calcium
CEC : Capacité d’Echange Cationique
CPPA-G-12 : Norme pour analyse des hémicelluloses D 1102 – 84 : Méthode dite standard pour cendres Fe : Fer
FOGRN : Formation en Gestion des Ressources Naturelles dans le Bassin du Fleuve Congo
G : Guaïacyle H : Hydrogène K : Potassium
LAEUM : Laboratoire d’Analyse Élémentaire de l’Université de Montréal
MEB-DEX : Microscopie Electronique à Balayage à Dispersion d’Energie de Rayons X Mg : Magnésium Mn : Manganèse N : Azote Na : Sodium P : Phosphore pH : Potentiel d’Hydrogène
Py-GC/MS : Pyrolyse combinée à la Chromatographie en Phase Gazeuse couplée à la Spectrométrie de Masse
RCW : Ramial Chipped Wood
RDC : République Démocratique du Congo S/G : Rapport Syringyle - Guaïacyle SOM : Matière Organique du Sol TCS : Technique Culturale Simplifiée
I.1 Mise en contexte
Dans de nombreux pays en développement, et sous l’emprise de la pauvreté, la manière d’utiliser les ressources de base comme les terres, les eaux, les forêts, la faune se soldent par des dégradations, souvent irréversibles (Maldague, 2010). En Afrique tropicale et plus particulièrement dans les zones péri-urbaines, la pression agricole et sylvopastorale sur les jachères se traduit par une forte diminution de leurs surfaces, de leurs durées mais aussi de leurs ressources pastorales et ligneuses (Masse et al., 1998 cité par Harmand et Ballé, 2001). Cela se révèle dans l’incapacité pour les jachères naturelles de jouer leur rôle de restauration de la fertilité physico-chimique et biologique des sols (Flore et al., 1993 cité par Harmand et Ballé, 2001). En conséquence, la situation alimentaire actuelle de maints pays en développement devient donc source de fortes inquiétudes.
Pour augmenter la production alimentaire dans ces pays, il devient indispensable de faire converger toutes les ressources vers l’amélioration de la productivité des petites exploitations familiales afin qu’elles puissent produire suffisamment de vivres, de combustibles et d’emplois et ainsi réduire l’exode vers les villes (Dalzell et al., 1988 ; Bekunda et al., 2010). Or, l’une des contraintes importantes à l’amélioration de la production agricole est le faible degré de fertilité naturelle des sols tropicaux. Dans la zone tropicale humide, les sols souvent profonds et de nature ferralitique présentent des bonnes caractéristiques physiques (structure, porosité, drainage), mais des caractéristiques chimiques médiocres : capacité d’échange cationique faible (5-16 méq/100g), désaturation, acidité et toxicité dues à l’aluminium et au manganèse (Levang et al., 1997). En effet, les terres affectées aux cultures vivrières pluviales sont généralement celles qui sont les plus pauvres, marginales, superficielles, arides et situées sur les versants des collines. Ce sont souvent des terres qui ont une faible teneur en matière organique (Gonzalez, 1992). Selon Eswaran et al. (1997) cités par Bekunda et al. (2010), 55% de la superficie de terres en Afrique est inadéquate à l’agriculture durable. Même les sols de haute à moyenne (16 – 13 %) qualité offrent une opportunité limitée pour l’agriculture commerciale hautement productive.
gouverné par la concentration d’azote (N) interne (élément essentiel le plus déterminant) à condition que les facteurs environnementaux non nutritionnels ne soient pas limitants (Van Den Driessche, 1990). Malheureusement en conditions tropicales, l’azote fait souvent défaut dans le sol. Pour pallier cette déficience, on procède soit par l’apport – coûteux – d’engrais chimiques en fonction de sa disponibilité et son accessibilité, soit encore par application de la matière organique directement ou transformée. Mais la matière organique peut quelques fois manquer en raison de la faible productivité de la biomasse dans certaines régions, du feu de brousse annuel dans d’autres.
Les pays en développement ont un besoin pressant d’accroître leur production alimentaire afin de réduire la pénurie. Cette augmentation devrait se baser, entre autres, sur l’amélioration et l’entretien de la fertilité du sol, en renforçant la stabilité structurale à long-terme et la rétention d’eau du sol, et en fournissant davantage aux plantes les éléments nutritifs, surtout sous la forme organique (Vandermeer et al., 1998 ; Bekunda et al., 2010). L’une des voies préconisées est d’utiliser de manière efficiente les ressources locales, en envisageant d’une part la protection des sols contre l'exposition prolongée au rayonnement solaire – afin de créer un microclimat favorable – et, occasionnellement, les fortes précipitations et, d’autre part, une augmentation du taux de la matière organique en vue de préserver les éléments nutritifs pour les plantes et la structure du sol. De cette façon, la fraction organique du sol ne représente pas seulement une réserve importante en éléments nutritifs, mais elle constitue aussi un facteur bénéfique pour : faciliter le labour naturel par la macrofaune (amélioration de la structure, par exemple : les sols lourds deviennent plus granulés) ; augmenter la capacité de rétention de l’eau (les plantes résistent mieux aux périodes sèches) et ; faciliter la reprise de la fertilité naturelle du sol (amélioration de l’activité microbienne) (Gonzalez, 1992). Si l’on réussit cette tâche, les besoins en engrais minéraux importés de l’extérieur de la communauté et, spécialement, de l’étranger, seront réduits. Ce qui aura des effets bénéfiques sur la balance des paiements du pays et atténuera l’urgence de disposer d’un système de transport et de distribution adéquat (Dalzell et al., 1988).
L’apport de la matière organique au sol peut s’opérer sous différentes formulations : les déjections de bétail directement déposées au sol ou après collecte, traitement et
application ; les résidus de culture utilisés in situ ou transférés sur d’autres surfaces ; le compost (Bekunda et al., 2010), etc. Une fois au sol, une partie des constituants organiques (acides aminés, sucres, vitamines,...) est rapidement utilisée par les organismes comme source d'énergie, alors que la lignine (molécule très énergétique mais difficile d'accès car possédant plusieurs noyaux aromatiques) jouerait un rôle déterminant dans la formation des substances humiques (acides humiques, fulviques) (Stevanovic et Perrin, 2009 ; Lemieux, 1995). Lemieux (1995) a révélé que la biotransformation de la lignine n'est possible qu'en présence des champignons du groupe des basidiomycètes, spécifiquement ceux de la pourriture blanche. De plus, ils sont les seuls capables de produire les enzymes nécessaires à leur transformation (Zongo, 2009)
Cependant, la proportion chimique dans les tissus végétaux (tableau 1) est un facteur important dans le choix de la ressource à appliquer au sol. En outre, le rapport C/N des tissus végétaux détermine le taux de libération des nutriments au cours de la minéralisation dans le sol (Bekunda et al., 2010 ; Rana et al., 2009). A cet effet, l’intérêt des BRF réside dans leur composition chimique hautement diversifiée en molécules complexes : sucres, acides aminés, vitamines et protéines en plus des minéraux et des constituants structuraux du bois (Stevanovic, 2006).
Tableau 1. Composition chimique du bois, d’après (Stevanovic et Pérrin, 2009) Composition chimique du bois
résineux feuillus Cellulose 42 ± 2% 45 ± 2% Lignine 28 ± 3% 20 ± 4% Hémicellulose 27 ± 2% 30 ± 5% Extractibles 3 ± 2% 5 ± 3%
La décomposition des BRF s’échelonnerait sur une durée plus longue en raison de leur teneur élevée en matière ligneuse difficilement dégradable (Lalande et al., 1998 ; N’Dayegamiye et Angers, 1993). Ce processus est encore rendu plus difficile par l’interpénétration étroite, des trois polymères structuraux (lignines, hémicelluloses et cellulose), dans les parois cellulaires, où les lignines forment des vraies liaisons
covalentes avec les hémicelluloses (Stevanovic et Perrin, 2009 ; Stevanovic, 2006). Par ailleurs, Otrysko et Pagé (2001) ont établi une relation étroite entre le type de matière organique apportée au sol et les organismes du sol. La source de carbone, les teneurs en nutriments, la présence des métabolites secondaires et l’arrangement physique de la matière organique influencent le développement des communautés microbiennes du sol (Heal et Dighton, 1985 ; Swift et al., 1979 cités par Otrysko et Pagé, 2001). Si l’infection de la matière organique se fait par les bactéries ou actinomycètes au départ, les basidiomycètes seront repoussés et la dépolymérisation de la lignine sera aléatoire ou inhibée (Lemieux, 1998). De plus, si les BRF ne sont pas en contact intime avec le sol, la colonisation de la matière organique se fera normalement et les BRF seront métabolisés uniquement au profit des colonisateurs mais au détriment du sol (Chervonyj, 1998). Une fois le métabolisme primaire terminé, il s’en suit une multiplication abondante de la mésofaune avec BRF (jusqu’à 23 fois), en particulier les acariens et les collemboles (fongivores et leurs prédateurs), avec des variations sensibles selon les essences utilisées, la taille et le diamètre des rameaux, l’apport éventuel de N, etc. (Larochelle, 1993 et 1994 ; Wiggins et al., 1979). La phase de dégradation ainsi installée correspond à ce qu’on qualifierait de métabolisme secondaire. Cependant, plusieurs facteurs tels que l’essence forestière utilisée et les modalités d’apport (dose, périodicité, dimension des fragments, etc.) peuvent limiter les effets négatifs que génèrent l’enfouissement des BRF (Barthès et al., 2010). Stevanovic (2006) a estimé que les fragments des BRF devraient mesurer 5 - 10 cm de longueur qu’on étalerait en une couche de 1-3 cm sur le sol, ce qui correspond à 100 - 300 m3/ha. De plus, l’application devrait être opérée avant la saison de pluies. Par ailleurs, 3 ans d’intervalle suffisent pour chaque application, à raison de 25 à 50 m3/ha, afin d’entretenir l’effet BRF (Lemieux, 2005).
De toute évidence, le processus entropique établi est d'origine forestière et n'a rien de commun avec les techniques enthalpiques actuelles qui sont le propre de l'agriculture (Lemieux, 1995). En effet, la résilience de la forêt est assurée par des mécanismes de maintenance biologique alors que l'agriculture est très ouverte et sa stabilité dépend du mode de gestion du sol et notamment de la compensation des pertes en minéraux et en carbone (C) (Barthès et al., 2010). De ce fait, l'utilisation des BRF permettra donc de provoquer et de maintenir des sols «forestiers» en milieux agricoles et d'en contrôler la
fertilité et la structure sur de longues périodes: ce qu’on appelle «l'agriculture forestière sans arbres» ou la véritable agroforesterie, la base même du développement durable tant recherché et avec justesse (Lemieux, 1995).
Aujourd’hui, au regard des résultats généralement meilleurs que fournit l’amendement de sol avec des BRF et la nécessité de produire sainement sans à devoir dépenser énormément d’énergie (la production des BRF requiert tout de même une dépense d’énergie), la promotion d’une technique culturale simplifiée (TCS) s’avère donc indispensable. C’est ainsi que nous nous proposons, dans le cadre de cette étude préliminaire, de caractériser chimiquement les BRFs en vue de leur application future in situ et d’en tester les effets sur leur potentiel d’amélioration du rendement cultural ainsi que leur capacité de séquestration du carbone dans le sol.
L’utilisation des BRF nécessite cependant la caractérisation chimique de ces derniers afin d’en prévoir le comportement, dont la vitesse de décomposition, sur le sol. C’est à ce thème principal que nous tenterons de répondre dans cette étude. Pour ce faire, deux essences tropicales (Trema orientalis (L.) Blume et Leucaena leucocephala (Lam.) de Wit.) ont été sélectionnées en vue de caractériser chimiquement leurs BRF, lesquels seront comparés à la fois entre eux et à la nature chimique de leur bois de tronc. Nous chercherons :
- laquelle parmi les deux essences produirait les meilleurs indices chimiques (proportions en lignine, cellulose, hémicelluloses, en minéraux) à partir desquels on pourrait dégager des meilleurs indices de décomposition dans le sol et/ou en laboratoire ?
- les contrastes pouvant exister entre une légumineuse et un non fixateur de N.
I.2. Revue de la littérature
I.2.1. Description botanique de T. orientalis et L. leucocephala
I.2.1.1. T. orientalis var orientalis (Ulmaceae)
Plusieurs espèces du genre Trema ont été identifiées précédemment en Afrique mais la grande variation morphologique apparemment indépendante des facteurs environnementaux fait en sorte qu’il y ait reconnaissance aujourd’hui de seulement une espèce sur le continent : Trema guinensis (syn. T. orientalis) (Anonyme, 1998).
La recherche expérimentale entreprise à ce jour sur cet arbuste a permis sa caractérisation chimique en vue d’en isoler les principes actifs et d’en quantifier l’efficacité médicinale (Anonyme, 1998). Ainsi, des composés comme lupéol, xanthones, décussatin, epicatechin,… (Tchamo et al., 2001 ; Kuo et al., 2007) ont été isolés. Quant aux constituants structuraux du bois, de l’écorce et des racines, la littérature ne fournit pas assez de données sur le sujet. A notre connaissance, seuls Jahan Arbuste à croissance rapide. Il atteint 4 à 5 m de haut en 5 - 6 années, ses branches sont lisses. Les feuilles de 14 cm de long sont asymétriques, alternant régulièrement sur la branche, les marges sont finement dentelées tout autour. Dans chaque aisselle, il y a un petit bouquet de fleurs qui persistent toute l’année. On le trouve dans la forêt secondaire à travers l’Asie et l’Afrique tropicale. On utilise les feuilles (comestibles), l’écorce et la tige dans le traitement de la dermite, des troubles mentaux, de la déshydratation, des caries dentaires, de l’asthme et de l’épilepsie. Les feuilles et les fruits sont mangés par le bétail. Il abrite les chenilles comestibles (Baumer, 1995).
et Mun (2007), cités par Rana et al. (2009), évoquent des teneurs en lignine du bois de Trema comprises entre 45 et 50 %.
Bien que citée comme espèce agroforestière (Nair, 1993 ; Arbonier, 2009), peu d’études ont cependant été consacrées sur l’aptitude du Trema à la combinaison avec les cultures. Toutefois Styger et al. (2009), Carrière (2005) l’ont signalé dans les systèmes agroforestiers traditionnels tels que les agroforêts, les jardins de case et les jachères améliorées où il participe à la restitution de la fertilité des sols, d’après Lubini, (2003).
Les noms vernaculaires sont : Mudianuni, Kidianuni, pigeon wood (Anglais).
I.2.1.2. L. leucocephala (Mimosaceae)
Beaucoup d’essais en ferme consacrés à la culture en couloirs avec Leucaena dans les rangées ont permis de mentionner l’intérêt de cette essence quant à la fertilisation des sols. A cet effet, Juo et Lal (1977) ont sou1igné que L. leucocephala est une essence de jachère à régénérescence spontanée qui se révèle particulièrement efficace pour enrichir le sol en matières organiques et en cations échangeables. Par ailleurs, Kang et al. (1981) et Kang et Duguma (1985) ont montré que L. leucocephala plantée en haies pour les besoins de la culture en couloirs, libérait de fortes quantités de N et que plus la rangée était étroite, plus la quantité d'azote était importante. Ngambeki (1985) a également
L. leucocephala est une plante fourragère à croissance rapide. Elle peut atteindre 7 m de hauteur sur sol à pH 5,8 une année après le semis (Kamabu et Lejoly, 1994). Les feuilles et les fruits commençant à former les graines peuvent être consommés cuits. L’effet dépilatoire de l’absorption, bien connu chez les porcs et chez les chevaux, a sans doute un effet négatif sur la consommation par l’homme (Baumer, 1995 ; Mendieta-Araica et al., 2012). Fig.2 : Planche du Leucaena (www.tropicalforages.info)
mentionné que des économies non négligeables d'engrais azoté pouvaient être réalisées lorsqu'on pratiquait la culture en couloirs du maïs avec Leucaena.
Malgré sa faible capacité adaptative sur sol acide, L. leucocephala est considérée comme une essence prometteuse pour la pratique d’enrichissement de sols tropicaux. Cela est attesté par un nombre incalculable d’écrits sur le sujet.
I.2.2. Présentation de BRF
Les BRF correspondent au broyat (2-3 cm de diamètre) de bois provenant de branches non desséchées de diamètre inférieur à 7 cm (ou rameaux), avec ou sans feuilles/bourgeons. Il peut être utilisé en tant qu’amendement par paillage et/ou incorporation sur les premiers centimètres du sol (Davy, 2011 ; Dave of Darlington, 2005). L’incorporation superficielle de BRF permet d’activer son intégration au sol, mais nécessite par ailleurs un contrôle régulier des différents paramètres, afin d’éviter d’inverser l’attaque des lignines au profit des bactéries ou actinomycètes (Lemieux, 1998).
L’origine biologique des mécanismes en cause lors de son intégration au sol implique une grande influence de la température, plus la température sera élevée plus les bénéfices de la méthode seront rapidement acquis et perdus par absence d’applications nouvelles ultérieures (Lemieux, 1995).
I.2.3. Historique de BRF
La technique du paillage au BRF est née de la volonté de M. Guay, ministre adjoint au ministère des Terres et Forêts du Québec dans les années 1970, à valoriser les résidus d’élagage issus d’arbres situés sous les lignes à haute tension. Après broyage et application aux sols, les cultures ont connu des rendements accrus les années suivant l’épandage de BRF. On a observé par exemple une récolte de céréale sur la parcelle traitée atteindre plus de 170% de la récolte de la parcelle non traitée (Guay et al., 1981 et 1982). Ces résultats extraordinaires et inattendus furent le début au Canada, d’une
longue série d’expérimentations forestières et agricoles, menées par Gilles Lemieux de l’Université Laval. Depuis, ce dernier a tenté de diffuser la technique à travers le monde, jusqu’à sa disparition en 2009. Malgré les efforts de communication d’universitaires canadiens, le monde financier n’a guère accordé beaucoup d’intérêt quant à l’émergence de cette technique. Aujourd’hui encore cette technique est encore très peu connue en Afrique, où quelques pays (Burkina-Faso, Sénégal, Côte d’Ivoire, Madagascar) seulement ont lancé des recherches ainsi que des essais hors champ pour comprendre ses effets dans les circonstances locales.
I.2.4. Composition de BRF
Le bois juvénile est constitué chimiquement de cellulose, d'hémicelluloses, de lignines, de protéines, d'acides aminés, de sucres simples, ainsi que de métabolites secondaires comme les polyphénols, les alcaloïdes, les pigments et les régulateurs de croissance (Fengel et Wegener, 1984; Waring et Schlesinger, 1985; Lemieux, 1986; Haider, 1992). Il existe cependant des variations considérables quant à la composition chimique de BRF et sa concentration en nutriments par rapport au bois caulinaire (Miller, 1984; Lemieux, 1986).
I.2.4.1. Nutriments
I.2.4.1.1. Concentrations en nutriments
Les branches et rameaux représentent une faible proportion de la biomasse de l’arbre, ce qui renvoie à une faible fraction des nutriments dans ces différentes parties par rapport au contenu de l’arbre total. Néanmoins, le bois jeune, les racines, les branches et surtout l’écorce sont généralement plus riches en matières minérales que le fût (Istas et Raekelboom, 1952). En effet, les BRF contiennent en moyenne 0,46 à 1,15 % d’azote (N), 0,26 à 1,10 mg g–1 de phosphore (P), 1,40 à 5,30 mg g–1 de potassium (K), 3,00 à 13,99 mg g–1 de calcium (Ca) et 0,42 à 1,20 mg g–1 de magnésium (Mg) (Tremblay et Beauchamp, 1998). A l’opposé, le bois contient 20 à 25 % de Ca, 2 à 10 % de K, 0,5 à 1,5 % de P (www.medieco.info), 0,15 à 0,25 % de Mg et 1 à 1,1 % de N (Bernhard, 1970).
I.2.4.1.2. Source de variation des teneurs en nutriments
Les concentrations en nutriments, aussi bien dans le bois que dans l’écorce de l’arbre, décroissent de façon exponentielle avec l'accroissement du diamètre, exception faite pour le calcium contenu dans l'écorce et dont la concentration reste stable (Hendrickson, 1987; Hendrickson et al., 1987; O'Connell, 1997 ; Melesse et al., 2012). Cette diminution peut être liée au processus de conservation des nutriments au sein de l'arbre, une partie de ces derniers étant recyclés pour la fabrication de nouvelles pousses, entraînant ainsi une forte baisse de la teneur en nutriments dans le bois et l'écorce des plus grosses branches (Hendrickson, 1987). Par ailleurs, la teneur en nutriments des écorces est bien supérieure à celle du bois proprement dit (Dumon et Gélus, 1982). Dumon et Gélus (1982), Le Gall (2006), Jing-shuang et Jun-bao (2001) et Melesse et al. (2012) ont révélé des concentrations en nutriments bien supérieures dans les feuilles et fruits que dans les autres parties de l’arbre. On peut dans ce cas considérer l’ordre de variation des teneurs en N, K, et Mg selon : feuillage > écorce > bois. Le calcium, quant à lui, varie différemment : écorce > feuillage > bois (Abbott et Crossley, 1982; Hendrickson, 1987). À l’échelle de la forêt cela implique que les concentrations de nutriments dans les arbres diminuent avec l’âge de la placette et que les sols s’appauvrissent alors que la forêt vieillit (Augusto et al., 2000a; Trichet et al., 1999 ; Istas et al., 1954). Or, Ranger et al, (1996) ont trouvé que, quel que soit l'âge du peuplement, la concentration en nutriments suit toujours ce modèle : jeunes aiguilles > vieilles aiguilles > jeunes rameaux > vieilles branches > écorce de tronc > bois de tronc, sauf pour Ca, dont les concentrations sont plus élevées dans les organes plus âgés. D’autre part, les sels minéraux puisés à chaque saison dans le sol servent à la constitution des feuilles et des jeunes branches mais pas à celle du tronc qui est essentiellement constitué de bois mort. Il en résulte que la fertilité du sol influe sur les concentrations en nutriments dans les feuilles, les aiguilles ou les petites branches, mais pas ou peu dans le bois du tronc (Augusto et al., 2000a). Ainsi les arbres qui poussent sur des sols riches en azote peuvent présenter de fortes concentrations d’azote dans leurs feuilles (NEGTAP, 2001 ; Bégin et Doyon, 2010 ; Mendieta-Araica et al., 2012).
Les concentrations en nutriments varient aussi en fonction de l’essence d’arbre utilisée. Ainsi, elles sont plus élevées chez les feuillus que chez les résineux (van den Driessche,
1984 ; Barthès et al., 2010). En particulier, les feuillus ont une plus grande proportion du contenu total en nutriments (20 – 25 %) de l'arbre dans le feuillage que dans celui des résineux (8 - 10 %) (van den Driessche, 1984).
Il y a lieu de signaler que les teneurs en nutriments dans les rameaux varient selon les saisons, les variations étant les plus importantes au printemps et à l'automne (van den Driessche, 1984; Hendrickson, 1987). La variation de la teneur en nutriments pendant cette période est attribuée au phénomène de translocation rapide des rameaux vers les feuilles de feuillus, alors que chez les résineux ce phénomène est lent (Luxmoore et al., 1981). On observe de ce fait une plus grande concentration en nutriments dans les feuilles pendant cette période (Alban, l985). Par contre, les variations en nutriments sont minimales et les concentrations sont maximales dans les rameaux pendant l’automne et l’hiver (Hendrickson, 1987). C'est ce que l'on appelle bois dormant (Lemieux, 1990). Sous les tropiques, les concentrations en nutriments dans les tissus végétaux font aussi objet de variation en fonction des saisons. En effet, Melesse et al. (2012) observent des concentrations plus élevées en Ca et cuivre (Cu) dans les feuilles de Moringa oleifera pendant la saison sèche qu’en saison pluvieuse.
Le niveau de fertilité du sol est très déterminant quant à l’absorption des nutriments par les végétaux mais aussi il est évident que les concentrations en nutriments dans les tissus dépendent de cette fertilité (Le Gall, 2004 ; Dumon, 1982). A cet effet, Maliondo et al. (1990), Melesse et al. (2012) et Campo et Gallardo (2012) ont observé que diverses espèces croissant sur des sols riches ont de plus fortes concentrations en N, P, K, Ca et Mg dans les constituants aériens que les mêmes espèces croissant sur des sols pauvres. De plus, Melesse et al. (2012) évoquent l’existence d’une corrélation entre l’élévation et le niveau de concentrations en minéraux dans les feuilles et/ou les gousses vertes de M. oleifera et M. stenopetala. En général, les gousses vertes de M. stenopetala, à faible élévation (1100 m d’altitude), contenaient des concentrations plus grandes en minéraux que celles d’individus croissant à des niveaux moyens (1700 m). Il serait utile, spécialement pour les essences tropicales d’évoquer les relations qui existent entre la composition chimique des matières minérales du bois et la nature du sol (Istas et Raekelboom, 1952), parce que la nature acide de ce sol comporte deux effets importants pour les plantes : la solubilisation de l’aluminium (Al) et la perte de
nutriments cationiques dans les sols (Harter, 2007). En général sous des conditions naturelles, l’habilité des plantes à accumuler les éléments chimiques est fonction de la règle de biologie et de géochimie, alors qu’en milieu perturbé elle est régie par une balance dynamique (Jing-shuang et Jun-bao, 2001).
I.2.4.2. Polyphénols
I.2.4.2.1. Définition et structure
Les polyphénols sont des composés benzéniques contenant plusieurs hydroxyles phénoliques (Stevanovic et Perrin, 2009). Les polyphénols comprennent les phénols simples, avec un seul noyau benzénique (squelette C6 et C6-C1), les stilbènes avec deux
noyaux benzéniques (squelette C6-C2-C6) et des nombreux produits issus de la synthèse
des phénylpropanes (C6-C3) : les lignanes, les coumarines et les flavonïdes, qui sont à la
base des tanins composés (Stevanovic et Perrin, 2009). Il y a cependant des exceptions à cette définition. En effet, certains composés considérés comme extractibles sont des vrais polyphénols et s’incrustent partiellement à la structure des parois cellulaires. C’est le cas par exemple des tanins, capables de transformer les peaux d’animaux en cuir (Haslam, 1998). Cependant, les extractibles sont présents dans toute la structure poreuse du bois, à la fois dans les parois et dans les lumens ou encore dans les canaux, leur concentration restant toutefois très faible dans les cellules de rayons (Stevanovic et Perrin, 2009 ; Heng Yin et al., 2012 ; Dumon, 1982 ; Dumon et Gélus, 1982). De plus, les métabolites secondaires sont facilement extraits avec des solvants organiques ou l’eau, sans devoir procéder à des traitements sévères qui modifient chimiquement les constituants structuraux du bois (Stevanovic et Perrin, 2009).
Les polyphénols sont largement distribués dans tout le règne végétal et toutes les plantes possèdent une fraction phénolique (Tissaux, 1994). Ainsi, les extraits de différents bois et écorces sont toujours des mélanges complexes dépendant du solvant utilisé et de l’origine botanique de la plante (Heng Yin et al., 2012). On observe donc une variabilité morphologique. Il est cependant possible d’identifier le matériel végétal en effectuant une séparation chromatographique de ses extractibles et d’établir leur motif caractéristique pour une famille, un genre ou une espèce de bois (Stevanovic et Perrin,
2009). De plus, les extractibles contribuent de façon importante aux propriétés physiques et technologiques du bois. En effet, les propriétés comme l’odeur, l’acidité et la couleur sont liées directement aux extractibles (Moya et Calvo-Alvarado, 2012), ainsi que les propriétés technologiques comme la durabilité et le comportement du bois vis-à-vis de l’adhésion, de la finition, du séchage, de la mise en pâte et du blanchiment des pâtes (Lallier-Vergès et al., 2008 ; Dumon et Gélus, 1982 ; Dumon, 1982 ; Stevanovic et Perrin, 2009).
I.2.4.2.2. Sources de variation de la concentration en polyphénols
Les teneurs en extractibles sont dépendants de nombreux facteurs, dont l’élément anatomique considéré, la position dans l’arbre, le taux de croissance de l’arbre, l’origine géographique, la génétique et la saison (Stevanovic et Perrin, 2009). En effet, la teneur en extractibles augmente progressivement de la moelle vers la périphérie du bois de cœur, aussi bien pour les duramens foncés que pour les pâles. Par ailleurs, la faible teneur en extractibles dans le bois de cœur des arbres plus jeunes est souvent à la base de leur moindre durabilité et de leur sensibilité à la décoloration et à la pourriture.
Cependant, Shure et Wilson (1993) ont montré que les espèces pionnières à croissance rapide (Robinia, Liriodendron) possèdent des concentrations plus faibles en tannins hydrolysables et en polyphénols totaux, mais des concentrations plus importantes en tannins condensés que les espèces à croissance lente et tolérantes à l'ombre (Acer, Cornus).
Concernant le rapport entre la teneur en composés phénoliques et la fertilité du site de croissance, plusieurs auteurs (Yaakoubi et al., 2009 ; Roy et al., 2001 ; McKey et al., 1978) ont démontré l’existence de ce rapport. En forêt tropicale par exemple, McKey et al. (1978) ont trouvé que la teneur en polyphénols dans le feuillage était presque deux fois supérieure chez les arbres croissant sur des sols peu fertiles que chez ceux croissant sur des sols riches. De même, en forêt tempérée, Roy et al (2001) ont trouvé que les teneurs en composés phénoliques dans les feuilles d’érablière étaient significativement faibles dans les milieux xériques de deux sites en étude (Duchesnay et Tingwick). Une
autre étude (Snakkers et al., 2000, citée par Stevanovic et Perrin, 2009), sur le chêne sessile (Quercus petraea) a montré que les arbres de futaie ont moins d’extraits que les arbres de taillis sous futaie et que l’origine géographique n’a pas d’influence majeure sur la quantité d’extraits. En outre, Pot (2004) établit l’existence d’une influence génétique sur la quantité d’extractibles dans le bois de pin maritime (Pinus pinaster Ait.). Finalement, la concentration peut varier selon la quantité de radiation solaire, une quantité plus importante de polyphénols étant retrouvée chez les plantes croissant en conditions ensoleillées plutôt qu'en conditions ombragées (Waterrnan et Mole, 1994).
I.2.4.3. Lignine
I.2.4.3.1. Introduction
La lignine est une macromolécule amorphe et complexe, non linéaire, constituée d’un système aromatique et phénolique (Stevanovic et Perrin, 2009) (Fig.3.). La lignine, ou plutôt les lignines, car elles diffèrent d'un groupe systématique à l'autre, s’incrustent dans les parois cellulaires du bois et de l’écorce. De ce fait, elles font partie, ensemble avec la cellulose et les hémicelluloses des composés structuraux du bois. Alors que la cellulose et l’amidon sont présents chez toutes les plantes, même les moins évoluées, les lignines sont des constituants caractéristiques des plantes vasculaires ou trachéophytes (Stevanovic et Perrin, 2009).
Fig.3. : Structure probable de la lignine des feuillus (Istas et Raekelboom, 1952)
La lignine représente en réalité une famille des polyphénols (structuraux), polymères issus de la polymérisation déshydrogénative des trois alcools cinnamiques (précurseurs de la lignine) : alcool coumarilique, coniférilique et sinapilique (Stevanovic, 2006) (Fig.4.).
Fig.4. Unités phénylpropane précurseurs de la lignine d’après Stevanovic et Perrin (2009)
Il faut néanmoins retenir que la composition monomérique des lignines varie beaucoup suivant les espèces, les organes, les tissus et même les fractions de paroi cellulaire (Raven et al., 2007).
Les lignines sont classifiées en trois groupes majeurs, basés sur la distribution de leurs précurseurs. A cet effet, la lignine de résineux est une guaïacyle (G), homogène, issue de l’alcool coniférylique comme précurseur principal, riche en sous-structures condensées contenant des liaisons C-C, tandis que celle de feuillus est une lignine syringyle-guaïacyle (S/G), plus hétérogène mais moins condensée que celle de résineux (Stevanovic, 2006). Les différences de structures entre les deux types de lignines expliquent déjà les différences en abondances de la lignine dans les deux types de bois. Par contre, on observe chez les angiospermes herbacées un type différent de lignine caractérisé par un polymérisât d'alcool coniférylique, sinapique et p-coumarique (Erikson et al., 1990). Quel que soit le type de lignine dont il s’agit, il existe toujours une même sous-structure qui domine : c’est la sous- structure aryle éther, ou encore ß-O-4 qui représente la façon dont le 40 à 50 % d’unités phénylpropane de la lignine sont liées ensemble, peu important le bois dont provient la lignine (Stevanovic, 2006).
Cependant, les polyphénols et les substances humiques dans les sols forestiers sont reliés par leurs structures. Sur ce, la théorie de formation d’humus dans les sols forestiers à partir des polyphénols est bien connue. C’est ainsi que Stevanovic (2006) propose une hypothèse pour la biogenèse de l’humus à partir des BRF (c’est-à-dire à partir de lignines et autres polyphénols présents dans cette matière) : les substances humiques représentent une combinaison des biopolymères biodégradés. L’auteure considère que la progression de la biodégradation des polyphénols présents dans les BRF (les lignines et les polyphénols extractibles) dépend de leurs structures et de la nature des microbes présents dans le sol et sur les BRF au départ. En effet, les études de la biodégradation de la lignine ont démontré que seulement une petite portion de la lignine était dégradée et disponible sous forme de petites molécules, même après l’action puissante des champignons de la pourriture blanche. La majeure partie de la lignine sous forme macromoléculaire reste partiellement modifiée seulement (Tissaux, 2001). De ce fait, cette partie de la lignine préservée après la biodégradation, est liée à la matrice du sol et contribue à la formation d’humus par des réactions à la fois de
dégradation et de condensation (Stevanovic, 2006). En définitive, l’auteure suggère que les compositions élémentaires de deux substrats, des acides humiques et de la lignine de l’écorce, leurs contenus en groupements méthoxyles ainsi que le rapport C : N pour les BRF, les lignines, les tannins isolés et les acides humiques isolés des sols traités par les BRF devraient être comparés pour mieux comprendre le devenir des BRF dans les sols.
I.2.4.3.2. Source de variation de la concentration en lignine
La concentration en lignine est plus élevée chez les gymnospermes que chez les angiospermes (Stevanovic et Perrin, 2009 ; Eriksson et al., 1990). La nature de l’espèce constitue également un autre facteur de différenciation de la teneur en lignines des végétaux (Eriksson et al., 1990). Des différences sont aussi rencontrées lorsqu’on considère différentes parties d’un même arbre, elle est en effet moindre dans les branches que dans les rameaux (Lemieux, 1995, 1998, 2005 ; Stevanovic et Perrin, 2009 ; Stevanovic, 2006).
Cependant, Melesse et al. (2012), en comparant les extraits chimiques des gousses vertes et des feuilles de M. oleifera et M. stenopetala, ont révélé l’existence d’une forte corrélation entre l’élévation, la saison avec leurs concentrations en lignines. En effet des teneurs élevées en lignines étaient observées dans les gousses vertes de M. stenopetala à une faible élévation (1100 m d’altitude) par opposition à celles retrouvées à une moyenne élévation (1700 m). Alors que, les plus faibles concentrations en lignines étaient trouvées durant la saison sèche et à moyenne élévation dans les feuilles de M. oleifera.
Contrairement aux polyphénols, la concentration en lignine dans les tissus végétaux n'est pas corrélée à la fertilité du site et montre quelques variations au sein d'une même espèce (Muller et al., 1987). En effet, selon le type de paroi (xylème, phloème, fibres,…), la concentration en lignines est plus forte dans les vaisseaux que dans les fibres des angiospermes (Reis et al., 2006). Pour leur part, Mendieta-Araica et al. (2012) ont rapporté des teneurs en lignines, dans la fraction fine de M. oleifera, variant significativement lorsque le sol était amendé en N. Ces teneurs étaient plus élevées
lorsque la quantité de N passait de 0 à 261 kg N ha-1 an-1 avec une densité de 167,000 plants ha-1.
I.2.5. Expérimentations antérieures relatives à l’application des BRF
L’apport de BRF génère des effets positifs sur le rendement agricole, sauf pour la culture qui suit immédiatement un premier enfouissement en sol sableux sous climat tempéré (N’dayengamiye et Dubé, 1986). Ce faible rendement est expliqué par une immobilisation de N par la microflore et, peut néanmoins, être compensée par des apports de N minéral ou organique. Zongo (2009) considère que l’emploi des légumineuses est préférable dans le cas d’un amendement organique. D'après Larochelle (1994), une deuxième solution pour minimiser l'immobilisation de l'azote serait d'augmenter la taille des fragments et le diamètre des branches. Cependant, Beauchemin et al. (1990) notent que l'immobilisation ne persiste pas l'année suivant l'application des BRF. L’immobilisation de l’azote durerait environ un an suite à une incorporation au sol des BRF (Lalande et al. 1998), alors que dans le cas de la paille de blé elle ne durerait qu’environ 2 à 4 semaines (Zongo, 2009). Ceci serait probablement dû à une déminéralisation de l'azote du sol durant la deuxième année, ainsi qu'à un début de minéralisation nette des copeaux à la fin de cette période (N'Dayegamiye et Dube, 1986 ; Beauchemin et al., 1990). En revanche, le rendement des cultures suivantes est souvent supérieur à celui du témoin (Barthès et al., 2010). Pour sa part, Davy (2011) constate que la modalité BRF a produit 31,5 % moins de richesse au bout de quatre ans d’expérimentation, cette différence s’étant essentiellement créé la première année (faim de N).
En conditions tropicales, les BRF appliqués sur sols moins sableux ou en mulch (même sur sol sableux) ont un effet positif sur le rendement dès la première culture (Aman, 1996 ; Gomez, 2003). Par ailleurs, les apports des BRF, surtout en mulch, améliorent les propriétés physico-hydriques du sol : infiltration, humidité, porosité et stabilité structurales plus élevées,
compacité plus faible, température plus stable, érosion réduite (Lemieux, 2001). D’après Noël (2006), Davy (2011) et Javernaud (2012), l’application de BRF, à court terme, améliore significativement le contenu en eau du sol. En effet, Noël (2006) a
mesuré un accroissement de l’humidité des sols traités avec du BRF correspondant à 350 l/m3. Dans un premier temps, il semble que le bois se gorge d’eau, accroissant ainsi mécaniquement le stock d’eau du sol. Après 8 mois, cette capacité de stockage diminue, probablement suite à la dégradation du bois. Après un an et demi, l’humidité relative au témoin augmente de nouveau, peut-être suite à la capacité de rétention de l’humus formé. Globalement, sur les quatre années d’expérimentation, cette technique à permis d’économiser respectivement 29 et 26% d’eau (irrigation + précipitation) en plein champ et sous tunnel (Davy, 2011).
En zone tempérée, les BRF testés sur des sols à texture légère conduisent à l’immobilisation de N et P, surtout l’année de l’apport, alors que le pH et la CEC ne semblent pas affectés (Beauchemin et al., 1992a ; Larochelle, 1994 ; Lalande et al., 1998). Une étude révèle une diminution de 7 à 3 % et 63 à 8 %, respectivement en P et Ca du sol pendant que les concentrations en K et Mg augmentent à raison de 3 à 19 % et 7 à 8 % respectivement avec application des BRF (Otrysko et Pagé, 2001). En zone tropicale, les apports ligneux ou herbacés ont peu d’effet sur NO3 et Pass, surtout lorsque
les stocks (kg/ha) plutôt que les teneurs (g/kg) sont considérés. En revanche, le pH et la CEC tendent à être plus élevés après apport ligneux que sans apport organique ou même qu’après apport herbacé (Soumaré et al., 2002).
Les BRF déterminent une augmentation graduelle de la teneur en C du sol, faible l’année de l’apport (0 à 10 % en général), le plus souvent significative l’année suivante (10 à 40 %) et au-delà (20 à 70 % avec apports renouvelés) (N’Dayengamiye et Dubé, 1986 ; N’Dayegamiye et Angers, 1993 ; Lemieux et Germain, 2001 ; Lalande et al., 1998 ; Barthès et al., 2010). Par ailleurs, Gasser et al. (1995) et Tremblay et Beauchamp (1998) ont noté, à court terme, une augmentation significative du niveau de carbone organique du sol à la suite de l’application de BRF. De plus, l’augmentation du rapport C/N ne devient significative qu’à partir de la quatrième année (10 à 20 % avec apport renouvelé). L’humification des BRF est ainsi favorisée par des apports azotés simultanés. Il s’avère aussi que les quantités d’humus formés suite à l’apport des BRF sont bien supérieures aux apports d’autres techniques et amendements. Si on compare les BRF à des quantités équivalentes de fumier, alors qu’il faut un demi-siècle pour augmenter le taux d’humus de 1% avec du fumier, il ne faut que 10 ans avec les BRF
(Noël, 2006).
N'Dayegamiye et Angers (1993) ont noté que l'action des BRF à long terme porte majoritairement sur les fractions stables de la matière organique (fractions lourdes et humines), mais aussi sur la fraction légère, plus labile. Ce qui interagit significativement de manière positive sur la fertilité (Otrysko et Pagé, 2001) et à réduire le potentiel érosif du sol (Noël, 2006). En effet, les mesures réalisées par ce dernier ont révélé qu’en augmentant le taux d’humus de 1 %, en 17 ans, par de forts apports organiques, on a pu réduire les problèmes d’érosion à 25 % de ce qu’ils étaient au départ.
L’application de BRF favorise la réinstallation de l’activité biologique du sol. En effet d'après Tremblay et Beauchamp (1998), l'incorporation de BRF a stimulé temporairement le développement des populations de champignons alors que les populations de bactéries et d'actinomycètes sont demeurées stables. Par ailleurs, Larochelle (1994) a observé que la mésofaune est plus abondante avec BRF (jusqu’à 23 fois), en particulier les acariens et les collemboles (fongivores et leurs prédateurs), avec des variations sensibles selon les essences utilisées, le diamètre des rameaux, la taille des copeaux, l’apport éventuel de N, etc.
En définitive, les BRF améliorent à coup sûr la croissance des plantes, ce qui induit positivement le rendement cultural. A cet effet, Stevanovic (2006) évoque des rendements équivalents à 1000 % de masse de tomates au Sénégal, 400 % de masse de maïs en Côte d’Ivoire et en République Dominicaine, 300 % de masse de fraises au Canada et 45 % de masse de seigle en Ukraine. Zongo (2009) a enregistré également des rendements positifs du sorgho blanc qui sont passés de 0,47 t/ha pour les sites sans BRF contre 1,814 t/ha pour les sites avec BRF. Par contre, Davy (2011) a obtenu des laitues ‘Iceberg’ deux fois plus petites avec BRF que sans BRF à cause semble-t-il à un retard de 18 jours de la date d’entrée en production de la parcelle. Des améliorations peuvent également être observées sur les propriétés organoleptiques des fruits et des légumes cultivés sur des sols traités avec BRF (Lemieux, 2005). Il semble néanmoins que l’utilisation des BRF issus des légumineuses n’ait pas d’effet particulièrement favorable (Barthès et al., 2010). Leur intérêt réside tout de même dans l’apport supplétif de N afin d’inhiber la faim d’azote tel que suggéré par Zongo (2009).
I.3. But et objectifs
Le but de cette étude est de comparer les caractéristiques chimiques des BRF avec celles de bois de tiges entre deux essences tropicales (T. orientalis (L.) Blume et L. leucocephala (Lam.) de Wit.), en vue d’en sélectionner laquelle présenterait des meilleurs ratios lignine/N et C/N.
Les objectifs spécifiques poursuivis sont les suivants :
1. Analyser les composants chimiques majeurs, notamment la lignine de bois à l'état mature et juvénile pour chaque espèce ainsi que leur composition minérale (inorganique);
2. Comparer les résultats d’analyses chimiques entre espèces en vue d’identifier laquelle présenterait des ratios élevés (C/N ou lignine/N).
Le mémoire présente les parties suivantes :
Chapitre I : Introduction générale comprenant la mise en contexte, la revue de la littérature et le but et objectifs de l’étude.
Chapitre II : Caractérisation chimique de bois de tiges et de branches. Ce chapitre comprend un résumé, une introduction, le matériel et méthodes, les résultats, la discussion de résultats et la conclusion.
Chapitre 2 : Caractérisation chimique de bois de tiges et de
branches de Trema orientalis et Leucaena leucocephala
RÉSUMÉ
La caractérisation chimique des bois raméaux fragmentés (BRF) et du bois de tronc a révélé un fort degré de lignification dans les branches que dans le tronc chez Trema, alors que le résultat inverse a été observé pour le Leucaena. Mêmement, le contenu en cellulose est plus élevé dans les branches et plus faible au niveau du tronc contrairement chez Trema. Par ailleurs, les concentrations en cendres sont toujours plus élevées dans les branches que dans le tronc. La proportion en N dans les branches est nettement supérieure à celle du tronc pour les deux essences. Quant aux rapports C/N et lignine/N, les valeurs sont plus élevées dans le bois de tronc que celles des branches. De plus, les ratios du T. orientalis sont nettement plus élevés que ceux du L. leucocephala. En revanche, le ratio lignine: N des branches, naturellement plus faible, a conduit à la prédiction selon laquelle la décomposition de la matière ligno-cellulosique des branches serait plus rapide que celles du bois de tronc. Pour les deux espèces, l’analyse par pyrolyse-GC-MS du bois des branches et du bois de tiges a permis l’identification des composés dérivés majoritairement de la lignine, suivis par des polysaccharides. Ces résultats originaux démontrent l’intérêt de l’utilisation des BRF de ces deux essences comme amendements.
II.1. Introduction
Les problèmes majeurs auxquels l’agriculture moderne est confrontée actuellement sont la dégradation des sols et la pénurie de l’eau. Le recours aux engrais chimiques et autres fertilisants artificiels constitue évidemment une solution d’appoint mais occasionnent in fine la pollution des eaux par l’azote (Stevanovic, 2006). A cet effet, le changement du mode d’utilisation des terres voudrait que l’on réduise de manière significative l’usage des engrais chimiques dans les habitudes de culture. Une solution à long terme est offerte par la « technique de la forêt » sur les sols agricoles, par l’apport des bois raméaux fragmentés (BRF) afin de développer les sols fertiles comme le fait la forêt (Lemieux, 1995). De ce fait, plusieurs expériences relatives à l'application de BRF ont été réalisées en agriculture (N'Dayegamiye et Dubé, 1986 ; Beauchemin et al., 1990 ; N'Dayegamiye et Angers, 1993 ; Larochelle, 1994 ; Tremblay et Beauchamp, 1998 ; Tremblay, 1995 ; Domenech, 2011 ; Rey, 2009 ; Otrysko et Pagé, 2001 ; Javernaud, 2012 ; Davy, 2011) ainsi qu'en foresterie (St-Amand et Lauzon, 1995 ; Pettigrew, 1998), en vue de comprendre les effets de cette matière sur le sol et sur les végétaux.
Constitué chimiquement des molécules diverses (acides aminés, protéines, vitamines, sucres, nutriments essentiels, cellulose, hémicelluloses, lignine,…), les BRF représentent un matériau essentiel pour l’aggradation des sols dégradés. La lignine et les polyphénols sont les molécules à la base de la formation des substances humiques (Flaig, 1978 ; Stout et al., 1981 ; Haynes, 1986 ; Haider, 1994 ; Stevenson, 1994 ; Stevanovic, 2006). Cependant, la décomposition de la litière est conditionnée par un ensemble d'interactions complexes entre les organismes du sol, la température, l’humidité, la qualité du substrat et les propriétés physiques de la litière (Swift et al., 1979; Edmonds, 1987 ; Moore et al., 1999). De fait, la décomposition peut être mesurée en établissant un suivi de la perte de masse de la litière (Sokpon, 1995). Sous des conditions de macroclimat uniforme, le ratio C : N conditionne le taux de libération de N au cours de la minéralisation et, par conséquent, la vitesse de décomposition (Bekunda et al., 2010). De plus, la dégradation du substrat lignifié serait régulée par la perte de masse de la lignine. Or, la dégradation de la lignine est favorisée par des concentrations élevées en N et est limitée par des concentrations élevées en cellulose dans le matériau lignifié (Eriksson et al., 1990). Toutefois, la concentration initiale
faible en lignine influe négativement sur le démarrage de la décomposition de la matière ligneuse, elle se fait dans ce cas tardivement à cause de la disponibilisation d’une grande quantité de carbone par les microorganismes (Berg et McClaugherty, 1987). On note tout de même l’intérêt de mélanger plusieurs espèces dans les BRF car semble-t-il la décomposition de litière de bonne qualité stimulerait la décomposition de litières de moins bonne qualité (Tissaux, 2001).
Cependant, la difficulté majeure rencontrée lors de l’emploi de BRF est la faible productivité (surtout en première application), résultant de la forte immobilisation de N lors de sa décomposition (Davy, 2011 ; Domenech, 2011 ; Javernaud, 2012). Aussi, le P serait immobilisé pendant ce processus de décomposition (Tchervonyj, 1998). Toutefois, un apport azoté simultané (sous forme minéral ou organique) peut limiter ce phénomène d’immobilisation (N'Dayegamiye et Angers, 1993 ; Larochelle, 1994 ; Tremblay et Beauchamp, 1998 ; Zongo, 2009). Dans tous les cas, la nature de la matière ligneuse est d’une grande importance dans la mesure où elle conditionne son devenir dans le sol.
En conséquence, l’application des BRF valorise les résidus forestiers de par son apport de lignine d’origine forestière, qui interagit sur l’aggradation des sols. Ce procédé contribue en même temps à la préservation de l’environnement par recyclage des sous-produits et de résidus de l’exploitation forestière.
L’objectif de la présente étude consiste à caractériser les rameaux (< 7 cm de diamètre) de deux essences tropicales (Trema orientalis et Leucaena leucocephala) quant à leur composition chimique (nutriments, lignine, carbone, extractibles, cendres) afin de jeter les bases des expérimentations in vitro ainsi qu’in vivo, à venir, sur le suivi de la décomposition des BRF et leurs effets sur les cultures et le sol.
Les hypothèses de recherche sont :
Les ratios lignine/N et C/N des BRF diffèrent lorsqu’il s’agit d’une légumineuse ou d’une non légumineuse ;
La nature et la teneur en lignine des BRF diffèrent lorsqu’il s’agit d’une légumineuse ou d’une non légumineuse.
II.2. Matériel et méthodes
II.2.1. Description du site d’étude
La ville de Kinshasa, capitale de la République Démocratique du Congo (RDC), a à la fois le statut administratif de ville et de province. Située sur la rive sud du fleuve Congo au niveau du Pool Malebo; elle fait face à Brazzaville. Elle est la ville-province la plus peuplée du pays avec une population évaluée approximativement à 9 463 749 habitants, pour un taux d’accroissement de 5,3 % et une superficie de 10.725 Km2 (www.wikipedia.org).
La végétation de la ville a longtemps été dégradée et remplacée par une végétation éparse d’arbres fruitiers tels que : Mangifera indica (Mangier), Elaeis guinensis (Palmier), Persea americana (avocatier), Dacryodes edulis (safoutier), le plus souvent plantés dans les domaines d’habitation privés. Cependant, quelques reliques d’arbres forestiers peuvent également y être retrouvées (Milletia lorentii, Terminalia superba, T. catapa, Vernonia amygdalina, Trema orientalis, etc), ce qui témoigne du développement antérieur de forêt primaire voire secondaire sur le site. De plus, beaucoup d’espèces pionnières (Parinari excelsa, Panicum maximum, Tridax procumbens), envahissant souvent les espaces défrichés et transformés en milieu de production vivrière, n’évoluent guère en phase de succession secondaire à cause d’une forte pression anthropique dans les environs de la ville.
Le matériel géologique de la ville de Kinshasa est composé des formations meubles de recouvrement (alluvions, dépôts sablonneux d'âge pléistocène et pliocène) et quelques grès polymorphes et tendres de recouvrement, alors que le sol est dominé par une couverture sablonneuse de type Kalahari (Baert et al., 1991). Le climat de la ville de Kinshasa et ses environs appartient au climat tropical humide (Aw4) selon la
II.2.2. Production du broyat
Deux essences constituent la base de traitements : L. leucocephala et T. orientalis. Des rameaux et des disques du tronc ont été récoltés en janvier 2011 sur cinq individus de chaque espèce, lesquels étaient disséminés sur différents sites de la ville. Le diamètre maximal à la base des rameaux était de 6 cm pour le Leucaena et 5 cm pour le Trema, alors que celui du disque de tronc était de 22 cm pour le Leucaena et 19 cm pour le Trema. Le niveau de taille convenable des disques de tronc a été de 0,75 m au-dessus du sol (Kamabu et Lejoly, 1994). Le bois a été par la suite fragmenté dans une déchiqueteuse afin d’obtenir des broyats de petite taille. La fragmentation a concerné uniquement le bois après retrait de l’écorce en ce qui concerne le bois de tronc. Par ailleurs, toutes les branches choisies pour l’étude étaient considérées comme BRF à cause de leur diamètre qui était soit inférieur, soit égal à 7 cm. Les altitudes maximale et minimale des sites de récolte étaient 396 et 307 m, respectivement.
Les broyats ont été ensuite fragmentés à l’aide d’un broyeur à marteaux avec une grille assez large (3 à 4 mm d'ouverture). Le produit du broyage a été à nouveau broyé en utilisant une grille plus fine (1mm) et tamisé sur un tamis à secousses. Seule la fraction comprise entre 40 et 60 mesh a été conservée dans un sac de polythène qui a été soigneusement fermé aussitôt après le tamissage. Une quantité de 50 g de broyat tamisé était nécessaire pour analyser chimiquement tous les indices.
Avant de procéder aux analyses proprement dites, le prélèvement de l’humidité (moyenne de deux échantillons de 2 à 3 grammes), après stabilisation (8 jours) des échantillons, est donc indispensable en supposant qu’ils la conserveront durant toute la période desdites analyses (Sun et al., 2010).
Le test de Student a été fait sur les résultats d’analyse élémentaire du bois de tronc, le bois de branches ne faisant pas partie parce qu’étant analysé après constitution d’échantillons composites.
II.2.3. Extraction du broyat
L’extraction du broyat a été effectuée en deux étapes ; le broyat a d'abord été extrait au mélange éthanol anhydre-toluène puis, après séchage à l'air, à l'eau chaude (Stevanovic et Perrin, 2009). Dans chaque cas la détermination quantitative du pourcentage d'extractibles a été effectuée sur une fraction du broyat.
- L’extraction au mélange éthanol-toluène (mélange azéotropique de l’alcool éthylique anhydre et du toluène en proportions de 1 litre d’alcool éthylique anhydre et 427 ml de toluène) a été effectuée, de façon continue, dans un soxhlet et après pesée de 20 à 25 g de broyat dans un bécher (600 ml). Une fois terminée, le broyat a été séché à l’air et conservé dans un sac en polythène pour la détermination de l’humidité après stabilisation. L’évaporation du solvant contenu dans le ballon a été opérée à l'aide d’un rotavapor sous vide pour obtenir l'extrait sec. L’analyse s’est terminée par l’étuvage du ballon à 102°C pendant 12 heures, à l’issu duquel le pourcentage d'extractibles au mélange éthanol-toluène par rapport au bois initial a été déterminé.
- L’extraction à l'eau chaude a consisté à mettre environ 2 g de broyat, pesé à 0,1 mg, dans un ballon de 250 ml et à introduire 100 ml d'eau distillée dans un chauffe-ballon sous réfrigérant à reflux. Après 3 heures d'ébullition, le contenu du ballon a été filtré sur un creuset filtrant de porosité C, préalablement taré et lavé par la suite avec 1 litre d'eau chaude. Le creuset a été séché à l'étuve à 102oC pendant 24 heures, puis le broyat extrait a été pesé afin de déterminer le pourcentage d'extractibles à l'eau chaude par rapport au bois initial sec. L'extraction du restant de broyat a été séché seulement à l'air afin d’en déterminer, comme précédemment, l'humidité.