Développement d’une plateforme de production d’anticorps monoclonaux
Texte intégral
Figure
![Tableau 1 : Les dix molécules thérapeutiques les plus vendues dans le monde en 2013 [3] .](https://thumb-eu.123doks.com/thumbv2/123doknet/6996739.199091/9.892.105.810.177.756/tableau-molécules-thérapeutiques-vendues-monde.webp)
![Figure 1 : Schéma de la structure d’un anticorps humain [7].](https://thumb-eu.123doks.com/thumbv2/123doknet/6996739.199091/11.892.107.573.630.986/figure-schéma-la-structure-d-un-anticorps-humain.webp)
![Tableau 3 : Capacités de culture estimées pour la production d’anticorps monoclonaux dérivés de cellules de mammifère [4]](https://thumb-eu.123doks.com/thumbv2/123doknet/6996739.199091/14.892.103.791.314.522/tableau-capacités-estimées-production-anticorps-monoclonaux-dérivés-mammifère.webp)
![Tableau 4: Commercialisation d’anticorps produits à partir des différentes cellules hôtes [28]](https://thumb-eu.123doks.com/thumbv2/123doknet/6996739.199091/21.892.104.794.901.1059/tableau-commercialisation-anticorps-produits-partir-cellules-hôtes.webp)
![Figure 2: a : Profil classique de glycosylations effectuées par les cellules CHO ; b : Structure des principales glycosylations présentes sur les anticorps [20]](https://thumb-eu.123doks.com/thumbv2/123doknet/6996739.199091/23.892.98.819.270.547/classique-glycosylations-effectuées-structure-principales-glycosylations-présentes-anticorps.webp)

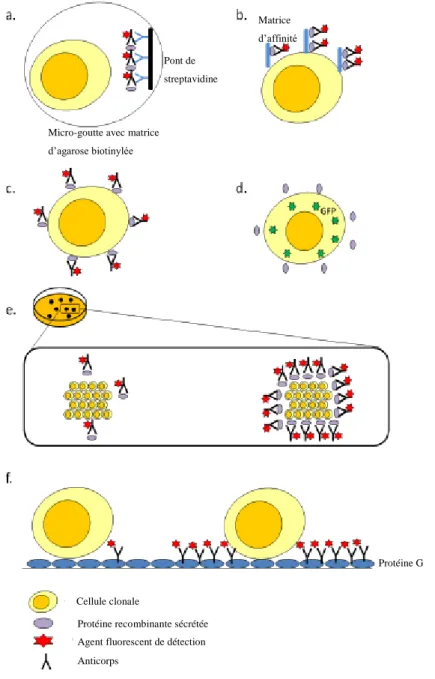
![Figure 6 : Illustration d’un processus typique de génération et sélection d’une lignée cellulaire de mammifère productrice d’une protéine recombinante [30]](https://thumb-eu.123doks.com/thumbv2/123doknet/6996739.199091/32.892.206.688.150.608/illustration-processus-génération-sélection-cellulaire-mammifère-productrice-recombinante.webp)

Documents relatifs
Aussi, plusieurs Žquipes INRA se sont associŽes ˆ la sociŽtŽ Biocytex afin de produire des anticorps monoclonaux spŽcifiques des iso- formes de myosine IIa et IIb chez le bovin,
Des années de recherche dans la physique des dispositifs à la base des semi-conducteurs ont conduit à la réalisation d’un modèle mathématique [6] .Ce modèle est capable d’opérer
La palmitoylation, qui consiste en J'ajout d'un acide palmitique sur les résidus cystéines de certaines protéines, est destinée à des protéines se retrouvant
Dès lors, pour les trois facteurs de transcription du groupe PEA3, nous avons procédé à une prédiction de la structure secondaire in silico en utilisant divers programmes de
Extending this idea to Seyfert 2 galaxies, we show here that two such active galactic nuclei (AGN) among the closest and brightest in the X-ray sky, NGC 1068 and NGC 4945, are
ﺎﻣ ﻰﻠﻋ ﺪﻤﺤﻣ ﻲﻟادﻮﺑ رﻮﺘﻛﺪﻟا ذﺎﺘﺳﻷا ﻞﺿﺎـﻔﻟا يذﺎﺘﺳأ ﻰﻟا ﺮﻜﺸﻟا ﻞﻳﺰﺠﺑ مﺪﻘﺗأ ﺎﻤﻛ فاﺮﺷﻻا ءﺎﻨﻋ ﻩﺪﺒﻜﺗ ﻰﻠﻋ و تﺎﻬﻴﺟﻮﺗ و ﺢﺋﺎﺼﻧ ﻦﻣ ﻲﻟ ﻪﻣﺪﻗ ـﻓ ءاﺰﺠﻟا ﺮﻴﺧ ﻲﻨﻋ ﻩﺮﺟأ ﻢﻬﻠﻟﺎ.. ﻰﻟا
Cinq ans après la fin d’une lutte collective longue et très médiatisée contre la fermeture d’une usine rurale de connectique du Sud-Ouest de la France, sous-traitante
