© Charlène Bouchard, 2019
L'impact de l'augmentation de l'apport d'aliments
solides avec différentes sources de protéines dans
l'élevage du veau de lait sur la croissance et la qualité
de la viande
Mémoire
Charlène Bouchard
Maîtrise en sciences animales - avec mémoire
Maître ès sciences (M. Sc.)
ii
Résumé
L’objectif de l’étude était d’évaluer les effets d’un remplacement complet du supplément protéique commercial à base de tourteau soya (CON) par un supplément de protéique cubé à base de gourgane (PFB) ou de la gourgane entière roulée (FB) sur la croissance et la qualité de la viande des veaux de lait. Les veaux (84.50± 1.70 kg) ont été assignés à quatre salles comprenant chacune 12 parcs. Pour cet essai, deux veaux par parc ont ensuite été utilisés et ont reçu l’un des trois traitements. Toutes les rations étaient isoprotéiques. Pour l’évaluation de la qualité de la viande, 30 muscles longissimus dorsi (LD) ont été prélevés du côté droit de la carcasse, et ce, pour chacun des traitements. Pour la croissance des animaux, le poids final des veaux était plus élevé pour le traitement avec la gourgane cubée par rapport au traitement avec la gourgane non transformée (P<0.05). Aucune différence significative n’a été remarquée pour le groupe témoin par rapport aux deux autres traitements. Le gain moyen quotidien (GMQ), mesuré entre les jours 48 et 146, était plus élevé avec l’ajout de la gourgane cubée (PFB) par rapport aux deux autres traitements (P< 0.05). De plus, aucune différence significative n’a été observée entre les différents traitements pour la concentration en hémoglobine. Tous les veaux avaient une concentration en hémoglobine de 94g/l au début de l’essai et une concentration de 88.4 ± 4.2g/l à la fin de l’essai, et ce pour chacun des traitements. Par ailleurs, pour les paramètres de qualité de la viande tels que le pH, la couleur (L*, a*, b*), la force de cisaillement, la longueur des sarcomères et le potentiel glycolytique, aucune différence significative n’a été observée entre les traitements. Il est possible de conclure que la gourgane entière ou cubée peut remplacer le supplément commercial à base de tourteau de soya sans affecter la croissance ni la qualité de la viande.
iii
Table des matières
Résumé ... ii
Liste des figures ... v
Introduction ... v
Chapitre 1 ... v
Listes des tableaux ... vi
Chapitre 1 ... vi
Chapitre 2 ... vii
Remerciements ... viii
Avant-Propos ... x
Introduction ... 1
Portrait de la production bovine au Québec ... 1
Chapitre 1 : Revue des travaux antérieurs ... 5
1.1 Élevage du veau de lait ... 5
1.1.1 Alimentation du veau ... 5
1.1.2 Élevage du veau en Europe comparé au Canada ... 7
1.2 Croissance du veau : Système digestif... 11
1.2.1 Développement du système digestif ... 11
1.2.2 La gouttière oesophagienne ... 12
1.2.3 Digestion du lait par le pré-ruminant ... 15
1.3 Muscle ... 16
1.3.1. Myogenèse ... 16
1.3.2 Structure générale du muscle ... 19
1.3.3. Transformation du muscle en viande ... 20
1.4. Qualité de la viande ... 24
1.4.1 Couleur ... 24
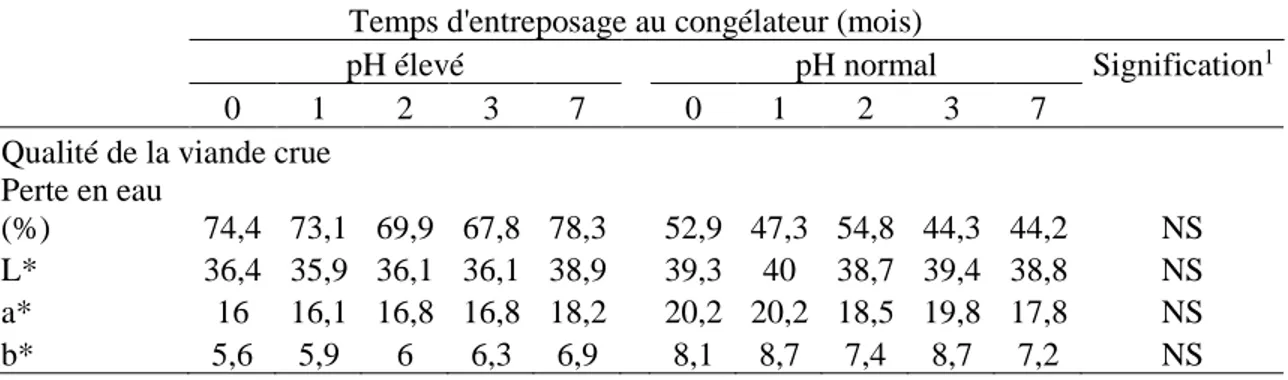
1.4.2. Potentiel glycolytique, pH et perte en eau ... 31
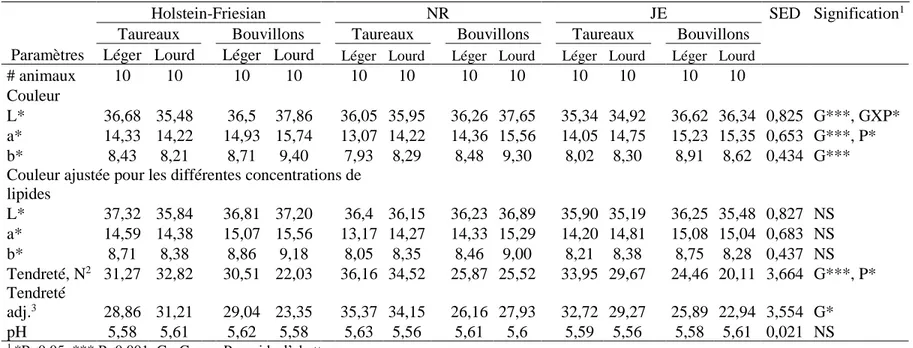
1.4.3. Tendreté ... 38
1.5. Alimentation ... 46
1.5.1 Les céréales ... 46
1.5.2 Gourgane (Vicia faba) ... 50
iv
1.6.1 Météorisation des veaux de lait ... 56
1.6.2 Anémie ... 58
1.6.3 Diarrhée néonatale ... 61
1.7 Conclusion ... 65
Chapitre 2: Article scientifique ... 66
2.1 Résumé ... 67
2.2. Abstract ... 68
2.3. Introduction ... 69
2.4. Materials and Methods ... 71
2.4.1. Animals ... 71
2.4.2. Diet ... 71
2.4.3. Growth and hemoglobin measurements ... 73
2.4.4. Colour, ultimate pH (pHu), samples and drip loss ... 73
2.4.5. Cooking loss and Shear force... 74
2.4.6. Sarcomere Length ... 74 2.4.7. Glycolytic Potential ... 75 2.4.8. Statistical Analysis ... 75 2.5. Results ... 76 2.6. Discussion ... 78 2.6.1. Growth performance ... 78 2.6.2. Health status ... 80 2.6.3. Meat quality ... 80 2.7. Conclusion ... 81 2.8. Acknowledgments ... 82 2.9. References ... 83 Conclusion générale ... 88
v
Liste des figures
Introduction
Figure 0-1 : Portrait global de la filière bovine ... 2
Chapitre 1
Figure 1-1 : Développement du système digestif du jeune ruminant ... 12Figure 1-2 : Variations du rythme cardiaque chez les veaux nourris à la chaudière et nourris à la tétine autour du repas de lait le matin ... 15
Figure 1-3 : Structure d’un muscle strié ... 20
Figure 1-4 : Évolution du pH musculaire après l'abattage de l'animal ... 22
Figure 1-5 : Relation entre le métabolisme énergétique musculaire et les caractéristiques musculaires déterminant la qualité de la viande ... 23
Figure 1-6 : Groupe hème de la myoglobine ... 25
Figure 1-7 : Les trois formes chimiques de la myoglobine ... 27
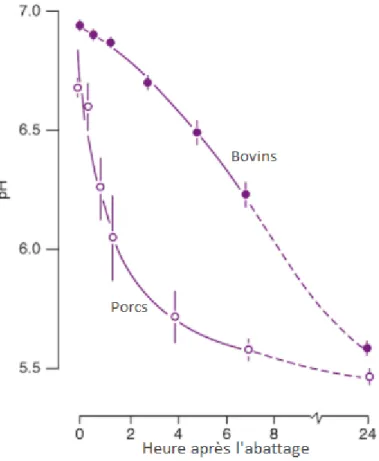
Figure 1-8 : Effet de l'heure après l'abattage sur le pH de différentes espèces ... 32
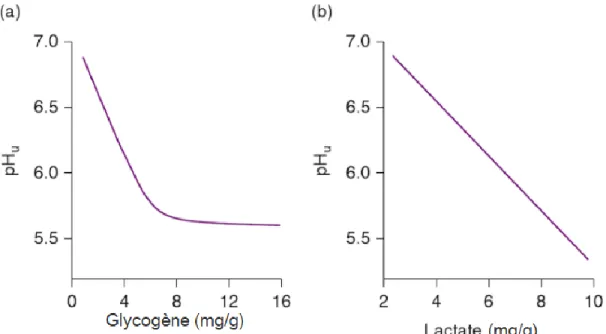
Figure 1-9 : a) L'effet de la concentration de glycogène sur le pH b) L’effet de la concentration de lactate sur le pH)………..33
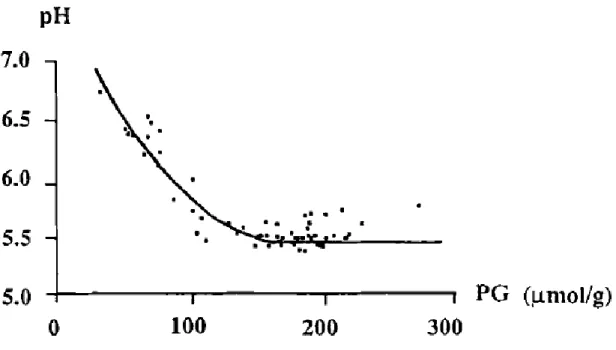
Figure 1-10 : L'effet du potentiel glycolytique sur le pH du muscle du veau ... 34
Figure 1-11 : Filament de myosine et d'actine ... 42
Figure 1-12 : Contraction des filaments d'actine et de myosine ... 43
Figure 1-13 : Effet de la température sur de la longueur des sarcomères pour le muscle LD et SM ... 44
Figure 1-14 : L'effet de la température sur la longueur des sarcomères et la perte en eau à la cuisson ... 45
Figure 1-15 : L'effet de la température à la cuisson sur la force de cisaillement ... 46
Figure 1-16 : Effet de la semaine sur l'évolution de l'hémoglobine dans le sang ... 60
vi
Listes des tableaux
Chapitre 1
Tableau 1-1 : Caractéristiques des principaux types de fibres du muscle squelettique ... 18 Tableau 1-2 : Valeur moyenne pour les concentrations plasmiques avant et après le transport ... 22 Tableau 1-3 : Effect du pH sur la qualité de la viande crue ... 28 Tableau 1-4 : Effet du temps d'entreposage au congélateur sur la qualité de la viande crue ... 28 Tableau 1-5 : Tendreté et couleur du muscle longissimus thoracis pour la race Holstein-Friesian, Norwegian Red x Holstein-Friesian (NR) et Jersey X Holstein-Friesian (JE) abattus à deux poids différents pour les taureaux et les taures ... 30 Tableau 1-6 : Caractéristique des carcasses normales et celles à couleur foncée (DFD) ... 35 Tableau 1-7 : Les effets du jeûne et du temps de transport sur le potentiel glycolytique, le pH et les paramètres de qualité de la viande ... 37 Tableau 1-8 : La moyenne (± écart-type) pour les paramètres sensoriels de la viande pour deux classes de pH ... 38 Tableau 1-9 : La valeur de la force de cisaillement (WBSF) et la perte en eau à la cuisson pour les muscles longissimus thoracis (LT), semitendinosus (ST), semimembranosus (SM), biceps ... 40 Tableau 1-10 : L’effet de la force de cisaillement mesurée pour le muscle longissimus lumborum et thoracis dans différents groupes d'animaux ... 40 Tableau 1-11 : Effet de la classe de l'animal sur les paramètres de qualité de la viande... 41 Tableau 1-12 : Propriété physico-chimique du muscle longissimus thoracis, % ... 48 Tableau 1-13 : Poids vif et gain moyen quotidien en fonction des différentes rations d'orge et de maïs ... 49 Tableau 1-14 : Paramètres de croissance et les charactéristiques de la carcasse sur des taureaux en fonction de différents ratios orge et maïs de leur ration ... 50 Tableau 1-15 : Profil en acides aminés du soya et de la gourgane ... 51 Tableau 1-16 : Ingrédient et composition chimique (%) des rations ... 52 Tableau 1-17 : L'effet de l'alimentation avec différentes protéines, le soya, la gourgane, le lupin et le pois sur les différents paramètres du muscle longissimus dorsi (LD) et semimembranosus (SM) ... 53 Tableau 1-18 : Effet de différentes rations avec des grains de légumes (féverole, pois, pois
chiche) et de soya dans les concentrés sur les paramètres de croissance et de qualité de la viande sur des agneaux ... 55 Tableau 1-19 : Composition chimique et digestibilité in vitro de la protéine brute et de la matière organique pour des cochons de la fève entière de la féverole, de sa fève sans coque et de sa coque seule ... 56
vii
Chapitre 2
Table 2-1: Composition and chemical composition of diets as feed ... 72 Table 2-2: Effect of faba bean (FB), pelleted faba bean (PFB) and commercial supplement (CON) in the concentrate on growth and slaughter parameters ... 76 Table 2-3: Effect of faba bean (FB), pelleted faba bean (PFB) and commercial supplement (CON) in the blood hemoglobin concentration in three different days ... 77 Table 2-4: Effect of faba bean (FB), pelleted faba bean (PFB) and soybean meal supplement (CON) in the concentrate on physical characteristics of the longissimus dorsi meat... 77
viii
Remerciements
La rédaction de ce mémoire signifie la réalisation d’un accomplissement personnel. Ce long cheminement fut parfois joyeux, parfois plus pénible, cela a sans aucun doute façonné la personne que je suis devenue et cela a permis de développer mon savoir-faire pour le milieu de la recherche. La réalisation de ce mémoire n’aurait pas eu lieu sans l’aide de ces nombreuses personnes et je tiens à leur rendre hommage.
Tout d’abord, je tiens à remercier le Dr. Dany Cinq-Mars, qui, à titre de directeur de mon mémoire, a su me transmettre ses valeurs et ses qualités professionnelles qui furent essentielles à la réussite de mon cheminement. Merci également pour sa disponibilité et sa patience qui furent essentielles pour mon cheminement de maîtrise.
Je veux également remercier Dr. Nadia Bergeron de chez Agrinova pour son temps, son expertise et de son aide tout au long de ma maîtrise. Elle m’a beaucoup conseillé et réorienté lors de la rédaction de ce mémoire. Pour moi, elle a joué le rôle de co-directrice et tous ses conseils ont été très appréciés. Je tiens à remercier également les employés de chez Agrinova : Annie Perron, Agr. et Alexandre Boilard pour leur aide lors de la réalisation de la phase animale de mon projet. Sans eux, les mesures à la ferme auraient été impossibles. Je tiens à dire merci à Agrinova qui sans eux mon projet de maîtrise n’aurait pas eu lieu.
Je remercie le Département de Sciences Animales de la Faculté d’agriculture, d’alimentation et de la consommation de l’Université Laval. Un merci particulier à Nancy Bolduc pour son aide lors des analyses en laboratoire.
Je remercie également le producteur de Prolacto, M. Jocelyn Fillion, d’avoir accepté la réalisation de mon projet sur sa ferme commerciale et de l’aide précieuse de son employé de ferme, Marie-Josée, qui m’a transmis toutes ses connaissances et son expertise dans l’élevage du veau de lait. Je remercie l’abattoir de Valleyfield et White Veal pour l’abattage et la découpe des veaux.
Je remercie également Agriculture et Agroalimentaire Canada, principalement Dr. Genevieve Drouin, pour son temps, son expertise et le transfert de son savoir-faire pour le protocole du potentiel glycolytique.
Je désire aussi remercier le Dr. Éric Pouliot pour son temps et le savoir-faire pour la réalisation des mesures de qualité de la viande.
ix
Je veux remercier Marie-Pierre Fortier du Centre du Développement du Porc du Québec (CDPQ) et son équipe pour son savoir-faire et son expertise sur les mesures de qualité de la viande en abattoir, pour son temps et le transfert de ses connaissances pour l’écriture de mon article scientifique.
Finalement, je tiens à remercier mes amis et ma famille qui m’ont encouragé tout au long de ma maîtrise. Je vous aime énormément!
Vous m’avez encouragée lors des moments les plus durs, vous m’avez poussée à ne jamais baisser les bras et vous avez cru en moi. Votre soutien a été d’une grande aide et c’est grâce à vous si j’ai persévéré et que j’ai eu le courage et la détermination d’atteindre mon objectif en finissant la rédaction de ce mémoire.
Ces deux ans de maîtrise n’ont pas toujours été faciles, mais c’est une expérience que je recommencerais sans faute. Je vous dis à tous un GROS MERCI!
x
Avant-Propos
Dans ce mémoire, le chapitre 2 est rédigé en anglais sous forme d’article scientifique dont je suis l’auteur principal. Les coauteurs sont le Dr. Dany Cinq-Mars et Nadia Bergeron qui ont participé à l’élaboration du dispositif expérimental et à la révision de l’article. Marie-Pierre Fortier a également participé à la révision de l’article et aux mesures de qualité de la viande. Le Dr. Éric Pouliot a aidé en apportant son savoir-faire pour l’analyse en laboratoire. L’article portant pour titre « Effect of increasing solid food intake with different protein sources on growth, carcass and meat quality in milk-fed calves » sera envoyé pour être publié dans la revue « Canadian Journal of Animal Science ».
1
Introduction
Portrait de la production bovine au Québec
Depuis une trentaine d’années, au Québec, la production bovine n’est ni en essor ni en régression, elle est stagnante (AAC, 2018a). Malgré la crise de l'ESB (encéphalopathie spongiforme bovine), aussi appelée ''crise de la vache folle'' en 2003, la consommation de bœuf par habitant n'a pas été affectée. Les mesures sanitaires ont été augmentées et ceci a eu pour effet de renforcer le lien de confiance entre les consommateurs et le système d'inspection (Forge et Fréchette, 2005; AAC, 2014). Ce qui a été le plus affecté par cette crise est le marché d'exportation, car plus de 60 pour cent de la production de bœuf canadien était exportée vers les États-Unis, le Mexique et le Japon (Forge et Fréchette, 2005; FPBQ, 2016b). Il n’y a pas eu d’autre crise de cette envergure depuis 2003. Par contre, les défis restent constants et les dernières années ont été difficiles pour le secteur bovin (FPBQ, 2016a). La production bovine est la quatrième production animale la plus importante au Québec, après la production laitière, la production porcine et la production avicole (FPBQ, 2016a). Les industries laitières et porcines sont prédominantes au Québec, tandis que la production bovine est prédominante en Alberta (Fournasier, 2007). À l'échelle canadienne, le Québec est un joueur modeste, puisque la production bovine représente 4% de la production nationale (Fournasier, 2007; FPBQ, 2016a). Sur le plan régional, la production bovine, secteur vache-veau, est localisée dans les régions périphériques du Québec, où la production fourragère est abondante comme l'Outaouais, l'Estrie, Chaudière-Appalaches et l'Abitibi-Témiscaminque (Fournasier, 2007; Thériault et Lamontagne, 2009). Par ailleurs, on compte actuellement près de 11 000 fermes bovines au Québec (FPBQ, 2016a). Plus précisément dans le secteur du veau lourd, le Québec est la province canadienne où l'on produit le plus de viande de veau avec près de 83% de la production nationale (FPBQ, 2016a; Ledoux, 1999).
La production bovine au Québec est divisée en deux filières soient : la filière bœuf et la filière veau (FPBQ,2016a) (Figure 1-1). Pour la filière bœuf, celle-ci rassemble les vaches de boucheries, les veaux d'embouche, les veaux semi-finis et les bouvillons d'abattage. De plus, il y a une sous-filière de vache de réformes qui proviennent des fermes de bovins de boucherie et des vaches réformées sur une ferme laitière. La filière bovine de réformes est aussi appelée bœuf commercial (FPBQ, 2016a). Le Québec produit environ 13% de ce bœuf (FPBQ, 2016a). Par la suite, on retrouve la filière veau qui rassemble les veaux indésirés sur les fermes laitières qui sont élevés de deux méthodes différentes : soit aux grains soit au lait (Figure 1-1).
2
Comme mentionné précédemment, l'élevage des veaux se divise en deux : l'élevage de veaux de grain et l'élevage de veaux de lait. La distinction principale entre les deux est leur mode d'alimentation, car l'un est nourri principalement avec des aliments solides, soit des céréales, et l'autre est nourri principalement de lait, qui est en fait un lactoremplaceur. Actuellement, le veau de lait reçoit, durant les 170 jours d’élevage, 2400 litres de lactoremplaceur et 50 kilogrammes d’aliments solides (Laplante, 2015). De nos jours, le veau de lait commence à recevoir du grain en petite quantité après 30 jours d’élevage (Laplante, 2015). Le veau de grain, quant à lui, reçoit 300 litres de lactoremplaceur et 750 kg d’aliment solides (Laplante, 2015). Pour le veau de grain, l’alimentation liquide avec le lactoremplaceur dure 65 jours (Laplante, 2015). Par la suite, il est Figure 0-1: Portrait global de la filière bovine, (FPBQ, 2016a)
3
nourri avec des aliments solides qui consistent en des céréales avec des concentrés protéiques, minérales et vitaminiques (Laplante, 2015).
L'élevage de veau de grain a connu un essor vers les années 1980, permettant de valoriser les veaux mâles laitiers (PBQ, 2016a). Il y a environ 200 fermes de cette production au Québec, principalement situées en Montérégie et au Centre-du-Québec (PBQ, 2016a). Une ferme moyenne de veaux de grain élève annuellement 700 veaux (PBQ, 2016a). Pour ce qui est de la production de veaux de lait, celle-ci a également connu un essor dans les années 1980. Avant cette date, les veaux laitiers mâles n'étaient pas valorisés, de même que les sous-produits laitiers comme la poudre de lait. Le Québec est le joueur dominant dans le Canada pour l'élevage de veaux de lait (PBQ, 2016b et FPBQ, 2016a). On y produit près de 80% de la production nationale de veaux de lait dont 50% est exportée (PBQ, 2016b). On retrouve environ 160 fermes de veaux de lait qui se situent dans les régions de la Montérégie, du Québec et du Centre-du-Québec (FBQ, 2016b). Une ferme moyenne au Québec élève autour de 700 veaux de lait annuellement (FBQ, 2016b; FTBQ, 2016). De plus, le nombre de veaux mis en marché par année pour les deux productions de veaux tourne autour de 70 000 veaux de grains et 134 000 veaux de lait, donc un total de 204 000 veaux élevés par année (Laplante, 2015).
En ce qui concerne l’exportation de viande de veau, celle-ci est calculée en viande rouge de veau et de bœuf. Les principaux importateurs sont les États-Unis, le Japon et Hong Kong (AAC, 2018b). Pour ce qui est de l’exportation de veaux vivants vers les États-Unis, il y a eu 5 036 veaux exportés en 2016 selon AAC (2017). Cela représente 0,65 pourcent des bovins exportés et 2,3 pour cent de la production totale de veaux. Le reste des veaux sont achetés par les abattoirs et les transformateurs et sont abattus au Canada (Laplante, 2015).
Les grosses entreprises dans le secteur du veau de lait sont : Délimax, Grober Nutrition et Pro-Lacto. Il y avait également Écolait, mais en novembre 2017, il a été acheté par Délimax pour ses activités en production et en transformation du veau au Québec. Ces entreprises achètent leurs veaux soit à l’encan ou directement à la ferme où ils peuvent faire affaire avec un courtier (Laplante, 2015).
C’est dans un contexte québécois que le présent travail a été réalisé. L’objectif de cette étude est d’évaluer l’effet du remplacement du complément protéique de la ferme composé de tourteau de soya par d’autres sources protéiques telles que la gourgane et la gourgane cubée sur la croissance et la qualité de la viande de veau de lait produit au Québec.
4
Dans le présent mémoire, la production du veau de lait au complet sera abordée jusqu’au produit fini de la viande. Les différentes sources de protéines ne devraient pas avoir d’effet sur la qualité de la viande, c’est ce qui sera vérifié.
5
Chapitre 1 : Revue des travaux antérieurs
1.1 Élevage du veau de lait
Comme mentionné précédemment, l’élevage de veau de lait est différent de l’élevage de veau de grain. La principale différence est au niveau de la consommation des aliments : lait versus concentré. Le veau de grain consomme 700kg d’aliments solides durant son élevage tandis que le veau de lait en consomme seulement 50kg (Laplante, 2015). Ce dernier va consommer plus de lactoremplaceur. Ce régime alimentaire différent a une répercussion sur la couleur de la viande du produit fini. Le veau de grain qui a une plus grande consommation de concentrés et par conséquent de fer durant son élevage, a une couleur de la viande plus foncée que celle du veau de lait qui en raison d’une alimentation faible en concentré et donc plus pauvre en fer, a une couleur de la viande plus pâle.
1.1.1 Alimentation du veau
Après le colostrum ingéré à la naissance, le veau doit recevoir un autre aliment lacté contenant principalement des protéines laitières, le lactosérum, jusqu'à ce qu'il atteigne un poids vif de plus de 200kg (Mornet et Espinasse, 1977; Diorio et al., 1999). Le veau reçoit deux types d'aliments lactés au cours de l’élevage : un lait de démarrage et un lait de finition. Le pourcentage de protéine contenu dans le lait de démarrage se situe entre 20% et 22% et le pourcentage de matières grasses entre 15% et 18% (Diorio et al., 1999). Le niveau de fer se situe entre 40 et 55 ppm (Diorio et al., 1999). Le lait de finition, quant à lui, contient généralement un taux de protéine plus faible et un taux de matières grasses plus élevé que le lait de démarrage (Diorio et al., 1999). Le lait de finition doit avoir un niveau de fer entre 8 et 11 ppm (Diorio et al., 1999). Dans un élevage de veau au Québec, il y a en moyenne 50 kg de poudre de lait de démarrage et 230 kg de poudre de lait de finition qui sont donnés au veau durant les 140 à 160 jours d'élevage. La diminution de la concentration en fer dans le lactoremplaceur est en lien avec la couleur de la viande désirée chez le veau lait. Si le veau ingère une concentration élevée de fer, la couleur de la viande devient plus foncée. Pour obtenir une couleur plus pâle, la quantité de fer donnée doit être contrôlée, c’est-à-dire que le minimum sera donné pour le développement du veau et pour que celui-ci ne soit pas carencé (Mornet et Espinasse, 1977). La couleur de la viande sera traitée à la section 1.4.1. Le lactoremplaceur est l'aliment principal ingéré par le veau de lait au cours de la production. Celui-ci fournit de l'énergie, des protéines (aCelui-cides aminés), des vitamines et des minéraux aux veaux. Dans le lactoremplaceur, les principales sources de protéines sont la poudre de lait écrémé et la poudre de lactosérum (Mornet et Espinasse, 1977). La poudre de lait écrémé est constituée
6
principalement de caséine, tandis que la poudre de lactosérum est constituée de lactoglobuline (45%), de lactalbumine (25%) et de globulines (11,5%) (Gautier et Labussière, 2011). Quand la protéine laitière est coûteuse sur le marché, elle peut être substituée par des protéines végétales comme le soya, la luzerne, la pomme de terre et la féverole (Mornet et Espinasse, 1977; Ruckebusch et al., 1991). Toutefois, la digestibilité des protéines laitières est meilleure que celle des protéines végétales quand le veau ne mange pas encore d’aliments solides en grande quantité, c’est-à-dire lorsqu’il est âgé de moins de quatre mois (Mornet et Espinasse, 1977; Ruckebusch et al., 1991). C’est qu’à cet âge, le veau digère 97% des protéines laitières. La digestibilité des protéines végétales augmente quand le veau a un mois. Toutefois, elle ne devient pas aussi efficace que celle des protéines du lait (Mornet et Espinasse, 1977; Ruckebusch et al., 1991). Quelques procédés sont mis en place pour améliorer la digestibilité des protéines végétales comme le chauffage et l’élimination partielle ou totale des glucides complexes qui sont connus pour favoriser le développement de la fermentation microbienne dans la partie distale de l’intestin (Mornet et Espinasse, 1977; Ruckebusch et al., 1991).
En ce qui concerne la matière grasse, un bon lactoremplaceur doit avoir une teneur en acide gras à longues chaînes (C18) inférieure à 20% et une teneur en acide de gras polyinsaturés inférieure à 4%. De plus, le pourcentage en acide gras à chaîne courte et moyenne ne doit pas être plus élevé que quinze et trente respectivement (Mornet et Espinasse, 1977). Généralement, les acides gras contenus dans le lait sont essentiellement sous forme de triglycérides (98%) et sont constitués à 65 pour cent d'acides gras saturés (Gautier et Labussière, 2011). Ainsi, les matières grasses du lactoremplaceur peuvent être soit le suif, le saindoux ou les huiles végétales peu insaturées comme le coprah, le palmiste et la palme (Mornet et Espinasse, 1977; Gautier et Labussière, 2011). Le suif et le saindoux sont des matières grasses animales et sont principalement constitués d’acides gras saturés à longues chaînes (Gautier et Labussière, 2011). Le gras du lait est une source d'énergie pour le veau et un important complément à la protéine du lait (Owen, 1991).
Du côté des hydrates de carbone, le lactoremplaceur est principalement constitué de lactose. Celui-ci provient de la poudre de lait écrémé et de la poudre de lactosérum, mais il peut également provenir de sous-produits laitiers tels que le concentré protéique de lactosérum. Par contre, le veau est sensible au niveau du lactose dans le lait. Un niveau de lactose supérieur à 40 pour cent augmente le risque de diarrhée s'il est âgé de moins de trois semaines (Owen, 1991). Une consommation excessive de lactose peut également occasionner une hyperglycémie, une hyperinsulinémie et une résistance à l’insuline (Toullec et Lallès, 1995). Il est donc important de porter une attention à la quantité de lactose que le veau va ingérer (Toullec et Lallès, 1995). Une
7
autre source principale d’hydrates de carbone du lactoremplaceur est l'amidon. L'aliment d'allaitement, s'il est enrichi en amidon, permet d'obtenir des carcasses plus lourdes, sans excès de dépôts adipeux (Mornet et Espinasse, 1977). Par contre, l'amidon est un hydrate de carbone complexe et il est moins assimilable par le jeune ruminant en début de vie si on le compare au lactose (Owen, 1991; Toullec et Lallès, 1995). Le lactose se digère rapidement dans l’intestin grêle, contrairement à l’amidon, qui se digère peu (Toullec et Lallès, 1995). La digestion de l’amidon augmente avec l’âge du veau et le développement de son rumen.
Les minéraux et les vitamines peuvent être incorporés directement dans le lactoremplaceur (Owen, 1991). Les quantités de minéraux et vitamines nécessaires à une bonne croissance sont généralement bien connues. Le ratio calcium/phosphore doit se situer entre 1.2 et 1.4 (Diorio et al., 1999). Les quantités de vitamines A, D et E peuvent varier grandement d’un lactoremplaceur à l’autre. Par contre, il faut faire attention à ceux qui ont une quantité massive de vitamine A (>10 000 UI/jour), car l’excès de vitamine A n’est pas éliminé par le veau, mais entreposé dans le foie (Diorio et al., 1999). Ainsi, une quantité autour de 1 700 UI de vitamine A par jour est recommandée, car la vitamine A aide à l’absorption et la distribution de la vitamine E (Kertz et al., 2017). Pour la vitamine E, une quantité entre 125 et 250 UI par jour a un effet bénéfique sur le statut immunitaire du veau (Diorio et al., 1999; Kertz et al., 2017). Comme mentionné précédemment, le fer est ajouté dans le lactoremplaceur, pour le bon développement du veau et pour son bien-être (Gygax et al., 1993; Diorio et al 1999). Le fer dans le lactoremplaceur sera détaillé à la section 1.6.2.
1.1.2 Élevage du veau en Europe comparé au Canada
La définition d’un veau lourd est un animal de l'espèce bovine domestique dont le poids vif est inférieur ou égal à 220 kg et qui n'a pas ses dents d’adultes (Mornet et Espinasse, 1977). La viande de veau est évaluée sous la forme de carcasse bovine, dont le poids inférieur ou égale à 130 kg avec une couleur typique soit une couleur rosée (Mornet et Espinasse, 1977). Depuis la nouvelle législation européenne en 2008, la définition commerciale du veau a changé (FranceAgriMer, 2010). On ne se réfère plus au poids pour décider si c'est un gros bovin ou un veau, mais à son âge (FranceAgriMer, 2010). Ainsi, le terme «veau» est utilisé pour des carcasses issues d'animaux ayant un âge inférieur ou égal à huit mois (FranceAgriMer, 2010).
Depuis le 1er janvier 2007, en Europe, les veaux en parcs collectifs ou individuels doivent pouvoir exprimer leurs mouvements naturels comme s'étendre, se reposer, se relever et faire leur toilette sans difficulté, et ce, tout au long de leur croissance (Conseil Européen, 2009). De plus, à partir de huit semaines, les parcs individuels sont interdits, sauf si l'animal est malade. Cette mesure
8
est justifiée par la nature grégaire des bovins (EUL, 2009). Toutefois, même si avant huit semaines, les parcs individuels sont permis, ceux-ci doivent permettre aux animaux d'avoir un contact visuel et tactile avec les autres veaux environnants. Les veaux ne doivent pas être attachés sauf pendant la distribution du lait durant une période d'une heure (EUL, 2009). Les locaux, les parcs, les équipements et les ustensiles doivent être nettoyés et désinfectés. Le sol des parcs ne doit pas être glissant pour empêcher que les veaux se blessent. L'aire de couchage doit être confortable, propre et drainée. La litière de couchage est obligatoire avant l'âge de deux semaines (Conseil Européen, 2009). Au Canada, on retrouve le même règlement depuis décembre 2017 (NFAC, 2017)
En Europe, pour la santé des veaux, il est obligatoire qu'ils reçoivent du colostrum de bovin dès leur naissance, c’est-à-dire durant les six premières heures de vie (Conseil Européen, 2009). De plus, tout veau malade ou blessé doit être soigné sans délai (Conseil Européen, 2009). Au Canada, il est fortement recommandé de donner du colostrum au veau dès sa naissance, mais ce n’est pas une pratique obligatoire (NFAC, 2017). Pour l’instant, dans la section santé du troupeau, un registre de santé et de soins est important pour la prévention et la détection des maladies. Avant l’utilisation de médicaments, un plan de santé doit avoir été discuté avec un vétérinaire (NFAC, 2017).
Les veaux sont nourris deux fois par jour. Aussi, ils doivent avoir accès à la nourriture en même temps que les autres animaux du parc (Conseil Européen, 2009). Les aliments doivent contenir suffisamment de fer pour assurer un niveau adéquat d'hémoglobine dans le sang soit un minimum de 4.5 mmol/litre de sang. Une ration minimale journalière d'aliments fibreux pour chaque veau âgé de plus de deux semaines est également obligatoire (Conseil Européen, 2009). L'accès à de l'eau fraîche en tout temps est obligatoire (Conseil Européen, 2009). Au Canada, les mêmes règles sont appliquées. Cependant, il y a une petite différence pour l’hémoglobine minimale dans le sang recommandé. La concentration en hémoglobine dans le sang au sein d’un même groupe doit être maintenue ou corrigée à au moins 8.5 g/dl (5.3 mmol/l), sauf durant les quatre dernières semaines d’élevage pour la production de veaux de lait (NFAC, 2017). Par contre, le niveau en hémoglobine sanguin ne doit pas descendre sous le seuil de l’anémie pour ces quatre dernières semaines (NFAC, 2017). Ainsi, si le taux d’hémoglobine sanguin est inférieur à 7.2 g/dl (4.5 mmol/l), les animaux doivent recevoir une supplémentation en fer, une obligation durant tout l’élevage (NFAC, 2017).
9
Pour ce qui est des importations pour les veaux en Europe, le pays exportateur doit avoir un certificat attestant que les animaux ont bénéficié d'un traitement équivalent à celui accordé aux animaux d'origine européenne (Conseil Européen, 2009). Au Canada, les produits de viande importés doivent être conformes aux normes canadiennes (ACIA, 2017). Les aspects pris en considération sont : la protection du consommateur (santé publique) et la prévention de l’introduction d’une maladie épizootique grave (santé des animaux) (ACIA, 2017). Les normes d’importation de viande au Canada sont similaires aux normes européennes. Les animaux doivent également avoir un certificat attestant que les animaux ont été élevés selon les normes établies dans l’entente faite entre le pays importateur et le Canada (ACIA, 2017).
Les pays européens peuvent avoir des différences au niveau de l'appellation de la viande de veau, mais tous utilisent la classification européenne pour la viande (FranceAgriMer, 2010). Cette classification consiste à désigner la couleur de la viande avec des chiffres, la conformation avec des lettres et l'état d’engraissement avec des chiffres (FranceAgriMer, 2010).
Pour ce qui est de la couleur de la viande, celle-ci est répartie en quatre classes (FranceAgriMer, 2010) :
• 1- Blanc • 2- Rosé clair • 3- Rosé • 4- Rouge.
La Conformation de la carcasse de l'animal est divisée en cinq classes : E. U. R. O. P. Le «E» est utilisé pour désigner une carcasse au développement musculaire exceptionnel avec un profil convexe de l'animal (FranceAgriMer, 2010). Ainsi, le quartier arrière est très épais, et le dos est épais et très large jusqu'aux épaules. La lettre «U» désigne une carcasse avec un fort développement musculaire et un profil convexe (FranceAgriMer, 2010). La lettre «R» désigne une carcasse rectiligne avec un bon développement musculaire (FranceAgriMer, 2010). Ainsi, le dos est relativement épais, mais manque d'épaisseur aux épaules. La lettre «O», pour sa part, est utilisée pour les carcasses avec un développement musculaire moyen. Le profil de la carcasse est généralement rectiligne voir concave (FranceAgriMer, 2010). La dernière lettre, le «P», est affectée aux carcasses avec un développement musculaire réduit et un profil concave. Le dos et les épaules sont étroits et les os sont saillants (FranceAgriMer, 2010).
10
Ensuite, les cinq classes pour l'état d’engraissement sont (tiré de FranceAgriMer, 2010) : • 1- Maigre : Aucune trace de graisse
• 2- Peu couvert : Graisses de couverture insuffisantes
• 3- Couvert : Légère pellicule de gras régulièrement répartie sur l'ensemble de la carcasse • 4- Gras : Graisses de couverture légèrement excédentaires. Le gras recouvre toute la
surface
• 5- Très gras : Graisses de couverture nettement excédentaires.
La classification canadienne de la viande est celle de l’USDA (BCRC, 2018). Elle est divisée en cinq classes : Canada Prime, Canada classe A, classe B, classe D, classe E (BCRC, 2018). Les meilleures catégories de viande au Canada se situent dans les quatre catégories suivantes (Tiré de BCRC, 2018) :
1. Canada Prime : Persillage très abondant 2. Canada AAA : Persillage abondant 3. Canada AA : Persillage peu abondant 4. Canada A : Persillage rare.
La majorité de la viande (97.1%) se retrouve dans la classe A (BCRC, 2018). La classe B est destinée aux carcasses d’animaux jeunes qui ne répondent pas à plusieurs exigences de qualité de la classe A (BCRC,2018). La classe D est appliquée aux carcasses d’animaux matures, les carcasses de vaches notamment (BCRC, 2018). Cette classe est généralement utilisée pour le bœuf haché (BCRC, 2018). La classe E est réservée aux taureaux adultes ou jeunes présentant une masculinité prononcée (BCRC, 2018).
La couleur de la viande de veau est cotée selon quatre niveaux au Canada. Le niveau 1 correspond à une viande très pâle, le niveau 2 a une viande de couleur rosée, le niveau 3 correspond à une viande rouge tandis que la viande de niveau 4 montre une couleur rouge foncé (Bovin du Québec, 2002).
La couleur de la viande est l’un des plus importants facteurs dans le classement du veau. Le détaillant et le consommateur font une distinction entre la viande de bœuf et celle du veau par la couleur de la chair (Bovin du Québec, 2002). La couleur de la viande de veau est déterminée à l’aide d’un appareil mesurant la « réflectance » de la couleur, au niveau de la poitrine (Bovin du Québec, 2002). Dans le veau, on recherche une couleur de chair qui est pâle, soit de niveau 1 et 2 (Bovin du Québec, 2002).
11
1.2 Croissance du veau : Système digestif
Le complexe gastrique des polygastriques est divisé en quatre parties soient le rumen, le réticulum (réseau), l'omasum (feuillet) et l'abomasum (caillette). Le rumen, le réticulum et l'omasum représentent les pré-estomacs. L'abomasum, quant à lui, est l'équivalent de l'estomac simple des monogastriques (Craplet et al., 1973). Le rumen est un large sac musculaire qui s’étend du diaphragme jusqu’au pelvis. Il occupe entre 80 et 85% du volume gastrique chez le ruminant adulte (Sautet, 1995). Le développement du rumen sera expliqué dans la prochaine section 1.2.1. Le réticulum, aussi appelé le réseau, a une muqueuse sous forme d’alvéole (Sautet, 1995). Il sert de tamis en retenant les particules trop longues dans le rumen (Jarrige et al., 1995). Il représente seulement 5% du volume gastrique des ruminants (Sautet, 1995). L’œsophage s’étend jusqu’au réticulum qui est logé en avant du rumen (Sautet, 1995). L’omasum, aussi appelé feuillet, est adjacent à l’abomasum. Il occupe 5% du volume gastrique (Sautet, 1995). Finalement, l’abomasum, aussi appelé caillette, est la partie glandulaire du système digestif (Sautet, 1995). Il occupe entre 7 et 8% du volume gastrique (Sautet, 1995).
1.2.1 Développement du système digestif
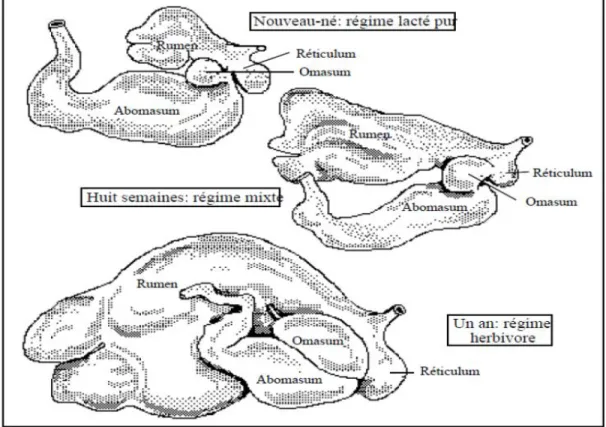
Lors de la naissance, le veau à l’état naturel a une alimentation exclusivement lactée. L'évolution de l'alimentation du veau est reliée avec le développement de la caillette (Mornet et Espinasse, 1977). Ainsi, à la naissance du veau, la caillette est dominante. Elle représente 50% de la capacité totale du complexe gastrique (Figure 1-1). Elle représente le double du réservoir rumino-réticulaire qui est pour sa part moins fonctionnel (Mornet et Espinasse, 1977; Ruckebusch et al., 1991). Ainsi, durant ses trois premières semaines de vie, le veau est considéré comme un monogastrique (Ruckebush et al., 1991). Quand le veau atteint l’âge de huit semaines, la caillette et le réservoir rumino-réticulaire sont de tailles similaires, car le complexe rumino-réticulum (réticulo-rumen) commence à se développer et devient, par le fait même, fonctionnel (Ruckebusch et al., 1991). Son régime alimentaire devient mixte, c’est-à-dire qu’il commence à ingérer des aliments solides en plus de son lait (Mornet et Espinasse, 1977). Ensuite, lorsqu’il est âgé de 12 semaines, le complexe rumino-réticulaire est rendu le double de la caillette et du feuillet réunis. Vers l’âge de quatre mois, la capacité du complexe rumino-réticulaire devient quatre fois plus grosses que la caillette et le feuillet (Mornet et Espinasse,1977). Finalement, c’est à l’âge d’un an que le veau est exclusivement herbivore (Ruckebusch et al., 1991).
12
Figure 1-1 : Développement du système digestif du jeune ruminant (Ruckebusch et al., 1991)
Les différentes parties de l'appareil digestif possèdent une croissance propre. La croissance du rumen est isométrique jusqu'à l'âge de quatre mois, c'est-à-dire qu'elle s'effectue au même rythme que la croissance de l'animal (Mornet et Espinasse, 1977). Par ailleurs, l'alimentation du veau influence le développement du rumen et de l'omasum. Si le jeune animal peut consommer du foin ou des concentrés à volonté, le rumen et l’omasum peuvent devenir deux fois plus développés que chez un veau restreint en concentré (Mornet et Espinasse, 1977).
1.2.2 La gouttière oesophagienne
La gouttière œsophagienne permet le passage direct des liquides du cardia jusqu'à l'orifice réticulo-omasal, le cardia étant la jonction oesophagienne (Mornet et Espinasse, 1977; Ruckebusch et al., 1991). Chez les jeunes ruminants, la gouttière œsophagienne se ferme suivant le réflexe du nourrisson.
Tout d'abord, les substances dégluties peuvent emprunter deux trajets différents. Elles peuvent tomber dans le complexe rumen-réseau ou se rendre directement dans la caillette grâce à la fermeture de la gouttière œsophagienne et la décontraction du feuillet (Craplet et al., 1973). La fermeture de la gouttière œsophagienne est un réflexe déterminé par l'ingestion du lait ou de l'eau
13
chez les jeunes ruminants (Craplet et al., 1973). Il s'atténue peu à peu avec le temps pour complètement disparaître chez le ruminant adulte.
Comme mentionné précédemment, chez le jeune animal, grâce au réflexe de fermeture de la gouttière œsophagienne, les aliments liquides et solides se rendent directement dans le feuillet et la caillette, le veau en début vie se comportant comme un monogastrique. L'ensemble rumen-réseau est détourné et l'activité microbienne est très réduite (Craplet et al., 1973). Il faut maintenir le réflexe de la gouttière œsophagienne pour l’élevage de veaux de lait, car une partie de la ration est constituée d’aliment liquide, c’est-à-dire de lactoremplaceurs ou d'aliments glucidiques qui doivent se rendre directement dans le feuillet et la caillette pour être digérée (Craplet et al., 1973). De plus, le complexe rumino-réticulaire se développe peu à ce moment, puisque seulement des liquides sont ingérés (Craplet et al., 1973).
Par ailleurs, ce sont les récepteurs dans la cavité buccale qui font fonctionner le réflexe de la fermeture de la gouttière œsophagienne (Ruckebusch et al., 1991). Quand les récepteurs pharyngaux ne sont pas correctement activés, le lait est transféré directement dans le complexe rumino-réticulum (Ruckebusch et al., 1991). Lorsque cet événement se produit, le veau est appelé ''buveur ruminal''. Pour éviter que le réflexe de la gouttière œsophagienne ne soit inactivé, il faut qu'il ait des changements dans la distribution du lait comme avoir recours aux tétines, respecter la température, les concentrations et les horaires de distribution du lait (Ruckebusch et al., 1991; De Passillé et Rushen, 2004). Par contre, selon Braun et Brammertz (2015), les conditions requises pour avoir le réflexe de la gouttière œsophagienne sont : un veau en santé et une odeur et un goût du lait normal, c’est-à-dire que le lait ne soit pas ranci. Les méthodes d’alimentation peuvent également affecter le réflexe discuté plus haut. En effet, Braun et Brammertz (2015) ont fait une étude sur l'effet de la température du lait, de la concentration du lactoremplaceur, du système d'alimentation et des grosseurs des tétines sur la fermeture de la gouttière œsophagienne. L'expérience de Braun et Brammerts (2015) a été faite sur six veaux Holstein. Pour toutes les concentrations en lactoremplaceurs (100, 125 et 150 g/l d’eau chaude à 41°C), le réflexe de la gouttière œsophagienne était activé et le lait n’a pas été observé dans le rumen (Braun et Brammertz, 2015). La température du lait autour de 20, 30 et 45°C a eu un effet positif sur le réflexe de la gouttière œsophagienne. Le réflexe étant actif, le lait ne va pas dans le réticulum (Braun et Brammertz, 2015).
14
Dans une autre étude, celle de Wise et al. (1984), une quantité de lait a été retrouvée dans le complexe rumino-réticulum pour les veaux qui buvaient leur lait avec une chaudière en immergeant leur museau dans le lait. Par contre, seulement quatre veaux étaient utilisés et seulement deux étaient de race Holstein, les deux autres étaient de race Guernseys, une race que l’on retrouve principalement en Angleterre et aux États-Unis (Wise et al., 1984). Pour la recherche de Braun et Brammertz (2015), impliquant des veaux alimentés avec une chaudière, seulement un veau a eu une faible quantité de lait dans son rumen. En conséquence, le réflexe de la gouttière n’a pas été activé lorsque le lait a été bu dans une chaudière avec un débit trop rapide, car le veau ne tétait pas (Wise et al., 1984; De Passillé et Rushen, 2004; Braun et Brammertz, 2015). Par contre, même si toutes les méthodes d'alimentation de cette étude ont démontré un bon fonctionnement du réflexe de la gouttière œsophagienne, il peut quand même y avoir un dysfonctionnement du réflexe si le veau boit de façon irrégulière son lait, c’est-à-dire avec un débit rapide et sans suce ou encore, s'il est malade (Braun et Brammertz, 2015).
Plusieurs études ont démontré que le réflexe de fermeture de la gouttière œsophagienne pouvait être activé par conditionnement (Orskov et al., 1970; Veissier et al., 2002; Braun et Brammertz, 2015). Ainsi, la vue du contenant avec du lait induirait le réflexe de la gouttière œsophagienne chez le mouton (Orskov et al., 1970; Braun et Brammertz, 2015). De plus, le bruit de la préparation du lait et l’odeur du lait sont des stimuli qui activent le réflexe de la gouttière œsophagienne, car le rythme cardiaque des veaux avant la tétée est supérieur à la normale (Figure 1-2) (Veissier et al., 2002; De Passillé et Rushen, 2004; Braun et Brammertz, 2015).
15
Figure 1-2 : Variations du rythme cardiaque chez les veaux nourris à la chaudière et nourris à la tétine autour du repas de lait le matin (Veissier et al., 2002)
Le réflexe de la gouttière œsophagienne peut être activé par un sel de sodium (Ruckebusch et al., 1991). Il peut également être inhibé par une injection d'atropine ou par l'application d'un anesthésique local dans la cavité buccale (Ruckebusch et al., 1991). Pour les jeunes ruminants, cette inhibition est utilisée quand on veut être certain que le médicament donné reste dans le complexe rumino-réticulaire (Ruckebusch et al., 1991).
1.2.3 Digestion du lait par le pré-ruminant
Le jeune veau est considéré comme un monogastrique pendant ses deux premières semaines de vie, la principale source d'alimentation étant le lait ou un lactoremplaceur qui est digéré dans la caillette (Roy, 1980). Puisque l'aliment principal de l'alimentation du jeune ruminant est liquide, l'ingestion de celui-ci se fait rapidement. Le réflexe de fermeture de la gouttière œsophagienne est stimulé et l’aliment liquide ingéré passe directement de l'œsophage à la caillette sans transiter dans le rumen (Guilhermet et al., 1975; Gautier et Labussière, 2011). De plus, le veau pré-ruminant a des glandes salivaires immatures, qui secrètent donc peu de salive (Guilloteau et al., 1995; Gautier et Labussière, 2011). Ces glandes salivaires sont dépourvues d'enzymes protéolytiques (Sissons, 1981; Gautier et Labussière, 2011) ce qui fait que la digestion des protéines commence seulement dans la caillette sous l'action des protéases gastriques (Guilloteau et al., 1995; Gautier et Labussière, 2011). La muqueuse glandulaire de la caillette sécrète trois endopeptidases : la chymosine, la
16
pepsine A et la pepsine B (Gautier et Labussière, 2011). Ces enzymes sont activées par l'hydrolyse sous l'action de l'acide chlorhydrique en milieu acide (Gautier et Labussière, 2011). La poudre de lait écrémé et le lait entier contiennent suffisamment de caséine pour permettre la coagulation dans la caillette. La coagulation résulte de l'édifice micellaire des caséines par l'acidification du milieu et l'hydrolyse de la caséine Kappa (Jensen et al., 2012). Cette hydrolyse aboutit à la formation de deux composés, la paracaséine-kappa et la caséinomacropeptide et provoque l'agrégation des micelles de caséines pour former le coagulum (Gauthier et Labussière, 2011; Jensen et al., 2012). Par contre, la formation du coagulum peut être altérée par un traitement thermique excessif du lait (Toullec et Lallès, 1995). Le traitement thermique change la composition du lait, donnant lieur à un pourcentage de protéine non-caséine plus faible et à une diminution du pourcentage de gras (Shillam et al., 1960). Par ailleurs, le chauffage du lait à 95°C pendant 45 secondes provoque un allongement des temps de coagulation et diminue l’arrivée intacte des caséines dans le duodénum (Toullec et Lallès, 1995). Ainsi, le lait chaud, c’est-à-dire ayant une température supérieure à 45°C, entraîne une accélération de l’évacuation gastrique des caséines et des lipides et une diminution de la protéolyse dans la caillette (Toullec et Lallès, 1995). Il est donc préférable de ne pas chauffer le lait excessivement, c’est-à-dire à une température supérieure à 45°C, pour une bonne digestion du lait dans la caillette et un coagulum de qualité (Toullec et Lallès, 1995).
1.3 Muscle
1.3.1. Myogenèse
La myogenèse est le processus de développement qui conduit à la formation du muscle squelettique (Cassar-Malek et al., 2010). Au début du développement, les cellules de l'un de trois feuillets primordiaux de l'embryon, le mésoderme, se différencient en cellules souches du muscle squelettique et ainsi, un muscle embryonnaire primordial se forme (Lawrie et Ledward, 2006; Cassar-Malek et al., 2010). La myogenèse procède en étapes successives qui impliquent différents types de cellules musculaires comme les myoblastes embryonnaires et fœtaux et les cellules satellites (Cassar-Malek et al., 2010). Les myoblastes, durant la myogenèse, prolifèrent et se retirent du cycle cellulaire et s'engagent dans une voie de différenciation pour former des myotubes (Sharma et al., 2001). Ainsi, pour former des myotubes multinucléés, les myoblastes fusionnent ensemble pour former une sangle musculaire et perdent, par la suite, leurs membranes cellulaires (Warriss, 2010). Autrement dit, les myotubes sont des fibres musculaires immatures et ils s’allongent pour devenir des fibres matures (Warriss, 2010). Seulement quelques cellules, appelées cellules satellites, ont la capacité de se répliquer après la naissance (Warriss, 2010). Ces cellules agissent comme une réserve pour le développement de nouveaux myoblastes (Warriss, 2010). Par
17
le fait même, les cellules satellites ne sont pas en phase terminale de différenciation. Elles démarrent leur phase de différenciation lors de blessures ou de dégénérations des fibres matures (Neuhaus et Braun, 2002). Le processus de différenciation de ces cellules est similaire à la myogenèse embryonnaire et il y a, finalement, une fusion ou un remplacement des myotubes blessés (Neuhaus et Braun, 2002). L’unité fonctionnelle pour la contraction dans les fibres se nomme sarcomères (Warriss, 2010).
De plus, la taille du muscle dépend en partie de l'étendue de la prolifération des myoblastes qui se produit avant la différenciation (Sharma et al., 2001). Les myoblastes croissent en longueur et vers la fin, les sarcomères s’ajoutent aux fibres (myofibrilles) (Warriss, 2010). Les fibres contractiles du muscle se nomment myofibrilles (Warriss, 2010). Le nombre de myofibrilles augmente par division longitudinale. Ainsi, les fibres musculaires acquèrent leurs propriétés contractiles et métaboliques (Cassar-Malek et al., 2010). Les propriétés contractiles des muscles bovins sont particulièrement avancées à la naissance (Cassar-Malek et al., 2010). Le nombre de fibres musculaires est déterminé génétiquement et il est fixé à la naissance également (Warriss, 2010). Seule sa taille augmente, le nombre reste le même tout au long de la croissance de l’animal (Warriss, 2010).
Pendant le développement embryonnaire, le processus de croissance musculaire est continu (Cassar-Malek et al., 2010). Au niveau cellulaire, l’accrétion du tissu musculaire peut être définie par l’augmentation du nombre (hyperplasie) et du diamètre et de la longueur (hypertrophie) des fibres (Cassar-Malek et al., 2010). Au cours du développement fœtal, la croissance musculaire se fait tout d’abord par hyperplasie (Cassar-Malek et al., 2010). Par la suite, l’hypertrophie des fibres se produit au cours de la période périnatale et postnatale (Cassar-Malek et al., 2010). Ceci implique également la fusion des cellules satellites avec les fibres existantes, contribuant à la croissance et à la régénération des fibres musculaires (Cassar-Malek et al., 2010). La formation de nouvelles fibres musculaires après le développement fœtal est vraiment limitée (Patel et al., 2002). Il est évident qu’il y a une augmentation remarquable de la taille des muscles à la naissance de l’animal et il y aura une augmentation continue pour des mois voire, même des années, tout dépendamment de la classe de l’animal (Patel et al., 2002).
18
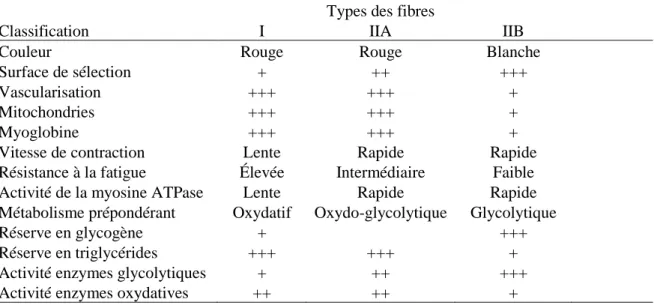
Au cours de la myogenèse, les fibres acquèrent leurs propriétés contractiles et métaboliques. Les fibres sont hétérogènes en raison de leurs différentes propriétés morphologiques et physiologiques (Jurie et Listrat, 2010). La classification des fibres musculaires repose sur un critère fonctionnel tel que la vitesse de contraction et sur un critère métabolique tel que le type de métabolisme énergétique (Jurie et Listrat, 2010). La plupart des classifications décrivent au moins trois types de fibres (Jurie et Listrat, 2010). Comme précédemment mentionnées, elles sont classées selon la vitesse de contraction, donc comme fibres lentes (type I) et fibres rapides (type IIA, IIX, IIB) (Jurie et Listrat, 2010). Les fibres de types I, à vitesse de contraction lente, possèdent une faible activité ATPasique et une activité glycolytique moins élevée que celle des fibres à contraction rapide (Jurie et Listrat, 2010). Les fibres I sont, également, plus riches en mitochondries et ont ainsi un métabolisme oxydatif (Jurie et Listrat, 2010) (Tableau 1-1). De plus, elles sont plus riches en lipides et ont un taux plus faible de glycogènes et de glucoses que les fibres rapides (Jurie et Listrat, 2010). Par ailleurs, les fibres de types I et IIA sont des fibres rouges, du fait qu’elles ont une vascularisation importante et une forte teneur en myoglobine (Jurie et Listrat, 2010) (Tableau 1-1). Ces deux types de fibres ont également une bonne résistance à la fatigue (Jurie et Listrat, 2010). Pour les fibres de types IIB, c’est l’opposé des fibres de types I et IIA, c’est-à-dire qu’elles ont une faible teneur en myoglobine, une faible vascularisation et une faible teneur en lipides (Jurie et Listrat, 2010) (Tableau 1-1). En conséquence, les fibres IIB ont une forte réserve de glycogène, donc elles ont un métabolisme énergétique glycolytique (Jurie et Listrat, 2010). Pour ce qui est du métabolisme énergétique des fibres IIA, c’est plutôt un intermédiaire qui se situe entre les fibres I et IIB, c’est-à-dire qu’il est oxydo-glycolytique (Jurie et Listrat, 2010) (Tableau 1-1).
Tableau 1-1 : Caractéristiques des principaux types de fibres du muscle squelettique Adapté de Jurie et Listrat (2010)
Types des fibres
Classification I IIA IIB
Couleur Rouge Rouge Blanche
Surface de sélection + ++ +++
Vascularisation +++ +++ +
Mitochondries +++ +++ +
Myoglobine +++ +++ +
Vitesse de contraction Lente Rapide Rapide
Résistance à la fatigue Élevée Intermédiaire Faible
Activité de la myosine ATPase Lente Rapide Rapide
Métabolisme prépondérant Oxydatif Oxydo-glycolytique Glycolytique
Réserve en glycogène + +++
Réserve en triglycérides +++ +++ +
Activité enzymes glycolytiques + ++ +++
19
1.3.2 Structure générale du muscle
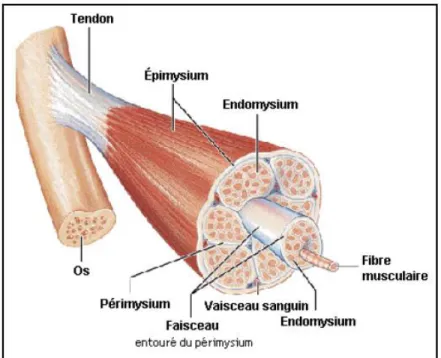
Le tissu musculaire représente 40 à 50% du poids vif des animaux domestiques (Jurie et Listrat, 2010). Les mammifères sont constitués de trois types de muscles : le strié squelettique, le lisse et le strié cardiaque (Cassens, 1987). Pour la production de viande, l'emphase est mise sur le muscle squelettique (Cassens, 1987 et Jurie et Listrat, 2010). La composition chimique de ce muscle est caractérisée par une forte teneur en eau (75%), en protéines (19%), dont 60% de protéines myofibrillaires et 10% de protéines du tissu conjonctif, et par une faible teneur en lipides (2.5%) (Jurie et Listrat, 2010). Un composé essentiel du muscle aidant à le maintenir soudé est la fibre (Lawrie et Ledward, 2006; Jurie et Listrat, 2010). Les fibres musculaires occupent 75 à 90% du volume musculaire (Jurie et Listrat, 2010). Elles sont des cellules avec plusieurs noyaux, c’est-à-dire plurinucléées, de 10 à 100 μm de diamètre. La longueur peut varier de plusieurs millimètres à plus de 30 cm (Lawrie et Ledward, 2006; Jurie et Listrat, 2010). Les fibres musculaires sont composées de faisceaux de myofibrilles (Cassens, 1987; Jurie et Listrat, 2010). Chaque myofibrille est constituée d'unités répétées appelées sarcomères (Jurie et Listrat, 2010). La structure ordonnée des sarcomères est basée sur l'alignement de filaments épais et fins (Jurie et Listrat, 2010). La myosine et l'actine sont les protéines majoritairement retrouvées dans les filaments (Jurie et Listrat, 2010). On retrouve également des mitochondries intermyofibrillaires et subsarcolemmales entre les myofibrilles et à la périphérie des cellules musculaires (Hocquette et al., 2000). Ces mitochondries sont responsables de la production d’une grande partie de l’énergie libre (ATP) fondamentale au muscle. L’ATP est produite à partir de substrats énergétiques apportés par la circulation sanguine, c’est-à-dire le glucose, le lactate, les acides gras, les triglycérides et les corps cétoniques ou par un milieu endogène, autrement dit le glycogène et les triglycérides (Hocquette et al., 2000). Les fibres musculaires sont aussi entourées d'un tissu de soutien que l'on nomme tissu conjonctif qui est l’endomysium (Jurie et Listrat, 2010). Une enveloppe plus épaisse de tissu conjonctif, aussi appelé périmysium, regroupe plusieurs fibres afin de former les faisceaux (Jurie et Listrat, 2010). Par ailleurs, c’est dans ce tissu que la majorité des vaisseaux sanguins irriguant le muscle est regroupée (Jurie et Listrat, 2010). Les différents vaisseaux sont finalement regroupés en muscle par une gaine encore plus épaisse que l’on appelle épimysium (Jurie et Listrat, 2010). C’est l’épimysium qui rattache le muscle à la structure osseuse et il est principalement composé de collagène (Jurie et Listrat, 2010). Le collagène est la protéine la plus commune dans la composition corporelle et de même, le composant majeur des tissus conjonctifs (Jurie et Listrat, 2010; Warriss, 2010) (Figure 1-3).
20
Figure 1-3 : Structure d’un muscle strié (Marieb et Laurendeau, 1993)
1.3.3. Transformation du muscle en viande
La transformation du muscle en viande repose sur des mécanismes biochimiques qui, après la mort des animaux, modifient la composition et la structure du muscle (Valin, 1984). Il y a plusieurs types de réactions qui sont mises en jeu dans le processus de transformation du muscle en viande (Valin, 1984). Des réactions hydrolytiques dissipent les réserves énergétiques du muscle soient l’ATP, la phosphocréatine et le glycogène et affectent l’état d’organisation des protéines, ainsi que sa structure (Valin, 1984). Le muscle reste biochimiquement actif, mais avec la diminution du fluide sanguin, le métabolisme passe d’aérobie (36 ATP formées par glucose métabolisé) à anaérobie [2 ATP de formées par glucose métabolisé] (Warriss, 2010). Le glycogène est la réserve de glucose pour l’énergie du muscle. La voie de transformation qui produit l’acide lactique par l’utilisation du glucose se nomme la glycolyse (Pearson, 1987; Warriss, 2010). Ainsi, le glycogène du muscle est changé en glucose, qui à son tour produit de l’acide lactique par la glycolyse. Cet acide est la cause de la diminution du pH dans le muscle (Pearson, 1987). Schématiquement, ceci est montré par la formule suivante (Pearson, 1987) :
21
Un facteur essentiel dans le processus de transformation du muscle en viande est la chute du pH occasionnée par l’accumulation d’acide lactique dans le muscle (Warriss, 2010). Le pH est un indicateur important pour le statut métabolique de la viande au stade «rigor mortis» (rigidité cadavérique) (Huff-lonergan, 2009). La production d’acide lactique et la valeur du pH ultime dépendent de la réserve en glycogène du muscle (Valin, 1984). Ainsi, une faible réserve en glycogène avant la mort de l’animal a pour effet de diminuer la production d’acide lactique, résultant à une viande pas assez acidifiée (Valin, 1984, Hocquette et al., 2000). Les réserves de glycogènes sont affectées par plusieurs facteurs externes tels que la manipulation des animaux, le transport, le stress ou l’activité physique avant l’abattage (Valin, 1984; Hocquette et al., 2000; Lawrie et Ledward, 2006).
Lors d’un stress causé par le transport ou une mauvaise manipulation des animaux, le taux de glycogène dans le muscle diminue de façon considérable pour se retrouver sous forme de glucose sanguin (Tableau 1-2). Par conséquent, le glucose sanguin est plus élevé lors que l’animal subit un stress, car cela permet au corps de réagir rapidement au danger (Tableau 1-2). Durant le transfert des animaux aux enclos de l’abattoir, l’animal peut être exposé à différentes sources de stress comme le jeûne, l’activité physique forcée, la rupture de son groupe social et de son environnement familier et le chargement et déchargement à l’abattoir. Tous ces stress peuvent affecter négativement la qualité de la viande (Valin, 1984; Hocquette et al., 2000; Van de Water et al., 2003; Mounier et al., 2006). Ainsi, il est important pour les animaux d’avoir un repos de 24 heures dans leur enclos à l’abattoir pour la récupération physiologique (Van de Water et al., 2003). En d’autres termes, pour refaire ses réserves de glycogène après le transport (Van de Water et al., 2003). Par conséquent, les bovins soumis à un stress élevé, comme un mauvais traitement lors de leurs manipulations à l’abattoir, ou lors du transport, démontrent une mobilisation de leurs réserves énergétiques, ce qui peut se traduire par une déplétion suffisante du taux de glycogène qui entraîne un pH ultime plus élevé (Valin, 1984). De plus, la resynthèse du glycogène est un phénomène très lent chez les bovins (Valin, 1984). Il n’y a aucune solution rapide pour accélérer la synthèse du glycogène (Valin, 1984).
22
Tableau 1-2 : Valeur moyenne pour les concentrations plasmiques avant et après le transport Adaptée de Van de Water et al. (2003)
Avant transport Après transport P1
Paramètres (Moyenne ±écart type) (Moyenne ±écart type)
Glucose (mg/dl) 101 ± 27 93,2 ± 30,1 *
Cortisol (ng/ml) 6,83 ± 10,84 17,3 ± 15,6 ***
Lactate (mmol/l) 19,0 ± 20,2 52,4 ± 32,9 ***
1 *, P <0,05; ***, P <0,001
Le pH du muscle est autour de 7,0, donc il est presque neutre. Après l’abattage, le pH diminue normalement jusqu’à environ 5,5 (Figure 1-4) (Lawrie et Ledward, 2006; Warriss, 2010). L’acidification est un facteur important pour la conservation de la viande. Pour la viande bovine, le pH est habituellement stable après 48 heures (Lawrie et Ledward, 2006). Par contre, si le muscle a un faible niveau de glycogène, le pH diminue lentement et se stabilise au-dessus de 6. De ce fait, la viande devient foncée, ferme et sèche (DFD : Dark, Firm and Dry) (Lawrie et Ledward, 2006). Si la chute de pH se fait plus rapidement que la normale et que le pH devient inférieur à 5.5, la viande est pâle, molle et exsudative (PSE : Pale, Soft and Exudative) (Lawrie et Ledward, 2006). Par conséquent, la chute du pH a un effet sur la couleur, la capacité de rétention en eau, la jutosité et la tendreté de la viande (Figure 1-5) (Hocquette et al., 2000).
23
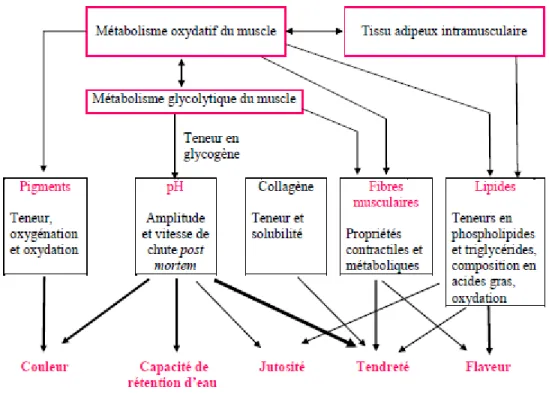
La couleur de la viande peut être influencée par le pH (Figure 1-5). La vitesse de chute du pH après l’abattage peut donner une viande DFD ou PSE (Hocquette et al., 2000). La couleur de la viande est également influencée par les pigments musculaires (Hocquette et al., 2000). La stabilité de la couleur dépend de l’état du pigment, de son oxygénation et de son oxydation (Hocquette et al., 2000). La jutosité et la flaveur de la viande sont influencées par la teneur en lipides du muscle, sa composition en acides gras et son état d’oxydation (Hocquette et al., 2000). La tendreté pour sa part varie selon la teneur et la solubilité du collagène et les propriétés contractiles et métaboliques des fibres musculaires (Hocquette et al., 2000). Par ailleurs, la tendreté est un peu influencée par la teneur en lipides du muscle (Hocquette et al., 2000). Par contre, le facteur qui a un effet sur la couleur de la viande, sur la capacité de rétention d’eau, sur la tendreté et sur la jutosité est l’évolution du pH post-mortem (Hocquette et al., 2000) (Figure 1-5).
Figure 1-5 : Relation entre le métabolisme énergétique musculaire et les caractéristiques musculaires déterminant la qualité de la viande (Hocquette et al. 2000)
24
1.4. Qualité de la viande
Pour le consommateur, la qualité de la viande est associée aux propriétés organoleptiques comme la couleur, la tendreté, la jutosité et la saveur. Plusieurs facteurs peuvent affecter la qualité de la viande comme le pH, le mode d’élevage de l’animal c’est-à-dire l’élevage en parc individuel ou collectif, l’abattage, l’âge des animaux, l’alimentation, le sexe de l’animal, etc.
1.4.1 Couleur
La couleur de la viande est le facteur le plus important pour les consommateurs, car elle affecte leurs décisions d’achat et leur acceptation (Muchenje et al. 2009). La couleur de la viande est constituée de trois indices soient le L*, le a* et le b*. Ces trois indices sont mesurés à l’aide d’un colorimètre. Le L* mesure la luminosité de la viande (100= blanc, 0 = noir). Ainsi, plus la valeur est élevée, plus la viande est claire (Mancini, 2009). L’indice a* mesure la rougeur de la viande. Il mesure le rouge positif et le vert négatif (Mancini, 2009). Ainsi, une augmentation de l’indice a* veut dire que la viande est plus rouge (Mancini, 2009). L’indice b* mesure, pour sa part, la couleur jaune (Mancini, 2009). Il mesure le jaune positif et le bleu négatif, donc plus la valeur de b* augmente, plus il y a de jaune dans la viande (Mancini, 2009).
La couleur de la viande peut être affectée par plusieurs facteurs, tels que les enzymes, l’alimentation, l’âge de l’animal et la race (Mancini, 2009). De plus, la pigmentation de la viande résulte de la présence dans la fibre musculaire d'une chromoprotéine renfermant le fer, la myoglobine (Charpentier et al., 1966; Labie et Eeckoutte, 1977). La molécule de myoglobine est constituée d'une simple chaîne de polypeptide de 153 acides aminés repliés autour d'un groupe prosthétique appelé l'hème (Bandman, 1987). L'extérieur de l'anneau est appelé 2-tétrapyrrole ou porphyrine. Le centre de l'anneau de l'hème contient un atome de fer où se lie l'oxygène (Bandman, 1987) (Figure 1-6).
25
Figure 1-6- Groupe hème de la myoglobine (Bandman, 1987)
Ainsi, plus la teneur dans la viande est élevée en myoglobine plus la viande est rouge. De plus, quand la myoglobine est sous la forme réduite, la viande est foncée (Labie et Eeckoutte, 1977). Le rôle de la myoglobine est de transférer à la chaîne respiratoire l'oxygène qui lui est apporté par le sang (Charpentier et al., 1966). En conséquence, la formation de cette chromoprotéine dépend de la disponibilité du fer dans le sang (Charpentier et al., 1966). Le fer peut provenir de l'apport alimentaire ou de la réserve accumulée dans le foie au cours de la gestation ou du fer libéré par la destruction des globules rouges (Charpentier et al., 1966).
Pour maintenir une viande de couleur pâle comme celle du veau de lait, il faut que le muscle, la viande, soit pauvre en myoglobine (Labie et Eeckoutte, 1977). Le lait des veaux est habituellement limité en fer, car l'addition de celui-ci dans la ration entraîne une augmentation de la synthèse de myoglobine et en conséquence, une couleur rouge de la viande (Labie et Eeckoutte, 1977). Pour avoir une viande de couleur pâle, il faut maintenir l’animal proche de l'anémie, c'est-à-dire avec un taux de fer dans le sang faible (7.2g/100ml) sans pour autant qu'il soit anémique (Labie et Eeckoutte, 1977; Enjalbert, 2009).