ETUDE EXPERIMENTALE DU TRANSFERT DE CHALEUR
A TRAVERS LES CORNES CHEZ LES BOVIDES:
DESCRIPTION ET IMPLICATIONS EVOLUTIVES
par
Karine Picard
memoire presente au Departement de biologie en vue de 1'obtention du grade de maitre es sciences (M. Sc.)
FACULTE DES SCIENCES
UNIVERSITE DE SHERBROOKE
Sherbrooke, Quebec, Canada, aout 1996
1*1
National Library of Canada Acquisitions and Bibliographic Services 395 Wellington Street Ottawa ON K1AON4 Canada Bibliotheque nationale du Canada Acquisitions et services bibliographiques 395, rue Wellington Ottawa ON K1AON4 CanadaYour file Votre reference
Our file Notre reference
The author has granted a
non-exclusive licence allowing the National Library of Canada to reproduce, loan, distribute or sell copies of this thesis in microform,
paper or electronic formats.
The author retains ownership of the copyright iti this thesis. Neither the thesis nor substantial extracts from it may be printed or otherwise
reproduced without the author's
permission.
L'auteur a accorde une licence non exclusive pennettant a la
Bibliotheque nationale du Canada de
reproduire, prefer, distribuer ou vendre des copies de cette these sous
la forme de microfiche/fihn, de
reproduction sur papier ou sur format electronique.
L'auteur conserve la propriete du droit d'auteur qui protege cette these.
Ni la these ni des extraits substantiels de celle-ci ne doivent etre unprimes
ou autrement reproduits sans son autonsation.
0-612-21816-3
ERRATA
1- A la page 46 (chapitre 2), V equation 2.7 aurait du se lire comme suit:
R -1 totale 1 hi-Aip+hi.Aie + 2-7i-L,-kk
lnf-^1
+• •Ie 'rrl-2e D. +1 h^-A^p+h^^eJ2- A la page 157 (annexe 6), 1'equation donnee au point b aurait du etre celle qui suit:
dT
;p.eau'meau^ _ ( -1-eau — -*-airll =— eau •p-(Aip + A^ ) (Teau ~~ T^ng, ou p=
dt
-T_ eau " airSOMMAIRE
Les comes des bovides sont constituees d'un comillon osseux fortement vascularise recouvert par un etui come de keratme n'offrant probablement
qu'une resistance limitee au transfert de chaleur. II est connu que les comes
jouent un role important dans les relations sociales et sexuelles, mais elles doivent aussi etre impliquees au niveau de la fhermoregulation.
La selection sexuelle semble orienter revolution de larges comes chez les bovides males, mais la perte de chaleur au niveau de ces larges comes doit imposer un cout energetique pour les animaux qui vivent dans des environnements froids. Afin de determiner si ces demiers ont developpe des adaptations pour limiter la perte de chaleur, la morphologie des comes des bovides des zones temperees a ete comparee avec celle des bovides des zones tropicales. Pour une longueur de come donnee, les bovides males des regions froides possedent de plus petits comillons osseux et des etuis comes plus epais par rapport a ceux retrouves chez les bovides des regions chaudes, restreignant probablement ainsi Ie flux de chaleur des comes vers Penvironnement. Chez les bovides males du genre Ovis, les especes et sous-especes des climats nordiques out des etuis comes plus epais que les sous-especes du desert. Le stress fhermique qui affecte la morphologie des comes pourrait etre une pression selective contrant la selection sexuelle chez les bovides males des zones
Dans Ie but de determiner si les caracteristiques morphologiques distinctes au niveau des comes des bovides des zones temperees versus celles des bovides des zones tropicales affectent difFeremment 1'echange de chaleur, la conductance thennique d'etuis comes a ete evaluee a Faide d?un modele theorique. La chaleur qui se dissipe du sang vers 1'environnement franchit trois resistances: entre Ie sang et les etuis comes, dans les etuis comes ainsi qu'entre les etuis comes et 1'air. Chacune de ces resistances a ete defmie par sa propre equation qui comporte les mesures des caracteristiques intrmseques (dimension, epaisseur et constitution) et extrmseques (epaisseur des couches limites et proprietes des fluides se trouvant a I'mterieur et autour des comes) des etuis comes qui sont impliquees dans Ie processus d'echange de chaleur. La conductance thennique au niveau des deux premieres resistances differe entre les bovides des zones temperees et ceux des zones tropicales. La perte de chaleur au niveau de la demiere resistance est correlee avec la longueur des comes mais ne differe pas
entre les climats. Les deux premieres resistances representent ensemble environ
50 % de la resistance totale, 1'autre moitie etant determinee par la resistance entre les etuis comes et 1'air. Chez les bovides des regions froides, la reduction dans la dimension des comillons osseux relativement a la dimension des etuis comes et Faugmentation dans Pepaisseur des etuis comes resulte en une restriction de la conductance thermique totale des etuis comes par rapport aux bovides des regions chaudes. La quantite de chaleur qu'un animal perd au niveau de ses comes n'est pas liee a sa capacite metabolique a soutenir un cout energetique donne. Enfin, 1'effet du vent sur la conductance thermique totale cTun etui come d'un mouflon d'Amerique (Ovis canadensis), une espece des
zones temperees, est moins important que sur celle d'un etii come d'oryx
algazelle (Oryx dammah\ une espece des zones tropicales.
La temperature a la surface des comes (Tsurf.comes) a ete mesuree sous des
temperatures ambiantes froides (Tambiante) chez un mouflon europeen (Ovis gmelinf) male originaire des regions temperees et chez deux aoudads (Ammotragus lervia), un male et une femelle, originaires des regions sub-tropicales. Des thermocouples ont ete places sur chacun des cotes interieur et
exterieur ainsi qu?a 1'amere d'une des comes des animaux. Sur chacun des
cotes, incluant 1'arriere des comes, un fhermocouple a ete place a 3 cm de la base et d'autres fhermocouples (quatre sur les cotes et cinq a 1'amere) etaient alignes a
distances egales jusqu'a 1'apex des comes. Pour les trois animaux et pour chacun des cotes, la Tsufcomes moyenne enregistree par les thermocouples diminue en allant vers Papex. Selon Ie profil de la Tsuif.comes etabli a 1'aide des donnees recueillies
simultanement par tous les fhermocouples, Ie mouflon europeen et les aoudads ne presenteraient pas Ie meme patron de vascularisation au niveau des comillons
osseux. L'examen des donnees de Tsurf.comes enregistrees par les fhennocouples places a la base sur la surface amere d'une des comes des animaux permet de
conclure que la Tambiante afifecte significativement Ie gradient fhemiique entre la
Tsuif.comes et la Tambiante (Tsuif.comes- Tgmbiante). Par COntre, IGS Correlations entre Ie (Tsurf.comes- Tambiante) et la Tambiante u'etaient pas toutes negatives telles qu'attendues. La Tguifcomes est periodique. La periodicite de la Tsurf.comes correspond a celle de la
temperature corporelle qui est reglee a environ 24 heures chez les mammtferes. La
determination de la Tguif.comes par les variations endogenes de la temperatiire
corporelle serait plus importante que celle qui serait due aux variations dans la
Les trois facettes de la recherche qui ont ete developpees dans ce memoire,
c'est-a-dire Fexamen de la morphologie des comes, des caracteristiques fhermiques des etuis comes et des temperatures de surface qui en resultent, pennettent de conclure que les comes des bovides joueraient un role au niveau de la fhermoregulation.
REMERCDEMENTS
Je tiens a remercier mes deux directeurs de recherche, Marco Festa-Bianchet et Donald W. Thomas, pour leur confiance, leur disponibilite et leur support tout au long de ce projet de maitrise. Je remercie aussi les deux autres membres de mon comite de conseillers. Hardy Granberg et William B. Shipley, pour leurs
encouragements ainsi que leurs expertises.
Je presente des remerciements au Musee canadien de la nature et au Royal Ontario Museum qui m'ont foumi Ie materiel necessaire aux experimentations. J'adresse un merci particulier a Andre Laneville et Fran9ois Belleville pour 1'aide qu'ils m'ont apportee dans Ie domaine de la thennodynamique et Ie temps qu'ils m'ont consacre. J'ai aussi grandement apprecie 1'aide de Patrick Cliche, un
ingenieur en electronique. Je suis tres reconnaissante envers les gens du Jardin
zoologique de Granby et les proprietaires de la Ferme d'elevage d'ongules de
Marbleton parce qu'ils m'ont donne acces a leurs animaux et m'ont ainsi permis
de realiser mon travail de terrain. Mille mercis a Stephane Demers pour F assistance qu'il m'a offerte a plusieurs niveaux dans ce travail. Enfin, j'aimerais remercier sincerement tous les etudiants gradues du laboratoire
d'ecologie (septembre 1993 a avril 1996) qui out participe de pres ou de loin,
d'une fa^on ou d'une autre, a 1'aboutissement de ce memoire.
Ce projet de recherche a ete subventionne par des fonds du Conseil de recherches en sciences naturelles et en genie ainsi que par des fonds pour la formation de chercheurs et d'aide a la recherche.
TABLE DES MATIERES
SOMMAIRE... i REMERCIEMENTS... v TABLE DESMATffiRES... vi LISTEDES TABLEAUX..,... x LISTEDES FIGURES... xm INTRODUCTION GENERALE... 1CHAPITRE 1 - PERTE DE CHALEUR VERSUS MORPHOLOGIE DES
CORNES... 61.1 Introduction... 6
1.2 Materieletmefhodes... 8
1.2.1 Mesures morphometriques et echantillonnage ... 8
1.2.2 Analyses statistiques... 12
1.3 Resultats... 12
1.4 Discussion... 17
CHAPITRE 2 - CONDUCTANCE THERMIQUE DES ETUIS CORNES 23
2.1 Introduction... 232.2 Materieletmethodes... 27
2.2.1 Technique utilisee et echantillonnage... 27
2.2.2 Modele theorique du transfert de chaleur a travers les comes... 30
2.2.2.1 Circuit fhermique equivalent... 30
2.2.2.2 Modelisation de cylindres a partir des mesures
prises sur les etuis comes... 32
2.2.2.3 Mesure de la capacite calorifique de la keratine
des etuis comes... 38
2.2.2.4 Mesure du coefficient de transfert de chaleur par
conductivite de la keratine des etuis comes... 38
2.2.2.5 Evaluation des relations temperature-temps du refroidissement a la temperature de la piece du systeme forme par les etuis comes chaufifes et 1'eau chaude
incorporee dans leur cavite... 42
2.2.2.6 Equations de la conductance themiique totale offerte par les etuis comes et du taux de transfert de
chaleur a travers les comes... 46
2.2.3 Analyses statistiques... 48
2.2.3.1 Morphologie des comes... 48
2.2.3.2 Resistance entre 1'eau et les etuis comes (Reau ou
m.p ^t -tveau.e^
2.2.3.3 Resistance dans les etiis comes (Rcome ou Rcome.p
ie.e>
2.2.3.4 Resistance entre les etuis comes et Fair (Rair ou
lir.p ®t Kair.e^
2.2.3.5 Resistance totale des etuis comes (Rtotaie)... 54 2.2.3.6 Contribution des trois resistances (Reau, Rcome et Rair) a la resistance totale des etuis comes (Rtotaie)... 55
2.2.3.7 EfFet du vent sur la conductance thermique totale
des etuis comes (Rtotaie )... 56
2.3 Resuhats... 57
2.3.1 Morphologie des comes... 57
2.3.2 Resistance entre Peau et les etuis comes (Reau ou Reau.p et
Reau.e)-• • ••••..•••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••..•••••••••••••• 65
2.3.3 Resistance dans les etuis comes (Rcome ou Rcome.p et
Rcome.e)... •...••... 68
2.3.4 Resistance entre les etuis comes et Fair (Rair ou Rair.p et
Rair.e)... 74
2.3.5 Resistance totale des etuis comes (Rtotaie)... 74
2.3.6 Contribution des trois resistances (Reau, Rcome et Rair) a la resistance totale des etuis comes (Rtotaie)... 82
2.3.7 Effet du vent sur la conductance thermique totale des etuis
comes (Rtotaie )... 82
2.4 Discussion... 84
2.4.1 Caracterisdques morphologiques des comes et leurs effets
sur Ie transfert de chaleur... 84
2.4.2 Resistance entre Peau et les etuis comes (Reau ou Reau.p et
Reau.e).••••••••...•••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••••• 87
2.4.3 Resistance dans les etuis comes (Rcome ou Rcome.p et
i.e^
2.4.4 Resistance entre les etuis comes et P air (Rair ou Rau-.p et
R^.e)... 94
2.4.5 Resistance totale des etuis comes (Rtotaie)... 96
2.4.6 Contribution des trois resistances (Reau, Rcome et Rair) a la
resistance totale des etiis comes (Rtotaie)... 99
2.4.7 Effet du vent sur la conductance thermique totale des etuis COmeS (Rtotale )... 101
2.4.8 Problemes potentiels dans la methodologie utilisee pour calculer la conductance thennique totale des etuis comes (Rtotale-1)... 103
CHAPITRE 3 - TEMPERATURE DES CORNES SOUS DES
TEMPERATURES AMBIANTES FROIDES... 1083.1 Introduction... 108 3.2 Materieletmethodes... 110 3.2.1 Mefhodes d'echantillonnage... 110 3.2.2 Analyses statistiques... 112 3.3 Resultats... 114 3.4 Discussion... 127 CONCLUSION... 134 ANNEXES... 137 BBLIOGRAPfflE... 165 IX
LISTE DES TABLEAUX
1.1 Liste des cranes de bovides males sur lesquels des mesures
morphometriques onteteprises... 11
1.2 Analyses de regression lineaire de 1'effet du climat, de la longueur de la come et de la masse corporelle sur la
morphologie de la come... 16
2.1 Liste des cranes de bovides pour lesquels la conductance
thermique d'un des etuis comes a ete calculee... 28
2.2 Liste des equations necessaires pour la modelisation des cylindres utilises pour calculer la conductance fhermique des etuis comes a partir du modele theorique du transfert de
chaleur a travers les comes... 35
2.3 Analyses de regression lineaire de Peffet du climat et de la
longueur de la come sur la morphologie de la come ... 60
2.4 Analyse de regression lineaire de 1'effet du climat et de la longueur de la come sur Ie ratio surface de la cavite / surface
ducylindre-modele... 63
2.5 Analyses de regression lineaire de PefFet du climat et de la
longueur de la come sur la conductance thermique entre 1'eau et P etui come (Reau ) par unite de sa surface exteme... 67
2.6 Analyses de regression lineaire de 1'effet du climat et de la longueur de la come sur la conductance thermique dans 1'etui
come (Rcome ) par unite de sa surface exteme... 70
2.7 Analyses de regression lineaire de FefFet du climat et de la longueur de la come sur Ie rapport de la conductance thermique dans la peripherie de I'etui come (Rcome.p ) et de la
conductance thermique dans 1'etui come (Rcome )... 72
2.8 Analyse de regression lineaire de FefFet du climat et de la longueur de la come sur Ie coef&cient de transfert de chaleur
entre Fetui come et 1'air (hi)... 77
2.9 Analyse de regression lineaire de Feffet du climat et de la longueur de la come sur la conductance fhermique totale de
I9 etui come (Rtotaie ) par unite de sa surface exteme... 79
3.1 Temperatures moyennes enregistrees par les quatorze fhermocouples places a diverses positions sur la surface d'une des comes (Tguif.comes) d'un aoudad (Ammotragus lervid)
femelle... 115
3.2 Temperatures moyennes enregistrees par les quatorze
fhennocouples places a diverses positions sur la surface d'une
des comes (Tsuif.comes) d'un aoudad (Ammotragus lervid) male.... 116
3.3 Temperatures moyennes enregistrees par les quatorze
fhennocouples places a diverges positions sur la surface d'une
des comes (Tsmf.comes) d'un mouflon europeen (Ovis gmelini)
male... 117
3.4 Comparaison du gradient fhermique entre les temperatures enregistrees par les fhermocouples places a 3 cm de la base sur
la surface arriere d'une des comes des trois animawc et les temperatures ambiantes (Tsuf.comes - Tambiante) avec la temperature
ambiante(Tambiante)... 118
LISTEDES FIGURES
1.1 Exemple d'une radiographie. II s'agit d'un etui come de
mouflon d'Amerique (Ovis canadensis)... 10
1.2 Residus de la regression entre la masse corporelle et Ie ratio
surface du comillon osseux / surface de 1'etui come en
foncdon de la longueur de la come ainsi que 1'efifet du climat
surles deux variables... 13
1.3 Relation entre 1'epaisseur de 1'etui come et la longueur de la
come ainsi que Peffet du climat sur celle-ci... 14
2.1 Representation schematique du circuit thermique propose
pour Ie transfert de chaleur a travers les comes... 31
2.2 Representation schematique des dimensions des cylindres-modeles utilises pour calculer la conductance fhermique des etuis comes a partir du modele theorique du transfert de
chaleur a traversles comes... 37
2.3 Representation schematique du montage utilise pour mesurer
Ie coefi&cient de transfert de chaleur par conductivite de la
keratine des etuis comes (kk)... 402.4 Equations de chacune des resistances du circuit thermique
propose pour Ie transfert de chaleur a travers les comes ... 47
2.5 Residus de la regression entre la masse corporelle et Ie ratio surface du comillon osseux / surface de 1'etui come en fonction de la longueur de la come ainsi que 1'efifet du climat
surles deux variables... 58
2.6 Relation entre Pepaisseur de 1'etui come et la longueur de la
come ainsi que FefFet du climat sur celle-ci... 59
2.7 Residus de la regression entre la masse corporelle et Ie ratio surface de la cavite / surface du cylindre-modele en fonction de la longueur de la come ainsi que FefFet du climat sur les
deux variables... 62
2.8 Relation entire Ie ratio surface de la cavite / surface du cylindre-modele et Ie ratio surface du comillon osseux /
surface de 1'etui come... 64
2.9 Relation entre la conductance thermique entre Peau et 1'etui come (Reau) par unite de sa surface exteme et la longueur de
la come ainsi que 1'effet du climat sur celle-ci... 66
2.10 Relation entre la conductance thermique dans 1'etui come (Rcome ) par unite de sa surface exteme et la longueur de la
come ainsi que 1'effet du climat sur celle-ci... 69
2.11 Relation entre Ie rapport de la conductance thennique dans la peripherie de Fetui come (Rcome.p ) sur la conductance thermique dans 1'etui come (Rcome ) et la longueur de la
come ainsi que Feffet du climat sur celle-ci... 73
2.12 Relation entre la conductance themiique entre 1'etui come et
Fair (Rair ) et la longueur de la come... 75
2.13 Relation entre Ie coefficient de transfert de chaleur par convection entre 1'etui come et P air (h^) et la longueur de la
come ainsi que Feffet du climat sur celle-ci... 76
2.14 Relation entre la conductance thermique totale de 1'etui come (Rtotaie ) par unite de sa surface exteme et la longueur de la
come ainsi que PefFet du climat sur celle-ci... 80
2.15 Relation entre la conductance thermique totale de Petui come (Rtotaie ) par unite de sa surface exteme et la masse metabolique pour les bovides des zones temperees et pour les
bovides des zones tropicales... 81
2.16 Proportion de la resistance: 1) entre 1'eau et 1'etui come (Reau), 2) dans 1'etui come (Rcome) ainsi que 3) entre I'etui come et P air (Rair) par rapport a la resistance totale de 1'etii
come (Rtotaie) pour les bovides des zones temperees et pour
les bovides des zones tropicales... 83
2.17 Relation entre la conductance themiique totale de Petui come (Rtotaie ) par unite de sa surface exteme et la vitesse du
vent... 85
2.18 Effet de Faugmentation de la valeur du coefficient de
transfert de chaleur par convection entre I'eau et 1'etui come
(hi) sur la conductance fhermique totale de 1'etui come
(Rtotaie'1) par unite de sa surface exteme... 106
3.1 Difference entre les temperatures enregistrees par un fhermocouple place a 3 cm de la base sur la surface amere d'une des comes d'un aoudad (Ammotragus lervia) femelle et
les temperatu-es ambiantes (Tgurf.comes - Tambiante) en fonction de la
temperature ambiante (Tambiante)... 119
3.2 Difference entre les temperatures enregistrees par un fhermocouple place a 3 cm de la base sur la surface arriere d'une des comes d'un aoudad (Ammotragus lervid) male et les
temperatures ambiantes (Tsurf.comes - Tambiante) en fonction de la
temperature ambiante (Tambiante)... 120
3.3 Difference entre les temperatures enregistrees par un
fhennocouple place a 3 cm de la base sur la surface arriere d'une des comes d'un mouflon europeen (Ovis gmelini) male et
les temperatures ambiantes (Tsurf.comes - Tambiante) en fonction de
la temperature ambiante(Tambiante)... 121
3.4. Temperatures ambiantes (Tambiante) et temperatures enregistrees
par un fhennocouple appose a 3 cm de la base sur la surface amere d'une come d'un aoudad (Ammotragus lervid) femelle
(Tsurf.comes) entre Ie 3 et Ie 10 mars 1994... 123
3.5. Temperatures ambiantes (Tambiante) et temperatires enregistrees
par un fhennocouple appose a 3 cm de la base sur la surface arriere d'une come d'un aoudad (Ammotragus lervid) male
(Tsurf.comes) entre Ie 21 et Ie 31 mars 1994... 124
3.6 Temperatures ambiantes (Tgmbiante) et temperatures enregistrees
par un fhennocouple appose a 3 cm de la base sur la surface arriere d'une come d'un mouflon europeen (Ovis gmelini) male
(Tsurf-.comes) CUti-e Ie 26 et Ie 30janvier 1995... 125
3.7 Temperatures enregistrees par deux fhennocouples apposes sur
la surface aniere d'une come d'un aoudad (Ammotragus ler^id)
male (Tsurf.comes) enti-e Ie 21 et Ie 31 mars 1994... 126
INTRODUCTION GENERALE
I/ introduction generale de ce memoire decrit brievement les fondements scientifiques qui out servi a 1'elaboration du projet de recherche. La logique des hypotheses testees et toutes les mformations necessaires a leur bonne comprehension se trouvent a 1'interieur des dififerents chapitres de ce travail.
Kitchener (1991) a decrit les comes des bovides comme etant composees d'un etui come, constitue de cellules d'alpha-keratine impermeables et plates, qui
recouvre une moelle osseuse spongieuse. La surface exteme du corps osseux et la surface du sinus a 1'interieur de ce corps osseux sont couverts d'un fin reseau
vasculaire (Taylor, 1963, 1966). La moelle osseuse, Ie sinus central et les tissus les entourant sont approvisionnes par 1'artere temporale superficielle aussi bien
que par les arteres comuales dorsale et ventrale du moins chez les chevres
domestiques Toggenberg (Capra hircus) (Taylor, 1963, 1966). On sait que,
chez les mouflons d'Amerique (Ovis canadensis), ces vaisseaux sanguins emergent a la surface du comillon osseux en d'iimombrables nids capillaires qui sont surtout concentres a 1'arriere du comillon dans ses parties mediale ainsi qu'apicale et comcident avec les portions les plus minces de Fetui come (Taylor, 1962). On peut presumer que ce patron de vascularisation soit plus ou moins celui retrouve pour la plupart des comes des bovides. Retenons surtout qu'un apport sanguin contmuel est manifestement necessaire dans un tissu vivant etLes comes joueraient un role proeminent dans les interactions sociales et sexuelles (Geist, 1966a, 1968). Or, on s'accorde pour dire qu'il existe, chez la majorite des especes de bovides, un dimorphisme sexuel prononce non seulement dans la grandeur et dans la forme des comes mais aussi dans leur utilisation
(Geist, 1966a; Eccles et Schakleton, 1986; Estes, 1991; Locati et Lovari, 1991).
L'elucidation des fonctions des comes a surtout ete orientee vers les males, leurs roles, chez ces demiers, etant plus evidents. Beaucoup de bovides malesdeveloppent de larges comes au stade adulte. C'est Ie cas, enti'e autres, des especes de la sous-famille des Caprinae (genres Ovjs, Capra, Ammotragus, etc.).
Par exemple, chez les mouflons d'Amerique, 1'impressionnant cercle que forment les comes des males represente environ 8 % du poids des animaux soit de 6 a 15 kg environ (Geist, 1966b). Generalement, il est suppose que les caracteres
sexuels extravagants, tels les comes, soient appams et maintenus par selection
sexuelle resultant de la competition intrasexuelle par les combats. Chez les bovides, quand les males se battent entre eux pour determiner 1'acces aux femelles en estrous, ce sont souvent ceux qui possedent les plus larges comes qui remportent les combats et qui ont Ie succes reproducteur Ie plus eleve (Geist, 1966a, 1966b). En consequence, la taille des comes doit etre sujette a une forte selection sexuelle (Geist, 1971; Hogg, 1988). Les comes constituent meme un des exemples les plus cites de selection sexuelle bien que leurs significations adaptatives aient ete largement discutees et disputees. A preuve, la litterature suggere quatre autres fonctions pouvant etre exercees par ces structures: elles seraient des indicateurs de rang de dominance, serviraient a la defense centre les predateurs, joueraient un role dans la thermoregulation et, finalement, seraient un indice de la qualite genetique des males sur lequel les femelles baseraient leur
choix de partenaire pour Faccouplement (Geist, 1966a, 1966b, 1968;
Stonehouse, 1968; Henshaw, 1969; Bubenik et Bubenik, 1990; Kitchener, 1991).
En contrepartie, Putilite des comes chez les femelles demeure pratiquementinexpliquee. C'est sans aucun doute la grande variete de formes de comes, leur
presence sporadique chez les femelles (un tiers des bovides femelles ne possedent pas de comes) et Ie manque de concordance des correlations entre leurs fonctions et Ie type d'habitat qui ont conduit differents auteurs a diverger
d'interpretations sur leurs significations (Kiltie, 1985; Estes, 1991; Locati et
Lovari, 1991).Comme il a ete mentionne ci-dessus, on a souvent attribue aux comes un role
dans la fhermoregulation comme stmctures de dissipation de chaleur lors de temperatures chaudes (Taylor, 1966; Stonehouse, 1968; Henshaw, 1969). Une etude realisee par Taylor (1963, 1966) sur Papport sanguin et Ie patron de vascularisation des comes de chevres domestiques Toggenberg suggere que ces demieres ont un role de thennoregulation parce qu'elles se vasodilatent en
reponse, entre autres, a la chaleur ainsi qu'a 1'exercice et se vasoconstrictent quand une chevre est au repos ou dans un environnement froid. Tout ceci n'est
pas etonnant car les extremites du corps sont d'une grande importance dans la regulation de la temperature corporelle meme chez les animaux dont Ie corps est bien isole (Scholander, 1954; Schmidt-Nielsen, 1990). Les comes joueraient done un role dans la thennoregulation parce que ce sont des extremites et non pas parce qu'elles auraient evolue specialement pour remplir cette fonction. En effet, ce serait simplement a cause du flot sanguin y circulant que ces extremites nues (la keratme n'ofifrant probablement qu'une resistance linutee au transfert de
chaleur) servent comme lieu de perte de chaleur quand la temperature ambiante
est basse ou quand la temperature du corps est trop elevee.
Plusieurs especes de bovides des regions nordiques sont pourvues de larges comes et Ie fait de posseder ces larges comes augmente 1'aire exposee au froid favorisant ainsi une plus grande perte de chaleur. Cette perte de chaleur accroitrait les couts de thermoregulation et la charge metabolique parce que, meme dans les froids extremes des hivers nordiques, les animaux doivent maiatenir Ie flux sanguin dans leurs comes afin de les proteger du gel (Schmidt-Nielsen, 1990). Les bovides males doivent done, en plus d'investir dans la
croissaace de leurs comes pour s'assurer d'un certain succes reproducteur,
probablement investir dans une plus grande production de chaleur quand ils sont soumis aux conditions hivemales.
L'inevitable flux de chaleur se dissipant dans Fenvironnement et la charge metabolique pouvant etre imposee sous des climats froids ont oriente les trois
facettes de la recherche presentee dans ce memoire, c'est-a-dire 1'examen de la
morphologie des comes, des caracteristiques thermiques des etuis comes et des temperatures de surface qui en resultent.
Dans Ie premier chapitre de ce travail, la morphologie des comes des bovides des regions froides est comparee a celle des bovides des regions chaudes. Les differences dans 1'epaisseur des etuis comes ainsi que dans la dimension des
presentee comme etant un facteur qui pourrait limiter la pression de selection sexuelle pour de larges comes chez les bovides males des regions temperees.
Au chapitre 2, un modele theorique du flux de chaleur a travers les comes a ete elabore afin d'estimer la conductance thennique d'etuis comes de diverses especes de bovides originaires de regions climatiques distinctes. Les valeurs trouvees pour les etuis de bovides des zones temperees ont ete comparees a celles trouvees pour les etuis de bovides des zones tropicales afin de determiner si les caracteristiques morphologiques distinctes affectent dififeremment Fechange de chaleur.
Si des caracteristiques morphologiques distinctes existent au niveau des comes des bovides des regions froides versus celles des bovides des regions chaudes et si, en plus, celles-ci affectent la dynamique de Fechange de chaleur, on s'attendrait a ce que la temperature a la surface des comes d'animaux originaires de ces deux regions soit differente sous les memes conditions de temperatures ambiantes basses. Les bovides des regions nordiques devraient presenter des comes plus froides et ainsi perdre moins de chaleur que les bovides adaptes aux regions chaudes sous une temperature ambiante froide donnee. Cette hypothese
fait Fobjet du chapitre 3.
CHAPITRE 1
PERTE DE CHALEUR VERSUS MORPHOLOGffi DES CORNES
1.1 IntroductionLes comes des bovides doivent etre impliquees dans la fhermoregulation parce qu'elles sont constituees d'un comillon osseux fortement vascularise recouvert par un etui come de keratine n'offrant probablement qu'une resistance linutee au
transfert de chaleur (Taylor, 1963, 1966; Bubenik et Bubenik, 1990). Taylor
(1963, 1966), Stonehouse (1968) et Henshaw (1969) ont suggere que les comes
presentaient une surface relativement grande par laquelle la chaleur pourrait etre dissipee quand I'animal fait face a un stress de temperature cause par un environnement chaud. La vascularisation et la conductance thermique qui pennettraient Pevacuation de chaleur sous des conditions climatiques chaudes devraient aussi imposer une charge metabolique aux animaux qui vivent sous des climats temperes plus froids. Meme si Taylor (1963, 1966) a demontre que les chevres domestiques Toggenberg sont capables d'abaisser la temperature de leurs comes pour ainsi reduire la chaleur perdue, Kitchener (1991) a note que certaines antilopes, gardees en captivite dans des jardins zoologiques dePhemisphere nord, ont souffert d'engelures aux comes. Un tissu vivant comme
Ie comillon osseux doit etre maintenu au-dessus du point de congelation afin d'eviter la necrose ou autres dommages importants. D en resulte que, sous des temperatures ambiantes basses, les comes doivent inevitablement etre une avenue pour la perte de chaleur. De fait, la perte de chaleur a ete evaluee comme
equivalant a 29% du taux de metabolisme basal pour un aoudad male (Ammotragus lervid) faisant face a des temperatures hivemales (Picard et al., 1994). Par contre, cette espece subtropicale n'a pas ete sujette a une selection
pour des traits pouvant reduire Ie flux de chaleur vers I'environnement.
Confrontes aux climats nordiques et, par consequent, a des pressions de selection differentes de celles rencontrees par les especes tropicales, la morphologie des comes des bovides des regions temperees aurait probablement evolue pour limiter Ie transfert de chaleur a ce niveau. Diminuer la perte de chaleur devrait etre particulierement important quand un animal est au repos et qu'il ne genere aucun exces de chaleur contrairement a ce qui se produit lors d'activites. Dans ces conditions. Ie transfert de chaleur par les comes devrait avoir a etre compense par thennogenese et representerait ainsi une charge metabolique additionnelle. Ce cout metabolique pourrait etre considerable notamment durant 1'hiver quand les animaux doivent survivre a la saison de vegetation dormante qui offre une ressource alimentaire de faible valeur nutritive (Albon et Langvatn, 1992). Les couts energetiques sont independants de la diete, mais 1'importance ou Pimpact sur Ie bilan energetique varie selon la qualite de la diete. En consequence, meme si les benefices apportes aux bovides males qui investissent dans la production de larges comes sont indeniables, les larges comes doivent aussi imposer un cout energetique appreciable.
Le transfert de chaleur par les comes serait affecte par trois principaux facteurs. Premierement, la temperature du sang qui arrive dans les comes detenninerait Ie gradient thermique Ie long duquel la chaleur est transferee dans Fair. La dimension des comillons osseux, quant a elle, determinerait la surface du reseau
capillaire duquel la chaleur est perdue. Pour finir, 1'epaisseur des etuis comes determinerait la resistance opposee au transfert de chaleur.
Si Ie fait de posseder de larges comes impose un cout energetique important aux bovides males vivant dans des climats froids, la selection natirelle agirait probablement sur les stmctures des comes qui afFectent la perte de chaleur. Or, il y aurait des differences au niveau de la morphologie des comes entre les especes des regions temperees et tropicales. Les deux hypotheses suivantes ont
alors ete testees:
1. Les bovides des zones temperees devraient avoir un plus petit ratio surface du comillon osseux / surface de 1'etui come que les bovides des zones
tropicales.
2. Les bovides des zones temperees devraient avoir un etii come plus epais que les bovides des zones tropicales.
1.2 Materiel et methodes
1.2.1 Mesures morphometriques et echantillonnage
Cinquante-quatre cranes de males de quinze especes de bovides out 6te utilises afin de mesurer la morphometrie d'une de leurs comes. Pour la plupart des specimens, 1'etui come de keratme a ete separe du comillon osseux et tous deux out ete recouverts d'une mince couche de peinture plastique. L'enduit de plastique a ensuite ete enleve et decoupe en petits morceaux plats qui out ete
places sur Fecran d'un analyseur d'images. L'analyseur d'images mesurait la
surface totale des morceaux et permettait ainsi de cormaitre la surface du comillon et de Fetui come. Quand il etait impossible de retirer 1'etui come du conullon osseux, la longueur et Ie diametore du comillon etaient mesures a partir de radiographies (figure 1.1). Par la suite, la surface du comillon etait estimee suivant la geometric d'un cone symetrique. Dans Ie present travail, les surfaces
sont presentees comme Ie ratio surface du comillon osseux / surface de 1'etui
come afin de tenir compte des differences dans la dimension des comillons osseux et des etuis comes. Des radiographies ont aussi ete utilisees pour mesurer 1'epaisseur de 19etui come a 1 cm d'intervalle Ie long du comillon
osseux. Pour chaque come, 1'epaisseur moyenne a ete etablie comme etant la moyenne de toutes ces mesures. La masse corporelle des individus a ete obtenue des registres des musees d'ou provenaient les cranes, des donnees personnelles
de Marco Festa-Bianchet sur les ongules sauvages (Festa-Bianchet et al., 1996) et aussi de deux ouvrages (Nowak et Paradiso, 1983; Berger et Cunningham,
1994). La longueur de la come consistait en sa mesure frontale de sa base
jusqu'a son apex. Les especes ont ete assignees a deux zones climatiques
(Tableau 1.1). Les especes des zones temperees habitent les regions ou Ie minimum aimuel est sous 0°C et les especes des zones tropicales vivent dans les regions ou Ie minimum annuel demeure au-dessus de 0°C (Anonyme, 1992).
Figure 1.1 Exemple d'une radiographie. II s'agit d'un etu come de mouflon d'Amerique (Ovis canadensis).
Tableau 1.1 Liste des cranes de bovides males sur lesquels des mesures morphometriques ont ete prises (N = 54).
Espece
Impala
Bubale
Aoudad
Antilope d'Amerique
Bison d'AmeriqueNilgai
Cephalophe
Antilope d'eau Chevre de montagne Boeufmusque Mouflon d'AmeriqueMouflon de Dall
Chamois
Saiga
Antilope hamachee
* Temoeree: Nom scientifique Aepyceros melampus Alcelaphus burselaphus Ammotragus lervia Antilocapra americana Bison bison Boselaphus tragocamelus Cephalophus sp. Kobus ellipsiprymnus Oreamnos americanus Ovibos moschatus Ovis canadensisOvis dalli
Rupicapra rupicapra Saiga tatarica Tragelaphus scriptus re mimmale amiuelle Zone climatique* Tropicale Tropicale Tropicale Temperee Temperee Tropicale Tropicale Tropicale Temperee Temperee Tropicale Temperee Temperee Temperee Temperee Tropicale est sous 0°C.N
2
1
1
1
5
1
1
4
1
1
6
17
9
1
1
2
Tropicale: region ou la temperature mmimale annuelle est au-dessus de 0°C.
1.2.2 Analyses statistiques
Une analyse de regression lineaire a ete realisee en utilisant Ie logarithme du ratio surface du comillon osseux / surface de Petui come comme variable dependante, Ie logarithme de la masse corporelle ainsi que la longueur de la come comme variables covariees et, finalement, la zone climatique comme facteur de categorisation. La masse corporelle et la longueur de la come affectent significativement Ie ratio surface du comillon osseux / surface de Petui come (log
masse corporelle: r = 0,48, d.d.l. = 52, P = 0,0002; longueur de la come: r = -0,45, d.d.l. = 52, P = 0,0007). La figure 1.2 presente les residus de la regression
entre la masse corporelle et Ie ratio surface du comillon osseux / surface de Petui come en fonction de la longueur de la come ainsi que 1'effet du climat sur les deux variables. Une deuxieme analyse de regression lineaire a permis de detenniner que la racine carree de 1'epaisseur de 1'etui come etait seulement affectee significativement par la longueur de la come (r = 0,76, d.d.l. = 52, P = 0,0001) et non pas par Ie logarithme de la masse corporelle (r = 0,16, d.d.l. = 52,
P= 0,24) (figure 1.3).
l.SResultats
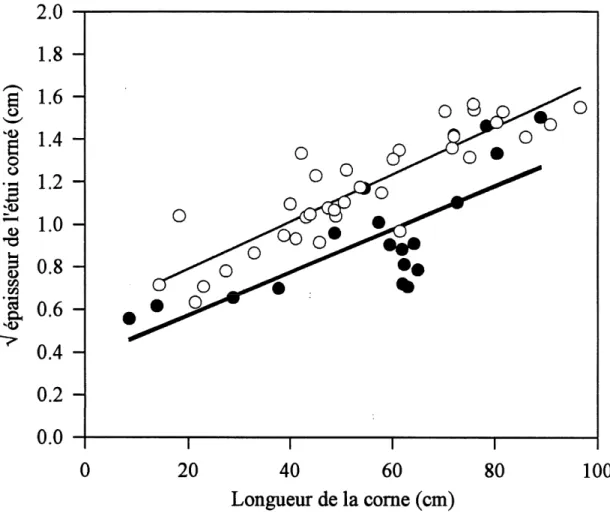
Les comes des bovides males des zones tropicales different morphologiquement par rapport a celles des bovides males des zones temperees. Chez les especes des regions chaudes telles Ammotragus lervta, Fantilope hamachee
(Tragelaphus scriptus) et Fantilope d'eau (Kobus ellipsiprymnus). Ie comillon
osseux vascularise a presque la meme surface que Fetui come. Au contraire,
-1.0
0
T40 60
Longueur de la come (cm) T80
100
Figure 1.2 Residus de la regression entre la masse corporelle et Ie ratio surface du comillon osseux / surface de 1'etui come en fonction de la longueur de la come ainsi que FefFet du climat sur les deux variables. Les points blancs et la droite de regression solide se rapportent aux bovides des regions temperees tandis que les points noirs et la droite de regression epaissie se rapportent aux bovides des regions tropicales.
0
I 1^- IT
20 40 60 80
Longueur de la come (cm)
100
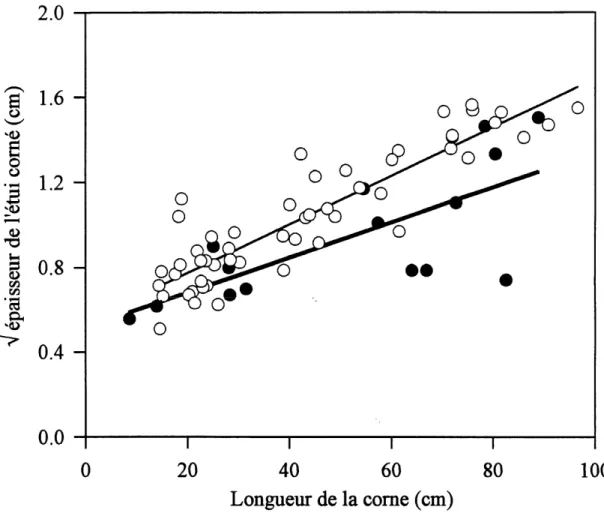
Figure 1.3 Relation entre 1'epaisseur de Fetui come et la longueur de la come ainsi que Feffet du climat sur celle-ci. Les pomts blancs et la droite de regression solide se rapportent aux bovides des regions temperees tandis que les points noirs et la droite de regression epaissie se rapportent aux bovides des regions tropicales.
chez les bovides des regions temperees comme Ie mouflon de Dall (Ovis dalli) et Ie mouflon d'Amerique, Ie comillon osseux ne represente en moyenne que 40,32 ± 13,40 % de la surface de 1'etui come. Les analyses de regression lineaire confirment que Ie climat affecte significativement Ie ratio surface du comillon osseux / surface de Petui come (Tableau 1.2). Pour une longueur de come donnee, les especes des zones temperees ont un plus petit ratio surface du comillon osseux / surface de 1'etui come et un etui come plus epais que les especes des zones tropicales (figures 1.2 et 1.3). Pour elimmer Ie probleme statistique potentiel relie au fait d'avoir regroupe un echantillonnage relativement grand d'Ovis (n = 32) avec seulement quelques donnees provenant d'autres bovides (n = 22), les memes analyses ont ete effectuees en incluant les donnees
d'un seul Ovis canadensis des zones temperees, d'un autre des zones tropicales
ainsi que d'un Ovis dalli, tous trois choisis aleatoirement panni 1'efifectif. Meme pour Pechantillonnage reduit. Ie climat affecte significativement Ie ratio surface
du comillon osseux / surface de 1'etui come (F = 7,87, d.d.l. = 1, P = 0,01) et 1'epaisseur de 1'etui come (F = 26,16, d.d.l. = 1, P = 0,001).
La difference observee dans Ie ratio surface du comillon osseux / surface de 1'etui come ainsi que dans 1'epaisseur de 1'etui come entre les bovides males des
regions temperees et ceux des regions ti-opicales pourrait avoir evoluee en
reponse aux proprietes mecaniques requises pour dijfferents styles de combats plutot qu'en reponse a la selection naturelle pour reduire les pertes de chaleur. La plupart des bovides des zones tropicales s'engagent dans des confrontations
consistant en une serie de poussees des adversaires en position tete-a-tete tandis que, de leur cote, beaucoup de bovides des zones temperees s'afifrontent en
Tableau 1.2 Analyses de regression lineaire de 1'effet du climat, de la longueur de la come et de la masse corporelle sur la morphologie de la
come.
A) Ratio surface du comillon osseux / surface de Fetui come* (N = 54)
Variable F P
Climat 14,18 0,0004
Masse corporelle* (kg) 32,23 0,0001
Longueur de la come (cm) 27,24 0,0001B) Epaisseur de 1'etui come* (N = 54)
Variable F P
Climat 37,38 0,0001
Longueur de la come (cm) 128,51 0,0001
* Les analyses ont ete realisees a 1'aide du logarithme du ratio surface du comillon osseux / surface de I'^tui come, du logarithme de la masse corporelle
et de la racine carree de 1'epaisseur de 1'etui come.
chargeant tete et comes premieres (Kitchener 1985, 1991). La charge resulte en
un choc extreme et pourrait demander une plus grande protection exteme sous la forme d'etuis comes plus epais, par exemple.
Afin de verifier I'implication evolutive possible du style de combat sur la morphologie des comes, les memes analyses que celles effectiees precedemment ont ete realisees sur trente-deux comes de bovides males du genre Ovis vivant en
Amerique du Nord. Ces animaux sont retrouves dans les deux zones climatiques mats n'utilisent qu'un seul style de combat, en 1'occurrence, la charge (Geist,
1971). Comme chez les bovides en general, la region climatique affecte
significativement Pepaisseur de 1'etui come (F = 4,82, d.d.l. = 1, P = 0,037).
Ainsi, les males du genre Ovis des regions temperees ont un etui come plus epais que ceux des regions chaudes. Tout autrement, aucun effet de la region climatique sur Ie ratio surface du comillon osseux / surface de I'etui come n'a ete
detecte chez les males du genre Ovis (F = 0,003, d.d.l. = 1, P = 0,99). Par
contre, Wehausen et Ramey (1993), avec un plus grand echantillonnage, out determine que les Ovis du desert ont un comillon osseux plus long que les Ovis
du nord pour un volume de come donne.
1.4 Discussion
Les analyses ont permis de demontrer que les bovides males habitant les regions froides ont au moins deux caracteristiques morphologiques pouvant probablement reduire la perte de chaleur a travers les comes. Les differences dans la morphologie des comes ne semblent pas s'expliquer par les contraintes
mecaniques pouvant etre imposees par dififerents styles de combats, mais semblent veritablement etre determinees par les contraintes thenniques. Le petit ratio surface du comillon osseux / surface de Petui come des bovides des zones temperees pourrait agir pour dimmuer la surface du reseau capillaire recouvrant Ie comillon osseux relativement a la dimension totale de la come. Une reduction dans la dimension du comillon osseux vascularise devrait dimmuer la surface par laquelle la chaleur est perdue dans 1'environnement. Le flux de chaleur ne serait done pas determine par la surface exteme de la come, mais aurait plutot lieu a
travers une plus petite surface. L'augmentation dans 1'epaisseur de 1'etui come
des especes des regions temperees comparee avec celle des especes tropicales pourrait contribuer a accroitre la resistance au flux de chaleur et devrait representer ainsi une intensification du facteur d'isolation. II est interessant de
noter qu'au niveau du genre (i.e. genre Ovis), Fepaisseur de I'etui come semble
plus soumise a la pression de selection qu'en est affecte Ie ratio des surfaces du comillon osseux et de I'etui come. On peut supposer qu'il existe une contrainte mecanique a ce niveau parce qu'une dimension minimale pourrait etre requise pour supporter 1'etui come de keratine relativement massive et les forces
absorbees lors des combats entre congeneres. Les analyses realisees sur les Ovis
avaient pour but d'examiner si les caracteristiques morphologiques etaient aussi
retrouvees au niveau du genre. Par centre, ces analyses n'ont pas pu etre repetees avec plus de taxa car il n5 a pas ete possible d'avoir acces a une variete
suf&sante de cranes de bovides. II serait evidemment interessant de determiner la robustesse des resultats obtenus en refaisant ces analyses pour d'autres groupes taxonomiques. Quelques taxa pourraient potentiellement etre candidats. On peut penser aux Rupicaprinae (Oreamnos, Rupicapra, Capricomis et
Nemorhaedus), aux bouquetins (Capra ibex) et aux chevres sauvages (Capra aegagrus). En efifet, ces deux groupes taxonomiques incluent des especes et sous-especes dont la distribution s'etend des regions chaudes et tropicales jusqu'aux regions froides subarctiques.
Dans son analyse, qui concemait la taille des comes en relation avec la
distribution geographique et climatique des bovides, Taylor (1966) n'a pas
trouve de correlation comme celle a laquelle on se serait attendu si les confraintes thermiques a£fectaient la moq)hologie des comes. De plus, il a estime la perte de chaleur a partir de 1'aire exteme de 1'etui come et if a pas tenu compte du fait que la surface exteme de la come et celle du comillon osseux a Finterieur pouvaient etre sujettes a differentes pressions de selection. Comme il est generalement accepte, les larges comes doivent etre Ie resultat de la selection sexuelle interagissant avec des facteurs environnementaux (Geist, 1971). Au contraire, la dimension du comillon doit etre sujette a dififerentes pressions selectives qui dependent du climat. De larges comillons osseux et de minces etuis comes devraient etre adaptes aux climats chauds des tropiques tandis que de petits comillons osseux et d'epais etuis comes devraient etre favorises sous des climats temperes plus froids.Les resultats de cette etude suggerent que la perte de chaleur au niveau des comes impose un cout sur la valeur selective des bovides males qui possedent de larges comes. Sans adaptations pour reduire la perte de chaleur, les males pourvus de larges comes se retrouveraient avec des couts de thermoregulation
eleves approchant ou meme excedant 29 % du taux de metabolisme basal comme
il a ete observe pour un aoudad garde en captivite dans un jardin zoologique quebecois (Picard et al., 1994). Quand les ressources alimentaires sont abondantes, les animaux devraient etre capables de soutenir cette charge
metabolique. Par contre, quand les animaux doivent survivre a la saison de
vegetation donnante limitee en qualite et en quantite de nourriture, la perte de
chaleur a travers les comes agirait comme une force contrant la selection sexuelle
pour de larges comes. De fait, la selection naturelle pourrait contribuer a limiter la perte de chaleur chez les bovides des zones temperees en diminuant la dimension du comillon osseux. Si un des roles du comillon osseux est de supporter Petui come relativement massif et de Ie proteger contore les chocs, il doit y avoir une limite mimmale pour la dimension du comillon osseux et, par
consequent, pour celle de Fetui come. C'est a ce moment que la selection pour
reduire la perte de chaleur devrait contrer la selection sexuelle pour une bonne
croissance de la come.
Ces suggestions, qui concement les contraintes thermiques influen^ant la dimension des comes ainsi que leur morphologie, sont supportees par Ie patron geographique dans la taille des comes des femelles de la famille des Caprinae chez lesquelles la selection sexuelle ne devrait probablement pas favoriser de
larges comes. Chez Ammotragus lervja, une espece adaptee au desert, les
femelles out des etuis comes d'approximativement 500 cm et des comillons osseux beaucoup plus volumineux que ceux que possedent les femelles du genre Ovis habitant les zones temperees et dont les etuis comes ne mesurent qu'environ 160 cm2. Meme parmi 1'espece Ovis canadensis, les femelles des especes du desert ont de plus longs comillons osseux que celles des
especes adaptees aux climats plus nordiques (Wehausen, 1989). Une difference dans 1'utilisation des comes pour la competition intraspecifique constituerait une alternative pouvant expliquer la difference dans la taille des comes chez les
femelles Ammotragus et Ovis. Par contre, il n'y a pas de donnees qui suggerent
que la competition pour Facces aux ressources alimentaires est plus grande pour les femelles Ammotragus que pour les femelles Ovis.
Parce qu'ils sont perdus a 1'automne, les bois des cervides ne representent pas un
site -d?echange de chaleur durant 1'hiver et n'imposent done pas de couts thermiques sous des conditions froides et difficiles comme c'est Ie cas pour les comes des bovides. Par centre, Ie velours et la vascularisation que les bois presentent durant la saison estivale les rendent propices aux echanges de chaleur. On pourrait alors croire qu'ils subissent une pression de selection naturelle pour maximiser leur surface de transfert de chaleur durant 1'ete quand les temperatures ambiantes sont elevees. Les bois des cenddes ne seraient done pas sujets a la meme pression de selection naturelle qui contrebalance la selection sexuelle pour de grandes comes chez les bovides. C'est pourquoi les selections naturelle et sexuelle auraient favorise des bois beaucoup plus imposants que les comes chez les cenddes du nord tels Ie cerf irlandais (Megaloceros giganteus) desomiais
eteint, Ie wapiti (Cervus elaphus), 1'orignal (Alces dices) et Ie caribou (Rangifer
rangrfer) (Geist, 1991). Evidemment, ces grands bois ne doivent pas aussi etreportes tout au long de 1'hiver comme c'est Ie cas pour les comes.
Note: Ce chapitre a ete accepte pour publication dans la revue Ecoscience sous Ie titre: « The cost ofhominess: heat loss may counter sexual selection for large
horns in temperate bovids ». Ainsi, dans les prochains chapitres, lorsque les resultats de cette partie du projet de maitrise seront cites, la reference de la publication sera donnee plutot que de se rapporter au chapitre 1 de ce memoire.
CHAPITRE2
CONDUCTANCE THERMIQUE DES ETUIS CORNES
2.1 IntroductionQuand la temperature ambiante est plus basse que la temperature du sang arteriel
emergeant a la surface des comillons osseux, la chaleur est necessairement
dissipee vers les etuis comes par convection forcee, a travers la keratine par conduction et, finalement, dans renvironnement par convection libre ou forcee selon qu'il y a absence ou presence de vent. Ici, il serait utile de definir les trois
modes de transfert de chaleur qui viennent d'etre mentionnes, c'est-a-dire la
conduction, la convection libre et la convection forcee. La conduction se fait entre deux corps physiques qui sont en contact et cela, peu importe leur etat (solide, liquide ou gazeux) (Schmidt-Nielsen, 1990). Plus exactement, elle consiste en un transfert direct de 1'energie cinetique d'un mouvement moleculaire et se produit toujours de la region la plus chaude vers celle qui est la plus froide (Schmidt-Nielsen, 1990). Le transfert de chaleur vers un fluide est presque toujours accelere par Ie processus de la convection qui refere au mouvement de masse du fluide. Prenons 1'exemple d'une surface chaude d'un solide qui est en contact avec un fluide froid. La chaleur est d'abord transferee du solide vers Ie fluide par conduction. Le fluide adjacent a la surface du solide devient alors plus chaud. Si Ie fluide est en mouvement. Ie fluide rechaufife adjacent a la surface du solide est alors remplace par du fluide froid. La chaleur perdue du solide vers Ie fluide est ainsi augmentee. fl y a convection libre lorsque Ie fluide rechaufife
adjacent a la surface du solide prend de Fexpansion et s'eleve pour etre remplace par du fluide plus froid tandis qu'il y a convection forcee lorsque Ie mouvement du fluide est cause par des forces extemes telles Ie vent et Ie courant (Schmidt-Nielsen, 1990).
Sous les climats tropicaux, les animaux utiliseraient la vasodilatation de leurs vaisseaux sanguins pour augmenter la quantite de sang qui circule au niveau des comillons osseux et pour permettre ainsi de dissiper 1'exces de chaleur genere par les activites metaboliques (Taylor, 1963, 1966). A partir de cette allegation, il pourrait etre suppose que la selection naturelle favoriserait 1'evolution de caracteres qui facilitent 1'echange de chaleur a travers les comes chez les
animaux des regions chaudes. L'evolution des caracteres pourrait notamment se
traduire par une augmentation dans la taille des comes et/ou dans la taille des
comillons osseux ainsi que par une reduction dans 1'epaisseur des etuis comes.
De telles adaptations morphologiques favoriseraient Ie transfert de chaleur et pennettraient ainsi aux bovides des regions chaudes de pouvoir evacuer la
chaleur en surplus a travers les comes. Par contre, sous des climats temperes, la
conservation de chaleur en hiver est probablement plus importante que la necessite de dissiper Ie surplus de chaleur accumule lors d'activites et/ou lors de la saison chaude. Sous ces conditions climatiques, les bovides doivent utiliser la vasoconstriction afin de restreindre Ie flux de sang et de chaleur dans les comes. Cependant, les animaux ne peuvent pas laisser geler les tissus vivaats des comes et cela impose une limite quant au degre de vasoconstriction ainsi qu'a la reduction de 1'echange de chaleur qui est possible. Les bovides dont les comes
presenteraient des caracteristiques morphologiques permettant une limitation du
transfert de chaleur pourraient beneficier d'une momdre charge metabolique lors de temperatures ambiantes froides. Ceci pourrait ^tre d'une tres grande importance quand 1'apport d'energie est reduit en saison de vegetation dormante
par la faible disponibilite d'une biomasse deja appauvrie en qualite (Albon et
Langvatn, 1992). Les comes des animaux habitant les climats temperes devraient done presenter des caracteristiques morphologiques offrant une certaine resistance au transfert de chaleur. Ces adaptations morphologiques pourraient prmcipalement se manifester par une reduction dans la taille des comes et/ou dans la taille des comillons osseux ainsi que par une augmentationdans 1'epaisseur des etuis comes.
U a ete demontre que les bovides males habitant les regions froides ont developpe au moins deux caracteristiques morphologiques pouvant fheoriquement reduire la perte de chaleur a travers les comes (Picard et al., sous presse). Pour une meme longueur de come, la reduction dans la dimension des comillons osseux relativement a la dimension des etuis comes retrouvee chez les especes des zones temperees versus celles des zones tropicales agirait pour diminuer la surface d'echange de chaleur en reduisant la surface du reseau capillaire approvisionnant les comes. De plus, les etuis comes des bovides des regions temperees qui sont plus epais que ceux des bovides des regions tropicales pour une meme longueur de come contribuerait a accroiti-e la resistance au flux de chaleur et representerait ainsi une intensification du facteur d'isolation. La selection naturelle favoriserait done des caracteristiques distinctes au niveau des comes selon que les especes viennent des zones tropicales ou temperees. Ces caracteristiques serviraient a faciliter ou a reduire
Ie flux de chaleur a travers les comes en fonction des conditions climatiques
auxquelles les animaux font face.
Plusieurs biologistes ont utilise la conductance thermique comme mesure specifique a un homeotherme permettant de comparer les adaptations thermobiologiques entre des especes vivant dans differentes regions climatiques (Scholander et al., 1950; Herreid et Kessel, 1967). La conductance themiique est la reciproque de 1'isolation ou de la resistance rencontree par Ie flux de chaleur qui se dissipe a travers un corps (Kreith, 1993). Dans Ie but de determiner si les caracteristiques morphologiques distinctes au niveau des comes des bovides des zones temperees versus celles des bovides des zones tropicales affectent differemment 1'echange de chaleur, la conductance thermique d'etuis comes a ete evaluee a 1'aide d'un modele fheorique du transfert de chaleur a travers les comes. Le modele fheorique combine les modes de transfert de chaleur par conduction et convection. Le transfert de chaleur par radiation pouvait dif&cilement etre reproduit vu la complexite et 1'mstabilite de ce phenomene en milieu naturel. Rappelons que Ie transfert de chaleur par radiation se produit en absence de contact direct entre deux objets, que tous les objets physiques dont la temperature se trouve au-dessus de 0°K emettent des radiations electromagnetiques (Pintensite et la longueur d'ondes des radiations dependent de la temperature a la surface des objets) et qu'ils en reyoivent des objets se trouvant dans leur voisinage (Schmidt-Nielsen, 1990). On comprend alors que les pertes de chaleur par radiation ont volontairement ete negligees dans cette etude a cause de la difficulte de les mesurer dans un contexte qui soit reel. Le modele theorique qui a ete utilise pour estimer la conductance thermique
de divers etuis comes tenait compte autant de leurs caracteristiques intrmseques que de leurs caracteristiques extrmseques. Les caracteristiques mtrinseques sont
celles liees, entre autres, a la forme, a la dimension, a Fepaisseur ainsi qu'a la
constitution des etuis comes tandis que les caracteristiques extrinseques se rapportent a 1'epaisseur des couches limites et aux proprietes des fluides se trouvant a 1'interieur et autour des etuis comes (Mitchell, 1976; Kreith, 1993).
2.2 Materiel et methodes
2.2.1 Technique utilisee et echantillonnage
La conductance thermique offerte en situation de convection libre par soixante-huit etuis comes de quinze especes de bovides a ete mesuree par la technique des courbes de refroidissement d'un systeme telle que decrite par Bakken (1976). Le systeme etudie etait forme des etuis comes ayant ete chauffes a 50°C et remplis d'eau chaude de 50°C qui se refroidissaient a la temperature de la piece (plus d'mformations sur la technique seront donnees dans la section 2.2.2.5). L'echantillonnage etait constitue de quarante etuis comes de male, vingt-quatre de femelle et quatre d'individu de sexe inconnu. Les especes ont ete assignees aux deux zones climatiques decrites dans Ie chapitre 1 (tableau 2.1).
L'evaluation de 1'effet de la convection forcee sur la conductance thermique des etuis comes a ete realisee en utilisant la meme technique que celle enoncee
ci-dessus mais a ete restreinte a seulement deux etuis comes. II s'agissait d'un etu
d'Ovis canadensis, une espece des regions temperees, et d'un etui d'oryx
Tableau 2.1 Liste des cranes de bovides pour lesquels la conductance thermique d'un des etuis comes a ete calculee(N=68). Espece Antilope sauteuse
Antilope d'Amerique
Bison d'AmeriqueNilgai
Cephalophe
Gazelle de Thompson Antilope rouanne Chevre de montagneNom scientifique
Antidorcas marsupialis Antilocapra americana Bison bison Boselaphus tragocamelus Cephalophus sp. Gazella thomsoni Hippotragus equinus Oreamnos americanus Zone climatique* Tropicale Temperee Temperee Tropicale Tropicale Tropicale Tropicale TempereeTaille de 1'
Femelles1
3
0
0
1
3
echantillon** Males1
5
1
1
0
1
Incomms1
0
0
0
0
0
1
0
* Voir page suivante.
bJ 00
Tableau 2.1 (suite). Espece Oryx algazelle Boeufmusque Mouflon d'Amerique
Mouflon de Dall
Chamois BongoNom scientifique
Oryx dammah Ovibos moschatus Ovis canadensisOvis dalli
Rupicapra rupicapra Tragelaphus euryceros Zone climatique* Tropicale Temperee Temperee Tropicale Temperee Temperee TropicaleTaille de V
Femelles0
11
2
3
0
echaatillon** Males1
16
6
7
1
Inconnus1
0
0
0
0
0
1
* Temperee: region ou la temperature minimale annuelle est sous 0°C.
Tropicale: region ou la temperature minimale annuelle est au-dessus de 0°C.
h<)
algazelle (Oryx dammah), une espece des regions tropicales. L'etui come d'Ovis canadensis a ete preleve sur Ie crane d'un male tandis que celui d'Ovjs
dammah provenait d'un specimen de sexe inconnu.
La surface du comillon osseux et la surface de 1'etui come de chaque specimen a 1'etude ont ete mesurees tel qu'explique au chapitre 1. La longueur des comes a aussi ete mesuree de la meme maniere que celle qui a ete decrite dans Ie chapitre precedent. La masse corporelle des individus a ete obtenue des registres des
musees d'ou provenaient les cranes, des donnees personnelles de Marco
Festa-Bianchet sur les ongules sauvages (Festa-Festa-Bianchet et al., 1996) et aussi de deux ouvrages (Nowak et Paradiso, 1983; Berger et Cmmmgham, 1994). La masse corporelle a ete elevee a la puissance 0,73 afin d'estimer la masse metabolique des animaux (Konoplev et al., 1978).
2.2.2 Modele theorique du transfert de chaleur a travers les comes
2.2.2.1 Circuit fhennique equivalent
Dans Ie but de decrire Ie plus explicitement possible la conductance thermique totale rencontree par Ie flux de chaleur se dissipant a ti'avers les comes (i.e. selon Ie systeme etudie: de 1'eau vers 1'environnement), un circuit fhennique a ete developpe. Un circuit thermique simple, comportant assez de details afin d'etre biologiquement raisonnable, a ete elabore a partir de cylindres-modeles pourvus de cavite plus ou moins profonde conformement aux dififerentes comes etudiees (figure 2.la). Une serie de trois resistances represente les votes importantes du
a)
sr^s
Eau -> Etui comeAir
sr~2
Eau Etii comePeripherie
ExtremiteAir
b)
iU )1meI-
Eau -Etui come-Air
-I
Figure 2.1 Representation schematique du circuit fhermique propose pour Ie transfert de chaleur a travers les comes, a) Systeme physique et son cylindre-modele. b) Prmcipales resistances renconfa-ees par Ie flux de chaleur: entre 1'eau et 1'etui come (Reau), dans 1'etui come (Rcome) ainsi qu'entre I'etui come et I'environnement (Rair).
flux de chaleur: entre 1'eau et les etuis comes (convection), dans les etuis comes (conduction) ainsi qu'entre les etuis comes et 1'environnement (convection) (figure 2.1b). Chaque resistance de la serie est constituee de deux resistances en parallele se rapportant aux deux regions stmcturales des cylmdres-modeles, la peripherie et Pextremite (figure 2.1c). La composition de chaque resistance (six au total) se decrit avec sa propre equation. Les dimensions des etuis et des cylindres-modeles (section 2.2.2.2), la capacite calorifique et Ie coefficient de transfert de chaleur par conductivite de la keratine des etuis (sections 2.2.2.3 et 2.2.2.4) ainsi que la relation temperature-temps du refroidissement a la temperature de la piece du systeme forme par les etuis chauffes et 1'eau chaude incorporee dans leur cavite (section 2.2.2.5) devaient tous etre connus avant de pouvoir evaluer la conductance fhermique totale des etuis.
2.2.2.2 Modelisation de cylindres a partir des mesures prises sur les etuis comes
Dans 1'objectif de defmir les dimensions des cylindres-modeles, certaines mesures out ete prises sur chacun des etuis comes pour lesquels la conductance
thermique a ete estunee.
Le volume total de chacun des etuis comes etudies (Vtotai (cm)) a ete calcule par la sommation du volume de chaque etui (Vcome (cm3)) et du volume de sa cavite
(Vcavit6 (cm3)). Pour mesurer Ie Vcome, les etuis ont ete tour-a-tour plonges en
entier dans un recipient gradue contenant de 1'eau. Le volume recherche correspondait au deplacement de la colomie d'eau (1 ml = 1 cm). Pour determiner Ie Vcavite, la cavite des etuis comes a ete remplie d'eau qui a ensuite
c)
Peripherie
lU.p me.p Rair.AT.?
I-
lEau
Etui comeAir
-I
m.u.e )]ime.e r.f
Extremite
Figure 2.1 (suite), c) Circuit thermique en serie combinant les principales
resistances rencontrees par Ie flux de chaleur se dissipant entre 1'eau et Petui come, par la peripherie (Reau.p) et par I'extremite (Reau.e), dans Petui come, par la peripherie (Rcome.p) et par 1'extremite(Rcome.e), ainsi qu'entre 1'etui come et 1'environnement , par la
peripherie (Rair.p) et par I'extremite (Rair.e).
ete videe et mesuree dans un cylindre gradue. Le volume de 1'extremite de chaque etui (Vextremite (ml)) a aussi ete evalue en utilisant la technique du deplacement de la colonne d'eau dans un recipient prevu a cet effet. Dans Ie present travail, Pextremite d'un etui est definie comme etant la partie s'etendant
du bout du comillon osseuxjusqu'a 1'apex de 1'etui lui-meme.
Le diametre moyen exteme de chacun des etuis comes (Dmoy (cm)) a ete obtenu en mesurant Ie diametre a tous les 5 cm Ie long de chaque etui a 1'aide d'un
mban a mesurer et en calculant ensuite la moyenne de ces donnees.
L'epaisseur moyenne de chacun des etuis comes (Emoy (cm)) a pu etre determinee grace a des radiographies des comes. Les images ont ete prises de fa^on a pouvoir observer Ie cote Ie plus epais des etuis (devant) ainsi que leur cote Ie plus mince (arriere). Des mesures ont ete prises directement sur les radiographies a tous les 1 cm autour du comillon osseux. Enfin, la moyenne de
ces mesures a ete calculee.
Les equations qui integrent les mesures prises sur les etiis comes et qui sont necessaires pour la modelisation des cylindres utilises pour calculer la conductance thermique des etuis comes a partir du modele theorique du transfert de chaleur a travers les comes sont definies dans Ie tableau 2.2. La representation schematique des dimensions des cylindres-modeles se trouve a la
figure 2.2.
Tableau 2.2 Liste des equations necessaires pour la modelisation des cylindres utilises pour calculer la conductance thennique des etuis comes a partir du modele theorique du transfert de chaleur a travers les
comes. Les equations integrent les mesures prises sur les etuis comes.
Variable Definition
Unite Equation
^D
ext Diametre exterieur cmD
moyD
int Diametre de la cavite cm D^oy - (2 • E^y'L. Longueur de la cavite cm cavite T extremite-.
..-moy '" -^'..-moy
Longueur de Fextremite cm 4.V.extremite
7T-Dmoy
Llp Aire peripherique inteme cm2 n • (D^y - 2 • E^ov) • L'moy/ -"c
A2p Aire peripherique exteme cm2 ;T-Dmoy-Lc
* Voir page suivante
Tableau 2.2 (suite).
Variable Definition
Unite Equation
li(cTT'(D^ -2-E^)2 Aie Aire inteme de 1'extremite cm2 " v—moy - —moy.
^2e Aire exteme de Pextremite cm2
7T-Dmoy ouDrmoy Vtotal cavlte Vextr6mite icy
diametre moyen des etuis comes (cm) volume total des etiis comes (cm3)
(Vcome (volume des etuis comes) + Vcavitd) volume de la cavite des etuis comes (cm ) volume de Fextremite des etuis comes (cm3) epaisseur moyenne des etiis comes (cm)
Dext Dint L, Ai, 4e
Peripherie
Extremite A2eFigure 2.2 Representation schematique des dimensions des cylindres-modeles utilises pour calculer la conductance thennique des etuis comes a partir du modele theorique du transfert de chaleur a travers les
comes (Dext= diametre exterieur, Dmt = diametre de la cavite, Lc = longueur de la cavite, Le = longueur de Pextremite, Aip = aire
peripherique inteme, Asp = aire peripherique exteme, Aie = aire inteme de Pextremite et Aie = aire exteme de Fextremite).
2.2.2.3 Mesure de la capacite calorifique de la keratine des etuis comes
Quelques morceaux de 1'etui come d'une chevre domestique Toggenberg (50 g) out ete chaufifes a 100°C et plonges dans Feau froide a 5°C. Aussitot, 1'energie
thermique des morceaux de I'etui etait transferee dans Feau. Parce que la
capacite calorifique de 1'eau etait connue (Cp.eau =4,18 J/g/°C), tout comme sa quantite (100 ml) ainsi que sa temperature finale lorsque Ie systeme etait a Pequilibre, la quantite de chaleur transferee des morceaux de 1'etui vers 1'eau froide a pu etre calculee a 1'aide d'une simple regle de trois. Ce calcul pennettait de connaitre la valeur de la capacite calorifique de la keratine des etuis comes (Cp.come (J/g/°C)) ou, par defimtion, la quantite de chaleur necessaire pour augmenter la temperature de 1 g de keratme par 1°C. Cette experience a ete
realisee dans un recipient de styro-mousse ferme. L'augmentation de la temperature de Peau a ete enregistree a I9 aide d'un thermocouple
(cuivre-constantan, 26 Ga) prealablement calibre et d'un logiciel d'acquisition des donnees (Datacan par Sable Systems). Cinq replicats de ces manipulations ont
ete faits et la valeur de la Cp.come retenue a ete etablie conune etant la moyeime de toutes les valeurs estimees.
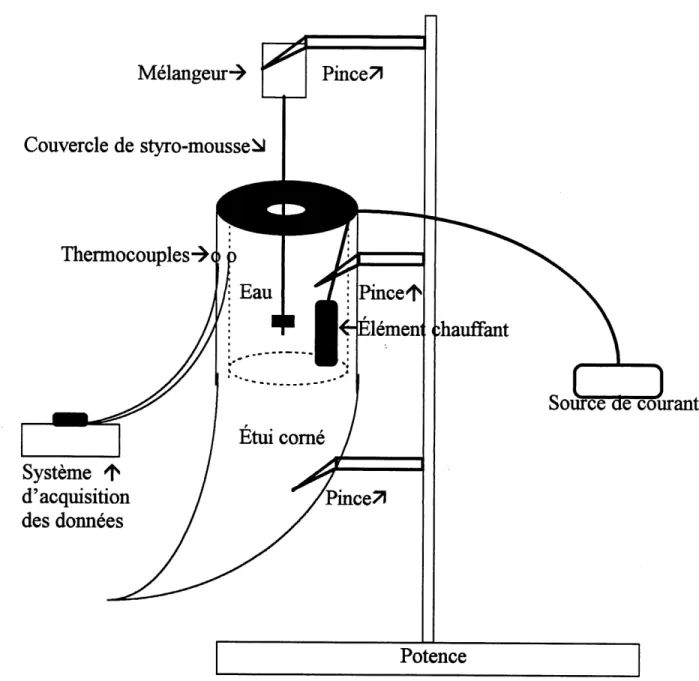
2.2.2.4 Mesure du coefficient de transfert de chaleur par conductivite de la keratine des etuis comes
Le coefficient de transfert de chaleur par conductivite de la keratine des etuis comes (kk (W/cm/°C)) a ete evalue a partir d'un seul etui d'Ovis canadensis male. Deux thennocouples prealablement calibres ont ete apposes sur la keratine