Les opioïdes : un verrou à la régénération chez le mammifère adulte ? Dialogue avec le système immunitaire et l'innervation sensorielle
323
0
0
Texte intégral
Figure

+7


Outline
Résumé scientifique
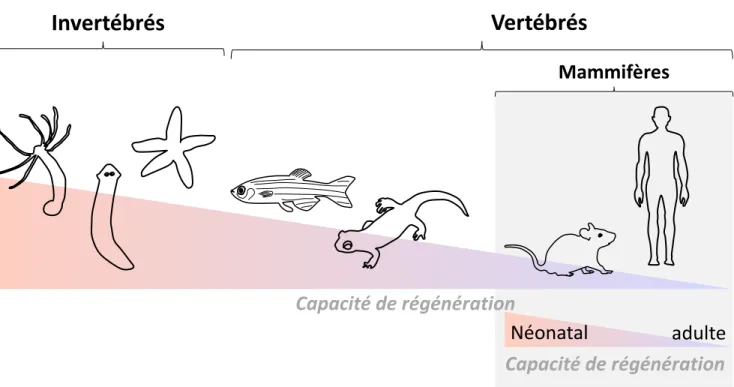
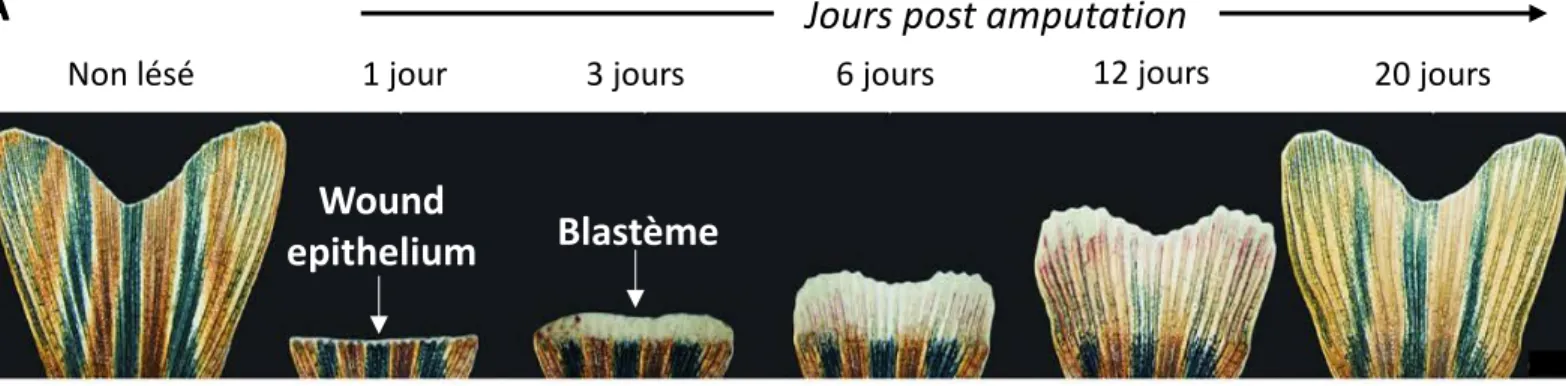
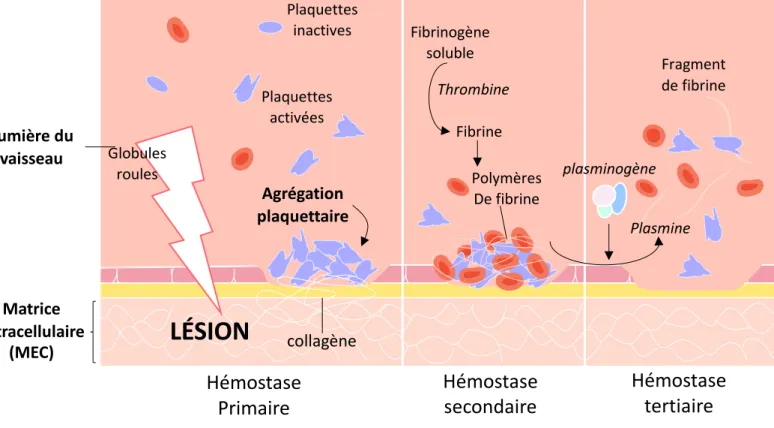
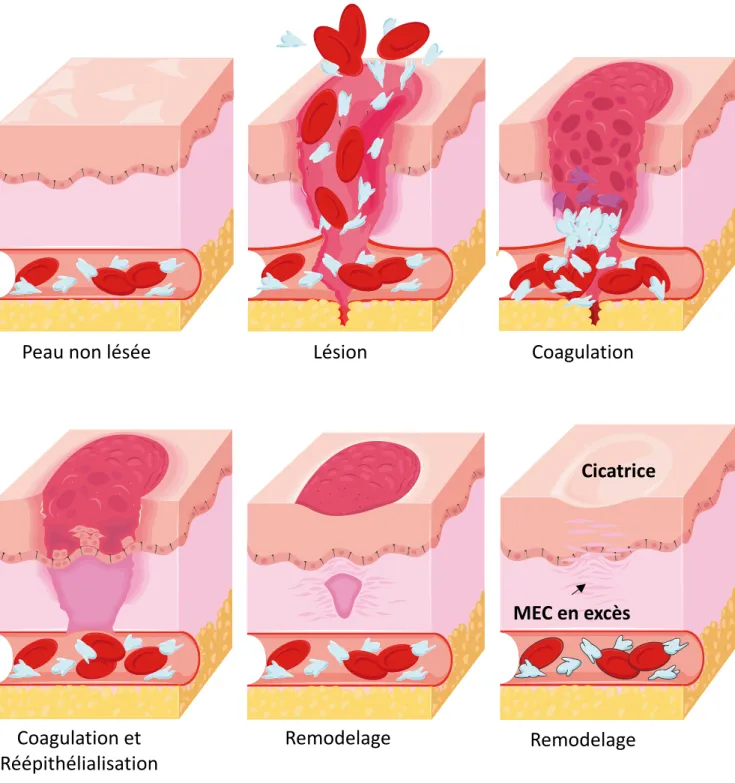
Figure 19 Système nerveux et régénération chez le mammifère adulte.
iii Les espèces actives de l’oxygène (EAOs)
II Les opioïdes
ii Conséquences cellulaires sur le nocicepteur
(1) Signature cellulaire et moléculaire des phases précoces de l’inflammation post-lésionnelle (2) Les neutrophiles produisent les EAOs nécessaires à la régénération (3) Les macrophages classiques dérivés de
Documents relatifs