Effet de la posture initiale du tronc sur le processus
d’initiation de la marche
Mémoire
Annie-Pier Fortin
Maîtrise en kinésiologie
Maître ès sciences (M.Sc.)
Québec, Canada
© Annie-Pier Fortin, 2015
iii
Résumé
L’initiation de la marche met en jeu des ajustements posturaux anticipatoires permettant l’exécution du premier pas. Ceux-ci sont caractérisés par un patron structuré de l’inhibition des soléaires suivis de l’activation des muscles tibiaux antérieurs. Le but de cette étude est d’analyser l’effet d’une inclinaison naturelle du tronc sur le patron moteur caractérisant l’initiation de la marche et de déterminer les conditions de posture initiale prédisant les deux composantes du patron.
Vingt-quatre jeunes adultes ont participé à cette étude. Deux groupes ont d’abord été créés à partir de l’angle du tronc et deux modèles de régression ont ensuite permis de prédire les deux composantes du patron, soit l’inhibition des soléaires et l’activation du TA.
Les inclinés arrière inhibaient les soléaires moins souvent que les inclinés avant et présentaient une activation initiale du soléaire plus petite et une co-activation initiale plus élevée. Les modèles de régression ont mis en évidence qu’une activation initiale du soléaire plus grande prédisait une inhibition.
v
Table des matières
Résumé ... iii
Table des matières ... v
Liste des tableaux ... vii
Liste des illustrations ... ix
Remerciements ... xi
Avant-Propos ... xiii
Introduction générale ... 1
Chapitre 1 : Recension des écrits ... 3
1 - Le contrôle de la locomotion... 3
1.1 - Le processus volontaire ... 3
1.2 - Le processus de régulation ... 4
1.3 - Le tronc cérébral ... 5
1.4 - Le processus automatique : La moelle épinière ... 7
2 - Les ajustements posturaux ... 8
3 - Initiation de la marche ... 9
3.1 - La phase d’anticipation ... 10
3.2 - La phase d’exécution ... 12
3.2 - Vitesse et direction ... 12
3.3 - Longueur et largeur du premier pas ... 14
3.4 - Jambe préférentielle versus jambe non-préférentielle ... 15
3.5 - Démarrage déclenché versus auto commandé ... 17
3.6 - Inclinaison du tronc ... 17
4 - Initiation de la marche et vieillissement ... 19
4.1 - La phase d’anticipation ... 21
4.2 - La phase d’exécution ... 23
5 - La variabilité ... 24
Résumé - Chapitre2 ... 27
Chapitre 2: Effect of natural trunk inclination on variability in soleus inhibition and tibialis anterior activation during gait initiation in young adults ... 29
Conclusion générale ... 49
vii
Liste des tableaux
Table 1: Initial condition at baseline and EMG pattern in backward leaner (BW) and forward leaner (FW) ... 36 Table 2: Soleus inhibition for backward leaner (BW) and forward leaner (FW) and for all subjects without groups division ... 37 Table 3: Prediction of soleus inhibition in swing and stance leg ... 40 Table 4: TA amplitude prediction in swing and stance leg ... 41
ix
Liste des illustrations
Chapitre 1
Figure 1: Boucles rétroactives des deux systèmes neurologiques impliqués dans la régulation du mouvement. ... 5 Figure 2: Les mouvements volontaires sont en premier lieu initiés dans le cortex prémoteur et le cortex moteur primaire qui sont régulés par les ganglions de la base et le cervelet. Le cortex moteur projette ensuite soit vers le tronc cérébral ou vers la moelle épinière. À partir de la moelle épinière, des motoneurones inférieurs (MN) activent les muscles qui permettent de produire le mouvement. . 7 Figure 3: Les ajustements posturaux anticipatoires (APA) débutent par l’inhibition des soléaires (SL) suivis de l’activation des tibiaux antérieurs (TA) et se terminent au décollement du talon (HO). Le centre de pression (CP) commence son déplacement de 100 à 200 ms après l’inhibition des SL. La phase d’exécution commence à HO et se termine par le contact du talon (HS). ... 10 Figure 4: Trajectoire du centre de pression (COP) et du centre de masse (COM) lors de l'initiation de la marche avec la jambe droite. Alors que le centre de pression se déplace en postérieur vers la jambe oscillante, le centre de masse amorce une chute vers l'avant et vers la jambe d'appui (5). ... 11 Figure 5: Déplacement du centre de pression (CP) selon la vitesse de marche lors d'un départ avec la jambe gauche (F=rapide, N=normal, S=lent). Le CP se déplace à de plus grandes amplitudes dans l'axe antéro-postérieur avec une augmentation de la vitesse (32). ... 14 Figure 6: Variations en médiolatéral de la trajectoire du centre de masse et de la largeur du premier pas lors d’un départ avec la jambe préférentielle et non-préférentielle (37). ... 16 Figure 7: Déplacement du centre de pression lors de l'initiation de la marche chez de jeunes adultes et des personnes âgées (40). ... 22
Chapitre 2
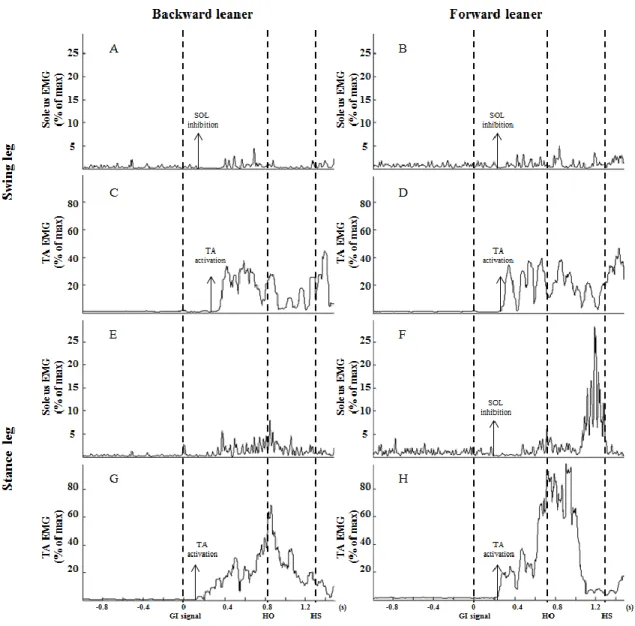
Figure 1: EMG pattern in a gait initiation trial of a representative BW and FW leaner in both legs. FW leaner shows a bilateral SOL inhibition while BW leaner shows a swing leg SOL unilateral inhibition. Panel A, C, E and G is EMG in swing leg soleus, swing leg TA, stance leg soleus and stance leg TA in a backward leaner. Panel B, D, F and H is EMG in swing leg soleus, swing leg TA, stance leg soleus and stance leg TA in a forward leaner. Soleus inhibition is present in panel A, B and F. The three vertical lines represent gait initiation signal (GI signal), off (HO) and heel-strike (HS). ... 38
xi
Remerciements
Tout d’abord, j’aimerais remercier mon directeur de recherche, Philippe Corbeil, de m’avoir supervisée tout au long de ma maîtrise. Merci de rendre les choses bien plus claires et évidentes et d’avoir pris le temps de discuter de questionnements à nature plus existentielle. Merci de m’avoir offert de bonnes opportunités.
Un merci spécial à Yoann Dessery de m’avoir tant aidée, et ce, à chaque étape de mon cheminement. Merci de ta grande disponibilité, de tes conseils et du temps que tu as pris à me former. Mes connaissances informatiques se portent sûrement mieux maintenant. Merci pour tout.
À Grant, merci de faire partie de ma vie, tout simplement.
Simon et Léandre. Merci d’avoir été mes amis, mais aussi de bons collègues autant lors des cours et des travaux que pour les soirées bien arrosées.
À Maxime et Isabelle, je suis heureuse d’avoir pu vous connaître. Ce fût un réel plaisir de pouvoir vous côtoyer au quotidien.
JP, Alexandra, Jérôme, Martin L, Olivier, Nico et Mathieu, chacun de vous m’a fait passer de bons moments. Merci.
À Normand Teasdale, Martin Simoneau et Marcel Kazsap, ce fût très agréable de vous rencontrer et d’entendre vos histoires du midi.
Merci à Sébastien Leteneur, Franck Barbier, Christophe Gillet et Émilie Simoneau de m’avoir accueillie dans votre laboratoire. Merci aux étudiants du LAMIH pour les nombreux fous rires.
Encore une fois, mes sincères remerciements à chacun d’entre vous. Le 2e sous-sol du
xiii
Avant-Propos
Effect of natural trunk inclination on variability in soleus inhibition and tibialis anterior activation during gait initiation in young adults
Annie-Pier Fortin, Yoann Dessery, Sébastien Leteneur, Franck Barbier, Philippe Corbeil
Cet article a été publié dans la revue Gait and Posture
Tous les auteurs ont donné leur approbation pour ce manuscrit. Yoann Dessery et Sébastien Leteneur ont préparé et participé à la collecte de données sous la supervision de Franck Barbier. L’article du chapitre 2 a été principalement rédigé par Annie-Pier Fortin en collaboration avec Yoann Dessery, sous la supervision de Philippe Corbeil.
1
Introduction générale
L’initiation de la marche se définit comme étant la période de transition entre la posture debout statique et le processus de marche stationnaire (1) qui s’explique par une combinaison d’activation musculaire et de forces gravitationnelles (2). Avant que le corps ne se place en état de locomotion, une phase initiale d’ajustements posturaux anticipatoires (APAs) permet de définir les futurs paramètres que l’on retrouve lors de la marche stationnaire (3,4).
Le phénomène dynamique précédant les premiers mouvements de la marche (décollement du talon), met en jeu des forces de réactions au sol (3–7). Les premiers évènements électromyographiques détectables lors de l’initiation de la marche sont l’inhibition des soléaires suivie par l’activation bilatérale des muscles tibiaux antérieurs (TA) se produisant en APAs avant l’exécution du premier pas (5,7–9). Une déficience dans la synergie de cette séquence musculaire pourrait amener des difficultés dans l’exécution du pas : une augmentation de la rigidité de l’articulation de la cheville, une instabilité posturale en phase de simple appui et une augmentation de la variabilité jumelée avec de petits pas de marche (10–12).
Des changements dans la fréquence du patron EMG sont observés chez des personnes âgées où les soléaires ne sont pas toujours inhibés avant l’activation du TA (12). De plus, l’activation du TA n’est pas toujours faite en anticipation chez cette population (12,13). Ce qui pourrait expliquer que la transition entre la posture debout et la marche chez les personnes âgées peut s’avérer difficile. Bien que l’on observe une diminution de la fréquence du patron moteur chez les personnes âgées, chez les jeunes adultes les soléaires ne sont pas toujours inhibés dans la plupart des essais (12). Certaines études antérieures chez de jeunes adultes sur l’initiation de la marche avaient inclus les essais dans leur analyse uniquement si l’activation initiale des soléaires était d’amplitude suffisante et si l’activation initiale des TA étaient inactive (9,12). Alors que ces essais ont été considérés comme une source d’erreur et retiré des analyses, ils pourraient faire parties de la variabilité normale du système et aiderait plutôt l’individus à s’adapter aux différentes contraintes de la tâche, de l’individus et de l’environnement (14). Il est nécessaire de comprendre cette variabilité pour avoir un regard complet du contrôle moteur lors de l’initiation de la
marche. Un premier enjeu consiste à identifier les facteurs pouvant expliquer en totalité ou en partie les variations du patron d’initiation de la marche. Cela s’avère fort important afin de mieux juger du caractère normal ou problématique de l’absence ou de la présence du patron d’activation musculaire lors des APAs de l’initiation de la marche.
Le premier chapitre de ce mémoire porte sur la recension des écrits. La première section de cette revue traite du contrôle de la locomotion alors que la deuxième et troisième section est axée sur l’initiation de la marche et sur l’effet du vieillissement sur le processus d’initiation de la marche, respectivement. Le deuxième chapitre du mémoire inclut un article à être soumis «Effect of natural trunk inclination on variability in soleus inhibition and tibialis anterior activation during gait initiation in young adults». Cet article traite de l’effet de la posture initiale du tronc sur la phase d’ajustements posturaux anticipatoires de l’initiation de la marche chez de jeunes adultes.
3
Chapitre 1 : Recension des écrits
1 - Le contrôle de la locomotion
La marche bipède est une tâche complexe nécessitant la coordination de plusieurs muscles qui permettent l’exécution de plusieurs pas consécutifs, amenant le pied à prendre un positionnement peu variable entre chaque pas tout en gardant le corps en équilibre. Cette tâche ne se simplifie pas seulement à marcher en ligne droite. Des changements de directions, d’environnements et de pentes sont plutôt des caractéristiques rencontrées dans cette activité de la vie quotidienne.
Le système nerveux central (SNC) joue un rôle important dans le contrôle de l’activité musculaire et dans la coordination des muscles qui assurent une marche répondant bien à l’environnement et amenant les modifications nécessaires au patron, si nécessaire (15). Le système nerveux central est divisé en trois niveaux hiérarchiques qui sont du plus bas au plus haut: la moelle épinière, le tronc cérébral et le cortex moteur. Chacun de ces niveaux permet la génération d’une réponse motrice selon les connections afférentes ou efférentes. Mis à part ces trois niveaux hiérarchiques, le système des ganglions de la base et le système cérébelleux situé dans le cerveau permettent l’initiation des mouvements et leur régulation par leur effet de modulation sur le cortex moteur (16). La moelle épinière possède aussi un réseau de connections impliqué dans le processus automatique de la marche (17).
1.1 - Le processus volontaire
L’aire motrice du cortex moteur est divisée en aire motrice primaire et en aire prémotrice qui permettent de contrôler le mouvement (16). Le cortex primaire moteur et l’aire motrice supplémentaire jouent un rôle différent dans la locomotion. Le cortex primaire moteur contribuerait aux mouvements des membres inférieurs alors que l’aire motrice supplémentaire, au contrôle postural lors de la marche. Cependant, le cortex prémoteur, qui inclut l’aire motrice supplémentaire, contribuerait aussi à la programmation et à la planification de la locomotion (18).
Les mouvements sont donc initiés au niveau du cortex moteur primaire et prémoteur. Le cortex moteur primaire reçoit de l’information du cortex somatosensoriel primaire et du cortex pariétal postérieur. Le cortex prémoteur et le cortex moteur primaire reçoivent aussi
de l’information des ganglions de la base et du cervelet via deux boucles rétroactives qui permettent de réguler les mouvements. Les aires prémotrices vont ensuite projeter vers le cortex moteur primaire et vers la moelle épinière. Cependant, davantage de projections vers la moelle épinière seront issues du cortex moteur primaire (16).
1.2 - Le processus de régulation
Alors que les mouvements sont initiés dans le cortex moteur primaire et prémoteur, les ganglions de la base et le cervelet forment deux boucles rétroactives et projettent vers le cortex moteur afin de réguler la locomotion (16).
C’est tout d’abord au niveau du système des ganglions de la base que se fait le contrôle des mouvements volontaires. Les ganglions de la base se regroupent en une unité fonctionnelle; le corps strié, qui est la combinaison de deux structures, soit le striatum et le pallidum. Ils sont le centre d’intégration du tonus musculaire, de la posture et des mouvements. Les ganglions de la base forment une boucle rétroactive avec le thalamus et le cortex cérébral moteur. Lors d’un mouvement volontaire comme l’initiation de la marche, le système des ganglions de la base est responsable de l’activation et de l’inhibition des motoneurones supérieurs. Il est aussi responsable de l’ajustement postural correspondant au même mouvement (19). Une lésion survenant aux ganglions de la base amène un syndrome d’akinésie qui consiste à une lenteur dans l’initiation des mouvements et à une réduction des activités motrices comme les mouvements volontaires (19). Chez les parkinsoniens, qui démontrent un trouble des ganglions de la base, la locomotion est caractérisée par des troubles lors de l’initiation de la marche, par des petits pas lors de la marche, par des instabilités lors de la propulsion et par des instabilités posturales (18).
Le cervelet entre aussi en connexion avec le cortex moteur par une boucle rétroactive. Le système cérébelleux permet plutôt la régulation des mouvements et leur coordination. Le vermis du cervelet est la partie impliquée dans les synergies musculaires lors de la marche et la posture debout. Il forme une boucle rétroactive le mettant en lien avec le cortex cérébral. La voie efférente du cervelet remonte jusqu’au thalamus puis jusqu’au cortex moteur. La boucle se continu dans la voie de retour jusqu’au cortex cérébelleux. L’un des syndromes de la marche associé à ce système résulte de l’atteinte du vermis qui amène une
5
ataxie à la marche s’apparentant à une démarche ébrieuse (19). La figure 1 illustre les deux boucles rétroactives du système d’initiation du mouvement (système des ganglions de la base) et du système de régulation (système cérébelleux). Ces deux systèmes reçoivent de l’information des différentes surfaces du cortex et projettent via le thalamus vers le cortex moteur.
Figure 1: Boucles rétroactives des deux systèmes neurologiques impliqués dans la
régulation du mouvement.
1.3 - Le tronc cérébral
L’aire motrice du cortex moteur influencera ensuite directement la moelle épinière par la voie corticospinale (ou voie pyramidale) ou descendra vers le tronc cérébral par la voie corticobulbaire ou vers la formation réticulée du tronc cérébral par la voie corticoréticulaire (Figure 2). Cette dernière serait fortement impliquée dans la préparation posturale précédant l’initiation de la marche (18).
Le cortex moteur peut influencer indirectement la moelle épinière via le tronc cérébral qui possède des voies descendantes vers celle-ci. Ces voies sont séparées en deux types : médiale et latérale (16). La voie médiale possède elle-même trois différentes voies;
vestibulospinale, réticulospinale et tectospinale tandis que la voie latérale contient la voie rubrospinale.
La grande partie des neurones provenant des centres supérieurs jouent un rôle d’excitation. Cependant, ils peuvent à la fois activer des interneurones excitateurs et des interneurones inhibiteurs. L’activation de ces deux types d’interneurones dépend du mouvement voulu et initié par les voies descendantes. La voie rubrospinale et corticospinale influenceraient les fléchisseurs et inhiberaient les extenseurs alors que la voie vestibulo-spinale faciliterait l’activation des interneurones qui contribuent à l’activation des extenseurs et l’inhibition des fléchisseurs (20,21). La moelle épinière contient des interneurones inhibiteurs Ia et les cellules de Renshaw localisés dans le Lamina VII. C’est la voie cortico-spinale qui active directement ces interneurones inhibiteurs Ia chez l’humain. Ils auraient pour rôle d’ajuster l’excitabilité des motoneurones inférieurs lors d’un mouvement et sont responsable de l’inhibition réciproque (21) qui amène une diminution simultanée de l’activité antagoniste lors de la contraction de l’agoniste (22).
7
Figure 2: Les mouvements volontaires sont en premier lieu initiés dans le cortex
prémoteur et le cortex moteur primaire qui sont régulés par les ganglions de la base et le cervelet. Le cortex moteur projette ensuite soit vers le tronc cérébral ou vers la moelle épinière. À partir de la moelle épinière, des motoneurones inférieurs (MN) activent les
muscles qui permettent de produire le mouvement.
1.4 - Le processus automatique : La moelle épinière
La moelle épinière possède aussi un réseau de connexion qui joue un rôle d’automatisme dans la locomotion. Des études chez les chats, où les racines dorsales étaient sectionnées, ne permettant alors pas de retour sensoriel vers les centres supérieurs, ont démontrés le rôle des générateurs de patrons spinaux de la marche (17). L’humain disposerait lui aussi de ce réseau complexe situé dans la moelle épinière, capable de produire un patron de marche rythmé sans intervention des centres supérieurs et de rétroactions des membres inférieurs. Une fois activé par les centres supérieurs, par la voie réticulospinale (18), un patron rythmé de flexion et d’extension est activé et celui-ci peut continuer par la suite sans nouvelles commandes des centres supérieurs (17,23). Les générateurs de patrons spinaux peuvent aussi être activés et modifiés via des afférences sensorielles des muscles des membres
inférieurs impliqués dans la flexion et extension (18,23). Ce patron de marche flexion et extension est donc utile lors de la marche en ligne droite ou encore sur une surface plane, où les centres supérieurs jouent un rôle minimal. Cependant, lors de modifications de l’environnement, où la perception de l’activité est nécessaire, les centres supérieurs sont indispensables (15).
2 - Les ajustements posturaux
Le centre de masse (CM) est l’emplacement où la masse d’un objet est également répartie. Chez l’humain, le CM est situé environ entre les crêtes iliaques lorsque le corps est placé en position anatomique. La position au sol de la projection verticale du CM varie en fonction de la position des segments corporels dans l’espace et la distribution relative des masses. Par exemple, une flexion de l’épaule à 90 ° fera déplacer le CM vers l’avant. Le centre de pression (CP) quant à lui, est le point d’application des forces de réactions au sol sous les pieds. Dans le cas où les deux pieds sont au sol, le CP se trouve à mi-chemin entre les deux chevilles, si le poids est également réparti entre les deux pieds.
L’exécution d’un mouvement volontaire, ne se résume pas seulement à l’activité musculaire et à la cinématique des segments impliqués dans le mouvement. L’intervention de d’autres segments entreront en jeu et permettront le maintien de l’équilibre. Ceux-ci sont des ajustements posturaux (24). Ces ajustements posturaux peuvent accompagner le mouvement avant que celui-ci ne se produise, pendant et après ce dernier (25). Les ajustements posturaux anticipatoires (APAs) sont des ajustements posturaux se produisant avant la perturbation de l’équilibre qui suivra nécessairement un mouvement du corps dans l’espace. Par exemple, lors d’un mouvement volontaire à partir de la position debout, comme la flexion du bras, l’inhibition ou l’activation des muscles impliqués dans le contrôle de la posture sont les premiers à être activés (26,27). Ce sont des ajustements posturaux anticipatoires qui permettent de maintenir le CM à l’intérieur de la base de support. Le contrôle postural est anticipé par rapport au contrôle du mouvement du bras, ce qui prévient la perturbation de l’équilibre associée à l’exécution du mouvement de bras (27). La génération des APAs est affectée par différents facteurs comme par l’amplitude et la direction du mouvement à effectuer. De plus, en raison de leur effet déstabilisateur, les APAs seraient éliminés dans une situation d’instabilité posturale (28). La posture initiale
9
les modifierait aussi, comme dans le cas d’un levé sur les orteils, où une inclinaison avant ou arrière du tronc modifie les APAs au niveau du temps et de la force des muscles en préparation (29). Les APAs sont aussi indispensables à la création du mouvement où celui-ci serait compromis s’ils étaient absents, d’amplitude insuffisante ou bien encore décalés dans le temps (29,30).
3 - Initiation de la marche
Lors du contrôle postural en position debout, le CP se déplace en antéropostérieur et en médiolatéral et est contrôlé respectivement par l’activation des muscles de la cheville et de la hanche afin de maintenir le CM à l’intérieur de la base de support. Lors de la marche, le contrôle de l’équilibre devient différent de celui en position debout. Tout d’abord, lors de l’initiation de la marche, une chute avant volontaire et contrôlée du CM devant la base de support permet d’accélérer le corps vers l’avant. Ensuite, une fois la marche entamée, le CM se déplace sur le bord médial de chaque pied en phase d’appui. Enfin, lors de la terminaison de la marche, le corps cherchera plutôt à faire retourner le CM à l’intérieur de la base de support afin de retrouver sa position d’équilibre (5,7).
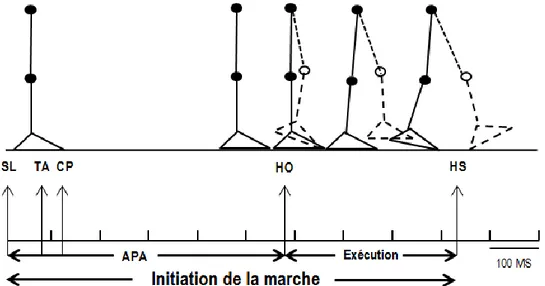
Le programme moteur est un groupe de commandes musculaires structurées ayant pour but précis de créer un mouvement volontaire (31). Les premières études sur l’initiation de la marche ont permis de détecter, avec les enregistrements électromyographiques (EMG), l’activation de certains groupes musculaires bien avant que le premier mouvement de marche ne soit observé (3,4,9). Tout d’abord, une inhibition de l’activité tonique du soléaire précède de 75 à 100 ms, une activation bilatérale du muscle tibial antérieur (TA) (3,8,9).
L’initiation de la marche peut donc se diviser en deux phases (Figure 3): la phase d’anticipation et la phase d’exécution (32). La première phase qui précède tout mouvement du corps commence par une inhibition des soléaires et se termine au décollement du talon (HO) de la jambe oscillante et peut varier entre 455 ms et 669 ms. Lors de cette phase, un phénomène dynamique est déjà mis en place puisqu’un déplacement du CP et du CM est observé (32). La seconde phase (Exécution) se termine au contact du talon (HS) de la jambe oscillante (1,3,33). La nomenclature utilisée pour l’appellation des membres inférieurs lors de l’initiation de la marche réfère à Mann et coll. (4) qui définissent la jambe d’initiation du
mouvement (celle qui quitte le sol) comme étant la jambe oscillante alors que l’autre membre inférieur demeurant au sol, se nomme jambe d’appui.
Figure 3: Les ajustements posturaux anticipatoires (APA) débutent par l’inhibition des
soléaires (SL) suivis de l’activation des tibiaux antérieurs (TA) et se terminent au décollement du talon (HO). Le centre de pression (CP) commence son déplacement de 100 à 200 ms après l’inhibition des SL. La phase d’exécution commence à HO et se termine par
le contact du talon (HS).
3.1 - La phase d’anticipation
Tout d’abord, l’inhibition des soléaires suivie par l’activation bilatérale des TA créent une diminution du moment de plantiflexion aux chevilles, tandis que les abducteurs de la hanche de la jambe oscillante créent plutôt une surcharge de celle-ci (5,7,8). La combinaison de l’activation des muscles de la cheville et de la hanche déplacent respectivement le CP en postérieur et en latéral vers la jambe oscillante (3–5,7), soit 100 à 200 ms après l’inhibition des soléaires (Figure 4). Pendant que le CP se déplace dans l’axe antéro-postérieur, le CM subit une chute vers l’avant et vers la jambe d’appui (3,7,32). Lorsque le CP atteint son déplacement maximal vers la jambe oscillante, une augmentation de la force de réaction verticale au sol surcharge la jambe oscillante, lui permettant d’exercer une poussée contre le sol dans le sens opposé (6,7). C’est alors que le corps amorce son premier mouvement de marche en décollant le talon du sol (33).
11
Figure 4: Trajectoire du centre de pression (COP) et du centre de masse (COM) lors de
l'initiation de la marche avec la jambe droite. Alors que le centre de pression se déplace en postérieur vers la jambe oscillante, le centre de masse amorce une chute vers l'avant et vers
la jambe d'appui (5).
À la fois l’inhibition des soléaires et l’activation des TA sont responsables du déplacement arrière du CP. Cependant, le déplacement du CP dans l’axe antéro-postérieur est principalement dû à l’activation du TA, où une forte corrélation existe entre les deux composantes (9). Crenna et Frigo (9) ont observé une séquence complète du patron moteur bilatéralement dans 81 % de leur essai d’initiation de la marche à vitesse naturelle alors que dans le 19 % restant, la séquence était observée unilatéralement. À vitesse de marche supérieure, la séquence devenait progressivement de plus en plus apparente bilatéralement, jusqu’à l’obtention du patron dans 100% des essais. Chez Polcyn (12), la séquence complète du patron moteur a plutôt été classée selon sa manifestation dans la jambe oscillante et la jambe d’appui. Respectivement, la séquence était observée dans 68 % et 76 % des essais chez leur groupe de jeunes adultes. Un nombre considérable d’essais avaient alors été retirés des analyses, soit 140 (sur 541) et 159 (sur 452) pour la jambe oscillante et la jambe d’appui, respectivement. Les essais étaient inclus seulement si le soléaire était actif et le TA inactif au moment de l’apparition du signal de départ. Le délai entre l’inhibition du soléaire suivi de l’activation du TA peut différer selon les études. Cet écart
est probablement due au critère de détection de l’inhibition des soléaires (34). Chez Crenna et Frigo (9) le délai observé était de 92 ± 29 ms et le début de l’inhibition du soléaire était utilisé pour déterminer le délai entre l’inhibition du soléaire et l’activation du TA. Chez Brunt et coll. (34), l’inhibition complète du soléaire était utilisée et le délai était de moins de 60 ms. L’activation du TA de la jambe d’appui surviendrait avant celle de la jambe oscillante (8).
3.2 - La phase d’exécution
Après le décollement du talon de la jambe oscillante, le CP continu sa trajectoire en se déplaçant rapidement sous la jambe d’appui dans l’axe antéro-postérieur. Un rapide transfert de poids de la jambe oscillante vers la jambe d’appui s’effectue ensuite lors du dégagement des orteils au sol de la jambe oscillante (4,35). Le CP termine finalement sa course sous le pied d’appui lorsque la jambe oscillante reprend contact avec le sol (HS) (3,4,33). De son côté, le CM continue son accélération vers l’avant et vers la jambe d’appui pour ensuite se rediriger vers la direction de marche désirée (Figure 4) (3,5,7,32). Les APAs créent donc des conditions dynamiques qui permettent au corps de se lancer dans un processus de marche. L’initiation de la marche ne peut commencer sans un découplage du CP et du CM. Le but est de créer une chute contrôlée avant du CM et d’effectuer un transfert de poids vers la jambe d’appui afin que la jambe oscillante puisse quitter le sol (7).
3.2 - Vitesse et direction
L’initiation de la marche a aussi pour but de placer le corps dans un processus de marche stable et ce, dans un temps invariable (1). Mann et coll. (4) et Miller (35) ont établi que les paramètres de la marche stationnaire étaient atteints à la fin du troisième pas alors que chez Brenière (1), la cadence et la vitesse du CM se stabilisait plutôt à l’intérieur du premier pas. Finalement, Jian et coll. (7) ont observé les paramètres stables de la marche après le deuxième pas seulement. Selon Brenière (1), la vitesse du CM, nulle en position statique de départ, augmente et atteint un sommet à la fin du premier pas. Selon Jian (7), avant le contact du talon, le CM a atteint une trajectoire proche de celle que l’on retrouve lors de la marche stabilisée. De plus, c’est 90 % de la vitesse de croisière qui est atteinte à l’intérieur du premier pas, pour finalement atteindre une stabilité à la fin du deuxième pas. Cependant, en recevant comme instruction de marcher à une vitesse rapide, les paramètres de la marche
13
n’étaient pas stabiliser à l’intérieur du premier pas (1). Dans cette condition de marche rapide, la vitesse se stabiliserait alors plutôt à l’intérieur du deuxième ou troisième pas (4,7). Un facteur qui pourrait expliquer ces différences, du moins en partie, seraient la dominance du membre inférieur (voir section 3.4). Alors que l’étude de Brenière (1) n’imposait pas une jambe de départ pour initier le premier pas, la jambe de départ droite était imposée aux sujets des autres études (4,7) et peut expliquer les différences dans la stabilisation de la marche au cours du processus, puisque la jambe droite n’était pas nécessairement la jambe préférentielle de départ ce qui aurait pu modifier l’exécution du premier pas (1).
Certaines variations dans les APAs et dans la phase d’exécution sont observées lorsqu’une personne reçoit des instructions concernant sa future vitesse de marche. Pour ce qui est du délai entre l’inhibition du soléaire et l’activation du TA, celui-ci ne serait pas affecté par une augmentation de la vitesse de marche (34). Cependant, Crenna et Frigo (9) ont observé une diminution de l’intervalle soléaire et TA lors des essais de marche rapide. Des modifications seraient plutôt observées dans la durée des deux phases qui démontre des covariations avec une augmentation de la vitesse de marche. Alors que la durée des APAs augmente avec la vitesse de marche, la phase d’exécution diminue. Bien que ces changements de temps surviennent dans chacune des phases, la durée de l’initiation de la marche demeure indépendante de la vitesse de progression (32). Ensuite, le déplacement du CP lors des APAs est en covariation avec l’augmentation de la vitesse de marche. Le CP se déplace à de plus grandes amplitudes avec une augmentation de la vitesse. Ce déplacement est toutefois limité à l’intérieur de la base de support et ne peut aller au-delà. L’accélération du CM est donc par le fait même, elle aussi limitée. Afin d’atteindre des vitesses de marche supérieures, la position initiale des pieds devra être modifiée comme déplacer davantage le pied de la jambe oscillante vers l’arrière (32). La figure 5 illustre l’amplitude du déplacement du CP selon la future vitesse de marche.
Figure 5: Déplacement du centre de pression (CP) selon la vitesse de marche lors d'un
départ avec la jambe gauche (F=rapide, N=normal, S=lent). Le CP se déplace à de plus grandes amplitudes dans l'axe antéro-postérieur avec une augmentation de la vitesse (32).
Les études antérieures sur l’initiation de la marche se sont davantage concentrées sur les caractéristiques des APAs et de la phase d’exécution lorsque les sujets employaient une direction avant et non lors de départs impliquant un changement de direction. Corbeil et Anaka (36) ont démontrés les modifications de la phase d’APAs lorsque l’initiation de la marche nécessite une rotation du corps pour se diriger à l’endroit voulu. L’amplitude des ajustements posturaux est modifiée selon l’amplitude du changement de direction. De plus, en initiant la marche dans le sens horaire, la vitesse du CM à HO est diminuée alors qu’elle augmentait lors d’un départ dans le sens antihoraire. Lors de cette dernière condition, les déplacements latéraux étaient augmentés. Les composantes antéro-postérieures des APAs lors de l’initiation de la marche permettraient donc d’ajuster la vitesse de marche désirée alors que les composantes médiolatérales créent les conditions de marche en lien avec un changement de direction horaire ou antihoraire (36).
3.3 - Longueur et largeur du premier pas
La longueur du premier pas se calcule comme étant la distance du talon de la jambe oscillante dans le sens antéro-postérieur entre sa position initiale et son contact avec le sol après la phase d’oscillation (37). La longueur du premier pas varie d’une étude à l’autre. Mbourou et coll. (38). ont étudié la variabilité de la longueur de pas chez de jeunes adultes,
15
des personnes âgées saines et des personnes âgées chuteuses. La longueur moyenne du premier pas observée chez leur groupe de jeunes adultes était de 0,745 m et la longueur du deuxième pas (première foulée) était ensuite d’environ 1,5 m. Peu de variations étaient observées dans la longueur des foulées après le premier pas chez cette population. Delval (33) a ensuite observé une longueur de pas de 0,70 ± 0,10 m lors d’une initiation de la marche en condition auto commandée. La longueur du deuxième pas était ensuite de de 1,44 ± 0,17 m. D’un autre côté, Couillandre et coll. (39) ont démontrés la variation de la longueur du premier pas en lien avec l’augmentation de la vitesse de marche. Pour une vitesse préférentielle du sujet, la longueur du premier pas était de 0,67 ± 0,08 m alors qu’elle augmentait à 0,75 ± 0,10 m pour une vitesse rapide de marche.
La largeur du premier pas quant à elle est la distance en médiolatéral du talon de la jambe oscillante entre sa position initiale et sa position à la fin du premier pas (37). La largeur du premier pas serait d’environ 0,12 m chez de jeunes adultes avec la jambe préférentielle de départ (37,40)
3.4 - Jambe préférentielle versus jambe non-préférentielle
Peters et Durding (41) se sont intéressés à la prévalence de la latéralité des membres inférieurs selon la dominance manuelle. Selon leurs résultats, 95% des droitiers manuels préféraient la jambe droite lors d’une tâche de coup de pied alors que 50 % des gauchers préféraient la jambe gauche. L’influence de la latéralité des membres inférieurs lors de tâches motrices a jusqu’à maintenant été moins étudiée que celle des membres supérieurs. Néanmoins, celle-ci amène des modifications dans les paramètres de marche (37) de même que la latéralité des membres supérieurs peut avoir une influence sur la régularité des mouvements des mains et des bras (42).
La jambe préférentielle est le membre inférieur utilisé pour manœuvrer et diriger. À l’opposé, la jambe non-préférentielle joue plutôt un rôle de stabilisation et de support pour la jambe préférentielle (41). Hart & Gabbart (43) ont étudié le rôle de stabilisation de la jambe préférentielle et non-préférentielle dans un contexte de stabilité unilatérale et bilatérale. Ils en sont arrivés à la conclusion que le choix de la jambe préférentielle dépend de la nature de la tâche ainsi que de sa complexité. Dans cette étude, la majorité des sujets
qui avaient pour jambe préférentielle le membre inférieur droit utilisait la jambe gauche pour se stabiliser dans un contexte de stabilité bilatérale. Lors d’une tâche de stabilité unilatérale la jambe droite était plutôt préférée. Cet aspect était inversement observé chez les sujets présentant une jambe préférentielle gauche.
Les études précédentes sur l’initiation de la marche n’imposaient pas aux sujets un membre inférieur de départ (1,3,9,32,33), ou donnaient comme instruction d’initier le mouvement avec la jambe droite (4,7). L’initiation de la marche selon la jambe préférentielle et non-préférentielle amène une asymétrie dans les paramètres de la marche. Entre autre, des variations dans la trajectoire du CM ainsi que dans la largeur du premier pas ont été observés selon ses deux conditions de départ (37). Tout d’abord, un départ avec la jambe non-préférentielle, amène le CM à employer une trajectoire plus médiale du décollement des orteils (TO) jusqu’au contact du talon (HS) de la jambe oscillante. Ensuite, la largeur du pas est plus grande dès le dépôt du talon de la jambe oscillante. La figure 6 illustre les variations en médiolatéral de la trajectoire du CM et de la largeur du premier pas dans les deux conditions. Selon Dessery et coll. (37), ces modifications dans le plan médiolatéral reflètent les besoins de stabilisation posturale du corps lorsque la jambe non-préférentielle initie la marche versus la jambe préférentielle.
Figure 6: Variations en médiolatéral de la trajectoire du centre de masse et de la largeur du
17
3.5 - Démarrage déclenché versus auto commandé
Deux processus différents de contrôle sont impliqués dans des mouvements initiés volontairement et des mouvements réactifs suite à un stimulus. Les deux types de mouvements sont modulés par des aires motrices non primaires. Le cortex moteur et le cortex prémoteur latéral seraient plutôt impliqués dans les mouvements initiés par un stimulus externe alors que les mouvements initiés volontairement seraient en lien avec l’aire motrice supplémentaire (44).
Delval (33) s’est intéressé aux mouvements auto commandés et déclenchés sur la durée des APAs lors de l’initiation de la marche chez des sujets sains. Selon sa définition, une condition déclenchée est un démarrage rapide de la marche suite à un signal visuel ou sonore, tandis qu’une condition auto commandée est un démarrage rapide du sujet selon sa propre initiative. Les résultats ont démontré des différences dans la durée des APAs et dans la vitesse d’exécution du premier pas. Tout d’abord, en condition auto commandée, la vitesse du premier pas plus rapide exprime une durée des APAs plus longue dans cette condition. En effet, Brenière (32) a démontré des covariations entre la durée des APAs et la vitesse de la phase d’exécution du premier pas.
Chez les individus atteint de la maladie de Parkinson, le cortex moteur fonctionne normalement (45). Cependant, la maladie amène chez eux une altération de l’activation de l’aire motrice supplémentaire et du putamen controlatéral (46). Des symptômes de brakinésie et d’akinésie, plus prononcés lors de production de mouvements auto-commandés, sont observés chez cette population en plus de symptômes de tremblements, de rigidité et d’une détérioration de l’équilibre (47). Il a été observé que les symptômes de brakinésie et d’akinésie sont moins prédominants lors de la performance de mouvements déclenchés par indices sensoriels (47). L’utilisation d’un indice sensoriel, pour déclencher des actes moteurs, chez cette population possède donc une importante implication pour des activités de transition comme l’initiation de la marche (33,47).
3.6 - Inclinaison du tronc
Le tronc représente plus de 50 % de la masse total du corps entier (48). Son rôle principal lors de la marche est de maintenir la tête en équilibre et d’assister les membres inférieurs
dans la génération du mouvement vers l’avant (49). Lors de l’initiation de la marche, le tronc permet de préserver la progression vers l’avant du corps suite au premier levé du talon (50).
Leteneur (51) a étudié l’effet d’une inclinaison du tronc avant et arrière sur le moment articulaires lors de l’initiation de la marche. L’angle au tronc était mesuré entre la ligne joignant les trochanters et les acromions et la verticale. Les inclinés vers l’arrière démontraient un moment de flexion à la hanche supérieur avant HO. L’inclinaison du tronc n’influençait toutefois pas les paramètres spatio-temporels de l’initiation de la marche. Lepers et Brenière (52) ont observés une inclinaison du tronc vers l’avant précédant HO afin d’accélérer la chute avant du CM spécifiquement lors d’essais de marche rapide. Ceci aurait pour but de favoriser l’action de la gravité lors de l’initiation de la marche.
Lors du processus de marche, le tronc est initialement incliné vers l’avant de 6º par rapport à la position debout neutre et peut augmenter jusqu’à 13º avec une augmentation de la vitesse (48). Lors de l’adoption d’une posture du tronc incliné vers l’avant lors de la marche, des stratégies compensatoires au niveau de la cinétique des membres inférieurs seraient nécessaires (53). Leteneur (54) a aussi étudié l’effet de l’inclinaison naturelle lors de la marche en utilisant la même classification d’inclinaison du tronc que l’étude précédente sur l’initiation de la marche. Les sujets inclinés vers l’arrière se propulsaient avec un moment de flexion à la hanche supérieur lors de la phase de propulsion et les sujets inclinés vers l’avant maintenaient un moment d’extension à la hanche plus long tout au long de la phase d’appui.
Certaines modulations de la séquence du programme moteur existent lors de changements de la posture initiale de l’initiation de la marche. En initiant la marche avec une posture exagérément inclinée vers l’avant, amenant le CP à prendre une position plus antérieure à l’axe des chevilles, la synergie entre l’inhibition du soléaire suivie de l’activation du TA est diminuée (9). Ceci est principalement dû à une diminution de l’amplitude des TA, bilatéralement. L’inhibition du soléaire était quant à elle diminuée en amplitude seulement au niveau de la jambe oscillante. Edamura (55) a aussi démontré des résultats similaires où une inclinaison initiale du tronc exagéré vers l’avant modifiait l’amplitude de l’activation
19
l’instruction de s’incliner vers l’avant, en déplaçant le CP en antérieur (9) ou de s’appuyer sur le dossier d’une chaise devant eux permettant une posture exagérée inclinée vers l’avant (55) lors de la position debout précédant l’initiation de la marche. L’inclinaison naturelle du tronc, mesuré par angle segmentaire, et son effet sur l’expression du programme moteur propre à l’initiation de la marche n’a toutefois pas été investigué. L’étude de Leteneur et coll. (51) ont observé une différence entre une inclinaison avant et arrière du tronc sur les moments musculaires lors de l’initiation de la marche, mais leur résultat ne démontraient pas l’effet de cet inclinaison naturelle sur la séquence d’inhibition des soléaires et d’activation des TA en APAs.
4 - Initiation de la marche et vieillissement
Selon l’OMS, l’âge chronologique pour définir une personne âgée est fixé à plus de 60 ans. Le vieillissement est un phénomène planétaire dû à une baisse des taux de fécondité et à l’amélioration des soins de santé
Les troubles de la marche augmentent avec l’âge. À 60 ans, 85 % des gens démontrent une marche normale alors que ce pourcentage diminue à 18% à l’âge de 85 ans (56,57). L’une des principales conséquences découlant de cette réalité est le risque de chute qui augmente avec l’avancement en âge. (58). Près de un tiers des personnes âgées de 65 ans et plus et 50% des plus de 80 ans chutent par an(59,60). Au Canada, les chutes occupent le premier rang des causes d’hospitalisation chez les personnes âgées. 95 % des fractures à la hanches sont dues aux chutes et 20% des aînés ayant subi ce type de fracture décède dans l’année suivante (59).
Ce phénomène multifactoriel, est en partie dû aux modifications physiques, sensorielles et cognitives liées au vieillissement. Les changements moteurs reliés au vieillissement, comme les difficultés de coordination, l’augmentation de la variabilité du mouvement, la lenteur de mouvement et les difficultés au niveau de l’équilibre et de la marche sont dus aux dysfonctionnements du système nerveux central et périphérique (61,62).
Une diminution du volume du cerveau chez les personnes âgées comparativement à de jeunes adultes pourrait être reliée aux déficits moteurs observés chez les aînées(61,62). Les régions préfrontales et pariétales sont les plus susceptibles à une diminution de la matière
grise dans le cerveau (63–65) et le contrôle moteur est plus dépendant de ces régions du cerveau (62). L’atrophie du cortex somatosensoriel aurait aussi une implication dans le risque de chute et dans la diminution du contrôle de l’équilibre chez les personnes âgées (62) (66). Ensuite, le cervelet qui est responsable du contrôle de la posture et de la coordination des mouvements volontaires subit aussi les effets du vieillissement. Une étude longitudinale (67) s’étendant sur une période de cinq ans, a démontré que le noyau caudé et le cervelet présentaient une grande diminution de volume au cours de ces années. Ensuite, le système dopaminergique du striatum diminue avec l’avancement en âge (68). Une association existe entre le niveau de dopamine dans le striatum et l’équilibre et la marche (62). Selon Cham (68), la diminution de la dopamine dans le striatum est associé à l’augmentation de l’amplitude des oscillations posturales dans l’axe antéro-postérieures. Au niveau de la marche, cette diminution serait aussi corrélée aux paramètres caractérisant le vieillissement comme la diminution de la vitesse, de la cadence et une augmentation de la durée de double-support (62,68).
Au niveau du système musculo-squelettique, le vieillissement est associé à une diminution graduelle et inévitable de la masse musculaire (69). Cette atrophie musculaire débute dès l’âge de 50 ans et à 80 ans c’est 50 % des fibres musculaires des membres inférieurs et supérieurs qui sont perdues. La diminution de la masse musculaire est accompagnée d’une diminution de la force. Ces effets du vieillissement sur le système musculaire ont donc un important impact sur les activités de la vie quotidienne des personnes âgées.
Il est donc évident que la dégénérescence des structures au niveau du système nerveux central, périphérique et neuromusculaire ne peut que soumettre les personnes âgées l’instabilité. L’initiation de la marche étant un défi au niveau du contrôle postural (40,70), il est suggéré dans la littérature que cette tâche fonctionnelle sous contrôle volontaire, puisse augmenter le risque de chutes des personnes âgées chez qui le processus présente des modifications par rapport à de jeunes adultes (12,13,38,40).
21
4.1 - La phase d’anticipation
Le programme moteur utilisé chez les personnes âgées pour initier la marche ne diffère pas de celui que l’on rencontre chez des sujets sains en bonne santé. Cependant, la fréquence du patron moteur typique à l’initiation de la marche est diminuée chez une population vieillissante. Le muscle soléaire n’est pas toujours inhibé avant l’activation du TA et la marche serait initiée avec moins d’anticipation du TA (12,13).
Polcyn et coll. (12) ont démontré que le patron stéréotypé d’initiation de la marche était diminué chez les personnes âgées, où le soléaire n’était pas toujours inhibé avant l’activation du TA. La séquence complète du patron moteur était observée dans 49 % des essais au niveau de la jambe d’appui et dans 24 % des essais au niveau de la jambe oscillante. Chez les jeunes adultes, la fréquence d’utilisation du patron était plus grande, mais n’atteignait toutefois pas des pourcentages élevés (76 % pour la jambe d’appui et 68 % pour la jambe oscillante).
Alors que Polcyn et coll. (12) ont mis en évidence une diminution de la fréquence du patron moteur principalement due à l’absence de l’inhibition du soléaire chez les personnes âgées, Henriksson et coll. (13) ont observé des changements au niveau de la composante anticipatoire de l’activité du TA. Chez les jeunes adultes, l’activation du TA de la jambe oscillante était faite en anticipation (en APAs) alors que huit des sujets âgés ne démontraient pas d’activation du TA anticipée. La durée de l’activation du TA en APAs était aussi réduite chez les sujets âgés. Pour ce qui est de la jambe d’appui, les différences étaient moins marquées.
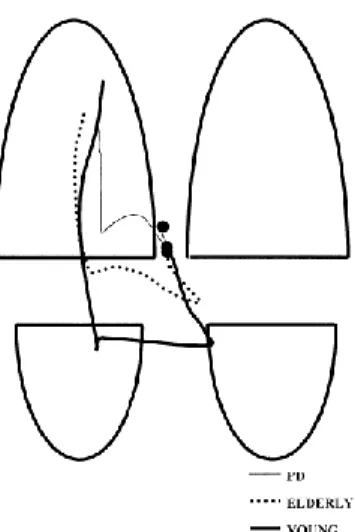
Ces modifications dans l’activation des muscles de la cheville en APAs résultent en une diminution de l’amplitude du déplacement du CP en postérieur. En début d’APAs, le CP se situe à la même position chez les sujets jeunes et âgés, soit au point milieu entre les deux pieds, antérieur à l’axe intermaléollaire. Lorsque le CP et le CM se dissocient, le CP se déplace davantage en postérieur chez les sujets jeunes comparativement à leurs homologues plus âgés (Figure 7) (40). Cela a donc pour résultat d’augmenter la vitesse de marche, d’augmenter la longueur du premier pas et d’augmenter la durée des APAs.
Figure 7: Déplacement du centre de pression lors de l'initiation de la marche chez de
jeunes adultes et des personnes âgées (40).
La capacité de générer une propulsion du corps vers l’avant par le déplacement antéro-postérieur du CP est donc significativement diminuée chez les sujets âgés (71). C’est-à-dire que pour un même déplacement du CP en postérieur, les personnes âgées vont créer un moment de force plus petit vers l’avant. Celles-ci devraient donc utiliser des stratégies autres que le déplacement du CP en postérieur lors des APAs pour se mettre en processus de marche (12,13). Des adaptations au niveau du haut du corps sont plutôt utilisées puisque le seul déplacement arrière du CP n’est pas suffisant à projeter le CM au-delà de la base de support. Laudani et coll. (71) ont observé un déplacement angulaire du haut du corps qui était supérieur en APAs chez cette même population. En déplaçant le haut du corps vers l’avant et vers la jambe d’appui, le centre de masse peut être projeté à l’extérieur de la base de support et ainsi générer une chute du corps vers l’avant (12). Il a été suggéré que cette stratégie pourrait amener de l’instabilité comparativement à une stratégie du déplacement du CP qui, quant à elle, permet plutôt de garder le haut du corps à l’intérieur de la base de support (12,72).
Des différences au niveau des forces de réactions au sol sont aussi observées chez des sujets âgés lors des APAs (6). Une initiation typique de la marche présente une augmentation des forces verticales au sol sous le pied de la jambe oscillante avant le levé du talon suivi d’un rapide transfert de poids vers la jambe d’appui lorsque le pied décolle du sol (6,7). Ce sont ces variations de forces verticales entre la jambe oscillante et la jambe
23
d’appui qui favorisent le passage d’une position de double support à simple support (6). Chez les sujets âgés, des modifications dans la répartition des forces de réactions verticales au sol dans la position de départ amènent des difficultés dans le passage d’une position de double-support à simple support. Chez de jeunes adultes, le poids du corps est également réparti entre les deux membres inférieurs lors de la position de départ (6,7) alors qu’il est davantage réparti sur la jambe d’appui chez les sujets âgés (13). Selon Henriksson (13), cette position initiale de marche chez les sujets âgés, serait une stratégie adoptée inconsciemment dans le but de répondre à une quelconque instabilité (73).
4.2 - La phase d’exécution
Selon Patchay (6), une phase d’ajustements posturaux anticipatoires qui est accomplie convenablement permet une exécution adéquate du premier pas. Les modifications dans la phase d’APAs chez les sujets âgés amènent donc par le fait même des changements dans la phase d’exécution. Les paramètres de marche comme la vitesse du CM, la durée de la phase d’exécution, la longueur du premier pas et sa variabilité chez une population vieillissante saine et pathologique ont été mis en évidence dans les études antérieures (38,40). Tout d’abord, Halliday et coll. (40) ont comparé l’initiation de la marche chez trois populations distinctes, soit chez de jeunes adultes, des personnes âgées en bonne santé et des parkinsoniens. Dans cette étude, les personnes âgées démontraient une vitesse de leur CM globale significativement plus lente en début de phase d’exécution (levé du pied). Les différences dans la phase d’exécution entre les deux populations se situent davantage au niveau de la longueur de pas qui démontre une certaine variabilité (38,40). Dans l’étude de Halliday (40), la longueur du premier pas était significativement plus grande chez le groupe de jeunes adultes que chez le groupe de sujets âgés. L’étude de Mbourou et coll. (38), s’est intéressée à la variabilité de la longueur du premier pas chez des personnes âgées non-chuteurs et non-chuteurs. Dans cette étude, les deux groupes de personnes âgés démontraient une longueur du premier pas plus courte que le groupe de jeunes adultes. Alors que les jeunes adultes avaient en moyenne une longueur du premier pas de 745 mm, celle des sujets âgés non-chuteurs était plutôt de 525 mm versus 295 mm pour les chuteurs. Cette étude a aussi fait ressortir la variabilité du premier pas qui est un facteur prédisposant au risque de chute. La variabilité était de 56 mm dans le groupe de jeunes adultes comparativement à 57 mm dans le groupe d’individus âgés non-chuteurs où la différence de
résultat n’était pas significative. Par contre, chez les âgés chuteurs, cette variabilité était significativement plus grande, elle était de 125 mm De plus, le temps passé en double support, suite au premier pas, était plus grand chez les individus âgés chuteurs. Selon Maki (74), la variabilité dans la longueur de la foulée, la vitesse, le temps de double-support et l’augmentation de la largeur de pas sont associés au risque de chute chez les personnes âgées. Les changements dans les paramètres de marche telle que la diminution de la vitesse de progression, de la longueur de pas et l’augmentation de la période de double-support seraient des stratégies de stabilisation utilisées en lien avec la peur de chuter. Cette étude apporte comme élément l’importance de la variabilité entre les foulées lors de la marche et la définit comme étant un fort prédicteur du risque de chute.
Selon Brenière (1), le processus de marche est atteint à l’intérieur du premier pas chez de jeunes adultes. Étant donnée l’augmentation de la variabilité dans la marche chez les sujets âgés chuteurs, la marche ne peut se stabiliser à l’intérieur du premier pas comme il a été démontré chez de jeunes adultes. Lindemann (75) a déterminé la distance ou le nombre de cycles de marche nécessaires pour atteindre ce même processus chez des personnes âgées frêles. Celles-ci nécessitaient en moyenne une distance de 2,5 m pour stabiliser leur marche. Les quatre premiers cycles de marche incluent dans cette distance démontraient beaucoup de variabilités au niveau de la vitesse de marche et de la longueur des foulées. Lindemann suggère donc d’augmenter la distance de prise de mesure lors de l’analyse de la marche chez une population d’individus âgés frêles afin que ceux-ci puissent stabiliser leur marche.
5 - La variabilité
La variabilité intra individuelle réfère à des changements réversibles à court terme dans la performance d’une personne lors d’une seule tâche ou lors d’une tâche répétée plusieurs fois à l’intérieur d’un court intervalle de temps. Il est suggéré qu’il y a un lien entre la variabilité intra individuelle et les dysfonctions neurologiques (76). Au niveau de la marche, il est reconnu traditionnellement que le système génère une marche stable et constante d’une foulée à l’autre et que la variabilité dans les paramètres de marche est plus présente chez des individus en moins bonne santé. Cependant, le patron de marche
25
stable. D’un autre point de vue, au lieu d’être vu comme nuisible au système et à la performance, la variabilité serait plutôt inhérente au système et aiderait l’individus à s’adapter aux différentes contraintes de la tâche, de l’individus et de l’environnement (14). Considérée dans le passé comme étant du bruit et éliminée des analyses en tant que source d’erreur, la variabilité donnerait plutôt d’importantes informations dans la compréhension du contrôle moteur de la marche (77).
Certaines modulations ont déjà été ciblées expliquant la variabilité du patron EMG de l’initiation de la marche chez de jeunes adultes. Tout d’abord, la future vitesse de marche influence l’amplitude de l’activation du TA (9,32). Ensuite, en demandant aux participants de s’incliner volontairement vers l’avant, on observe une diminution en amplitude de l’inhibition des soléaires et de l’activation des TA caractérisant l’initiation de la marche (9). L’effet de l’inclinaison naturelle du tronc et son effet sur la séquence prédéfinie de l’activité EMG n’a toutefois pas encore été étudié. Une inclinaison naturelle du tronc davantage vers l’avant pourrait en partie expliquer l’absence d’inhibition des soléaires que l’on observe chez de jeunes adultes. De plus, d’autres conditions de posture initiale caractérisées par l’inclinaison du corps, l’activation EMG initiale et la présence d’une co-activation des muscles de la cheville, pourraient aussi expliquer une partie de la variabilité que l’on observe dans ce programme moteur, chez une population de jeunes adultes.
Le prochain chapitre présente l’étude qui a été réalisée afin de comprendre davantage les sources de variation, incluant notamment l’inclinaison naturelle du tronc, du patron d’activation musculaire des soléaires et du TA lors des APAs précédant l’initiation de la marche.
27
Résumé - Chapitre2
Objectif : L’initiation de la marche est la transition entre la posture debout et le processus
de marche stabilisé mettant en place une phase d’ajustements posturaux anticipatoires (APAs) suivie d’une phase d’exécution du premier pas. Les APAs sont caractérisés par un patron électromyographique structuré mettant en place l’inhibition des soléaires suivie de l’activation bilatérale des muscles tibiaux antérieurs (TA). Ce patron est variable chez une population de personnes âgées et chez de jeunes adultes, mais à de moindre fréquence chez ces derniers. Les jeunes adultes représentent une population en bonne santé sans dysfonctionnement neurologique. La présence du patron d’activation musculaire devrait donc est observé dans la majorité des essais chez cette population. La posture initiale en position debout précédant l’initiation de la marche pourrait en partie expliquer la variabilité du programme moteur chez les jeunes adultes. Le but de cette étude est d’analyser l’effet d’une inclinaison naturelle du tronc sur l’inhibition des soléaires et d’activation du TA lors de l’initiation de la marche et de prédire l’inhibition des soléaire et l’amplitude d’activation du TA lors des APAs.
Méthode : Deux groupes de douze jeunes adultes ont été séparé en fonction de leur
inclinaison naturelle du tronc et ont complétés cinq essais d’initiation de la marche avec la jambe droite. Un modèle de régression a été utilisé afin de prédire l’inhibition des soléaires et l’activation du TA.
Résultats : Les inclinés vers l’arrière inhibaient le soléaire de la jambe d’appui moins
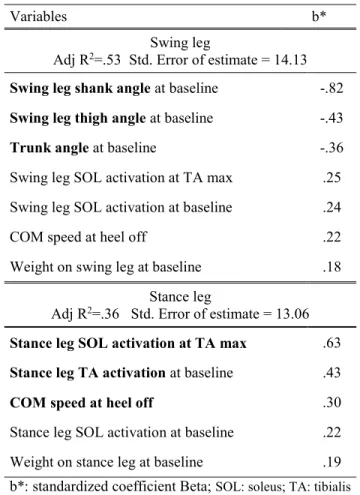
souvent (25,8% des essais) que les inclinés vers l’avant (55,6% des essais). Le modèle de régression montre qu’une activation initiale plus élevée du soléaire est la variable qui prédit le mieux la variation de l’inhibition du soléaire mais pas l’activation du TA. Peu de différences dans la cinématique étaient observées entre les deux groupes.
Conclusion : Peu de variations des APAs est due à l’inclinaison naturelle du tronc. Les
contributions à la variation seraient dues à la posture initiale, à la future vitesse de marche et à l’activation tonique initiale des soléaires. L’absence d’inhibition pourrait en partie être expliquée par l’inclinaison naturelle du tronc, où une inclinaison arrière amène une diminution de l’activité tonique du soléaire en position debout, précédant l’initiation de la
marche. L’inclinaison naturelle du tronc module le patron d’activation musculaire de l’initiation de la marche, particulièrement sur la fréquence d’inhibition des soléaires. Les effets pourraient aussi être dus à la variabilité inhérente et fonctionnelle qui dépend de la variation posturale, de la coordination musculaires et aussi du rôle des membres inférieurs.
29
Chapitre 2: Effect of natural trunk inclination on
variability in soleus inhibition and tibialis anterior
activation during gait initiation in young adults
Annie-Pier Fortin1, Yoann Dessery1, Sébastien Leteneur2-5, Franck Barbier 2-4, Philippe
Corbeil1
1 Université Laval, Département de Kinésiologie, Faculté de Médecine, 2300, rue de la
Terrasse, Québec, Qc, Canada G1V 0A6
2 Université Lille Nord de France, Lille, France
3 UVHC, LAMIH, Valenciennes, France
4 CNRS, UMR CNRS, 8201, Valenciennes, France
5 Centre de Rééducation Fonctionnelle La Rougeville, Saint-Saulve, France
Adresse de correspondance :
Philippe Corbeil, Ph. D.
Faculté de Médecine, Kinésiologie, Université Laval 2300, rue de la Terrasse (PEPS)
Québec, QC, Canada G1V 0A6 Fax. No : 418-656-2441
Telephone No. : 418 656-2131-5604
Annie-Pier Fortin
Faculté de Médecine, Kinésiologie, Université Laval 2300, rue de la Terrasse (PEPS)
Québec, QC, Canada G1V 0A6 Telephone No. : 418 656-2131 - 4920
Abstract
Background. Anticipatory postural adjustments (APAs) of gait initiation (GI) permit first
step execution. APAs are characterized by a structured pattern of soleus (SOL) inhibition followed by tibialis anterior (TA) activation. This pattern shows variability among young adults where SOL is not always inhibited before TA activation. Initial posture preceding GI could explain a part of this variability. The aim of the study was to investigate the effect of natural trunk inclination on APAs during GI.
Methods. Two groups of twelve subjects divided by natural trunk inclination angle
performed five gait initiation trials. A regression model was computed to predict SOL inhibition and TA activation.
Results. Backward leaners showed less SOL inhibition in stance leg (25.8% of trials)
compared to forward leaners (55.6% of trials). Regression model revealed that high tonic EMG activity in SOL in the stance leg is the variable that best explains SOL inhibition variation within trials but not TA activation.
Conclusion. Slight variations in APAs are due to natural trunk inclination but more
contribution in APAs is due to initial posture, future step speed and initial tonic soleus activity. Absence of SOL inhibition could be in part explained by natural trunk inclination, where the backward inclination leads to lower tonic SOL activity in quiet standing. These effects could be due to inherent and functional variability, which depend on postural variation, muscular coordination and limb roles.
31
1. Introduction
Gait initiation (GI), the transition from quiet standing to steady gait [1], is divided into two phases: anticipation phase and execution of the first step. According to Crenna and Frigo [2], gait initiation includes a basic single motor pattern that starts with inhibition of the soleus (SOL) and activation of the tibialis anterior (TA). Both generate the backward COP displacement, but TA burst is primarily responsible for this backward shift [2], which is positively correlated with speed at the end of the first step [3].
In most research in motor control, variability is traditionally equated with noise, detrimental to system performance and is typically eliminated from data as a source of error [4,5]. Techniques for reducing and eliminating both within- and between-participants variability have been used. For instance, trials were voluntarily discarded and trials were included only if the SOL and TA were respectively active and inactive prior gait initiation [2,6]. This indicates that SOL inhibition and TA activation pattern is not always observed in young adults, i.e., the SOL is not always inhibited before the TA is activated. Other authors propose that variability is inherent and functional, and that variability in movement is necessary for changes in the coordination of movement [4,7,8].
Some modulation of the motor pattern exists and can explain such variability. For example, initial posture involving an exaggerated forward inclination of the body during quiet standing prior to GI modulates the motor program by reducing both SOL inhibition and TA amplitude [2]. Furthermore, Leteneur and al. [9] have previously shown that natural forward (FW) leaning participants present smaller hip flexion moments than backward (BW) leaning participants just prior to heel-off in GI. They explained these results by suggesting a more efficient use of gravity in FW leaners to initiate gait. But natural trunk inclination and its effect on the EMG sequence during GI have not been investigated. Other postural characteristics, such as segmental angles and initial EMG activation, could explain variability in young adult. Effort must be made to understand movement control and coordination and the role of variability within that. This is important knowing that impairment in muscular agonist-antagonist synergy in both lower limbs could lead to abnormal patterns of propulsive forces and postural instability [10] .