INTERACTIONS ENTRE LE LOUP GRIS,
LE CARIBOU FORESTIER ET L’ORIGNAL
EN FORÊT BORÉALE AMÉNAGÉE
Thèse
Nicolas Courbin
Doctorat en biologie
Philosophiae Doctor (Ph.D.)
Québec, Canada
© Nicolas Courbin, 2013
Résumé
Ma thèse explore les changements dans les stratégies comportementales du loup gris (Canis lupus), de l‟orignal (Alces alces) et du caribou forestier (Rangifer tarandus caribou) en réaction à l‟aménagement de la forêt boréale. J‟ai étudié les facteurs qui déterminent la répartition spatiale des trois espèces en analysant leurs déplacements et leur sélection d‟habitat, puis en évaluant le jeu spatial prédateur-proie. Mon premier chapitre révèle que les différences de tactique de déplacements entre le prédateur et ses proies créent des probabilités relatives de rencontre asymétriques entre les espèces le long de leurs trajectoires de déplacement. Par exemple, compte tenu du temps passé dans les différents milieux, les caribous avaient un risque relativement élevé de rencontrer des loups dans les coupes en régénération, alors que les loups y avaient relativement peu de chance de rencontrer des caribous. Les loups croisaient les trajectoires des orignaux plus souvent que celles des caribous. Mon second chapitre démontre que l‟aménagement forestier influence le temps de résidence et les déplacements inter-parcelles du caribou et de l‟orignal en diminuant la connectivité de leur habitat. La théorie des graphes montre que, dans leur réseau de peuplements fortement sélectionnés par les proies, les deux proies évitaient les parcelles entourées de coupes et de routes. Les proies réduisaient également leur temps de résidence dans ces parcelles lorsqu‟elles devaient s‟y rendre en faisant de longs déplacements. Les loups sélectionnaient les parcelles de végétation les plus connectées à l‟intérieur du réseau de leurs proies, et cela plus fortement même dans les secteurs qui étaient largement utilisés par les proies. Mon troisième chapitre indique que de décembre à mai, la probabilité de cooccurrence du loup et du caribou est la plus forte en périphérie des secteurs de coupes, dans les massifs forestiers préservés pour la protection du caribou. Ainsi, il est avantageux de considérer les relations prédateur-proie pour élaborer des plans d‟aménagement efficaces à la protection des espèces animales. Ma thèse souligne l‟influence majeure des activités humaines sur les stratégies de sélection d‟habitat du loup, du caribou et de l‟orignal, sur la dynamique de leur répartition spatiale et, ultimement, sur le jeu loup-proie.
Abstract
My thesis explores the changes in behavioral strategies by the gray wolf (Canis
lupus), moose (Alces alces) and forest-dwelling caribou (Rangifer tarandus caribou), in
response to boreal forest management. I studied the factors that determine the spatial distribution of the three species, first by analyzing their movements and habitat selection, and then by assessing the predator-prey spatial game. My first chapter reveals that differences in movement tactics between the predator and its prey create asymetric relative probability of encounter between wolves and either of their prey along their movement paths. For example, given the time spent in the different land cover types along their paths, caribou had a relatively high risk of encountering wolves in regenerating cuts, while wolves had a relatively low probability of encountering caribou in these cuts. Overall, wolves intersected the paths of moose more often than those of caribou. My second chapter demonstrates that logging influences the residency time and inter-patch movement probabilities of caribou and moose by decreasing habitat connectivity. Graph theory showed that both prey species avoided network nodes (i.e., species-specific high-quality patches) surrounded by cuts and roads. Prey reduced their residency time in these nodes when they had to make relatively long inter-node movements to get there. Wolves selected the most connected nodes within their prey‟s network, a selection that was even stronger than the selection for areas of high utilization distribution of its prey. My third chapter shows that from December to May, the relative probability of co-occurrence between wolf and caribou was highest in the vicinity of cutblocks areas, in the forest blocks protected from harvesting for the conservation of caribou. The consideration of predator-prey spatial relationships is therefore essential to the development of efficient management plans aimed at species recovery. My thesis underscores the major impact of human activities on habitat selection strategies by wolves, caribou and moose, on the dynamics of their spatial distribution, and, ultimately, on the wolf-prey game.
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ... vii
Liste des tableaux ... xi
Liste des figures ... xv
Remerciements ... xxiii
Avant-propos ... xxvii
Introduction Générale ... 1
1. Les interactions prédateur-proie ... 1
1.1. Complexité trophique et interactions prédateur-proie ... 1
1.2. Compétition apparente et interactions prédateur-proie ... 2
2. Une approche comportementale ... 3
2.1. Importance des effets non létaux de la prédation ... 3
2.2. Le comportement animal : un garde-fou des changements populationnels ... 5
3. La répartition spatiale : une résultante comportementale ... 5
3.1. Le déplacement animal ... 6
3.2. La sélection d‟habitat ... 6
3.3. Évaluer la sélection d‟habitat et les déplacements... 8
3.4. Influence des activités humaines ... 9
4. Le jeu comportemental prédateur-proie ... 10
4.1. Le jeu spatial prédateur-proie ... 10
4.2. Jeu prédateur-proie et complexité trophique ... 13
4.3. Jeu prédateur-proie et perturbations humaines ... 14
5. Hétérogénéité spatiale, connectivité et relations prédateur-proie ... 15
5.1. Accessibilité, connectivité et décisions de déplacements ... 15
5.2. Connectivité et interactions prédateur-proie ... 17
5.3. Réseau spatial et théorie des graphes ... 18
6. Aménagement de la forêt boréale ... 20
7. Objectif et organisation de la thèse ... 21
8. Modèles d’étude : le caribou forestier, l’orignal et le loup gris ... 23
9. Aire d’étude : la forêt boréale aménagée à l’Est du Québec... 26
Chapitre 1. Multi-trophic resource selection function enlightens the behavioural game between wolves and their prey ... 29
Résumé ... 31
Abstract ... 33
1. Introduction ... 35
2. Material and methods ... 39
2.1. Study area ... 39
2.2. Telemetry data ... 40
2.3. Habitat covariates ... 41
2.4. Evaluation of wolf and prey habitat selection ... 41
3. Results ... 44
4. Discussion ... 52
4.1. On the use of a multi-trophic framework ... 52
4.2. Different outcomes of movement decisions between wolves and their prey in human-disturbed areas... 54
4.3. Hunting tactic of wolves ... 54
4.4. Different movement tactics between caribou and moose... 55
5. Conclusions ... 56
Chapitre 2. Logging-induced changes in habitat network connectivity shape behavioral interactions in the wolf-caribou-moose system ... 59
Résumé ... 61 Abstract ... 63 1. Introduction ... 65 2. Methods ... 68 2.1. Study area ... 68 2.2. Spatial data ... 68 2.3. Telemetry data ... 69
2.4. Caribou and moose habitat networks ... 69
2.4.1. Preliminary steps for building networks (Steps 1 and 2) ... 70
2.4.2. Species-specific habitat network ... 72
2.5. Movements of caribou and moose within their networks ... 74
2.5.1. Patch choice ... 74
2.5.2. Residency time and patch departure ... 77
2.6. Response of wolves to the network of their prey ... 78
2.6.1. Wolf habitat selection ... 78
2.6.2. Movement patterns of wolves ... 79
3. Results ... 81
3.1. Caribou and moose networks ... 81
3.1.1. Network implementation ... 81
3.1.2. Network typology ... 82
3.1.3. Patch choice ... 85
3.1.4. Residency time and risk of patch departure ... 91
3.2. Response of wolves to the network of their prey ... 94
3.2.1. Wolf habitat selection ... 94
3.2.2. Wolf movement patterns ... 97
4. Discussion ... 105
4.1. Connectivity and movement decisions of prey ... 105
4.2. Wolf response to prey spatial network ... 109
4.3. Management implications ... 111
5. Conclusion ... 111
Chapitre 3. Landscape management for woodland caribou: the protection of forest blocks influences wolf-caribou co-occurrence ... 113
2. Materials and methods ... 120
2.1. Study area ... 120
2.2. Telemetry data ... 122
2.3. Periods of the year ... 122
2.4. Scale and habitat characteristics ... 124
2.5. Snow model ... 126
2.6. Statistical analyses ... 128
2.7. Relative probabilities of co-occurrence between wolves and caribou ... 129
3. Results ... 130
3.1. Response of caribou and wolves to natural landscape ... 133
3.2. Response of caribou and wolves to forest management ... 133
4. Discussion ... 139
4.1. Similarities in habitat selection by caribou and wolves ... 139
4.2. Differences in habitat selection by caribou and wolves ... 140
4.3. Role of protected forest blocks ... 141
4.4. Implications for conservation ... 142
Conclusion Générale ... 145
1. Tactiques de déplacement et asymétrie du risque de rencontre dans le jeu loup-proie ... 146
1.1. Un nouveau cadre de travail pour évaluer les conséquences des tactiques de déplacements dans le jeu prédateur-proie ... 146
1.2. Ce que nous révèle l‟asymétrie des probabilités relatives de rencontre entre les espèces dans le jeu loup-proie ... 147
2. Le rôle de la connectivité de l’habitat dans les interactions loup-proie ... 150
2.1. Les décisions de déplacement des proies dépendent de la connectivité ... 150
2.2. La connectivité : un élément à considérer dans le jeu prédateur-proie ... 152
2.3. Limites de l‟étude de connectivité ... 153
3. Conservation du caribou forestier ... 155
3.1. Considérer les relations trophiques lors de l‟établissement des plans d‟aménagement forestier... 155
3.2. Implications pour la conservation et recommandations d‟aménagement ... 157
4. Contribution scientifique ... 159
5. Perspectives de recherche ... 162
Liste des tableaux
Table 1.1. Mixed-effects logistic regression models of habitat selection of seven wolves, 29
caribou, and 10 moose during the summer-autumn period (1 July to 14 November) in the Côte-Nord region, Québec (Canada), with their selection coefficients (β ± SE). Models were robust to cross-validation, as indicated by high mean Spearman rank correlations ( ). ... 47
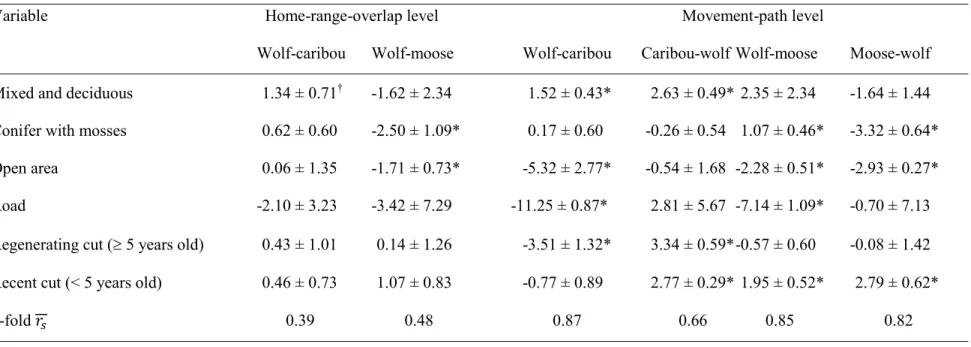
Table 1.2. Models of caribou (n = 197 sites, four wolves and six caribou) and
wolf-moose (n = 214 sites, two wolves and six wolf-moose) step intersections at the home-range-overlap (HRO) and movement-path (MP) levels during the summer-autumn period (1 July to 14 November) in the Côte-Nord region, Québec (Canada). The models were evaluated using mixed-effects logistic regressions based on a used/available design of RSF. Availability was evaluated (1) within the intersection of home ranges of both wolves and their prey (HRO level), and (2) along the movement path of each species (MP level), either along the predator path (Wolf-caribou and Wolf-moose) or along the prey path (Caribou-wolf and Moose-wolf). We show parameter coefficients (β ± SE). Models were robust to cross-validation, as indicated by relatively high . ... 48
Table 1.3. Percent (mean ± SE) of land cover type available within a 200-m radius buffer,
as determined from random locations drawn (1) within the home range of wolves, caribou and moose (HR level), (2) within the intersection between the home ranges of wolves and caribou or of wolves and moose (HRO level), and (3) along the predator paths (Wolf-caribou and Wolf-moose) or the prey paths (Caribou-wolf and Moose-wolf) (MP level), during the summer-autumn period (1 July to 14 November) in the Côte-Nord region, Québec (Canada). Availability was shown for wolves that were considered in wolf-caribou analyses or in wolf-moose analyses at HR and MP level. 49
Table 2.1. Mixed-effects logistic regression models of habitat selection of 24 caribou and
15 moose in winter, in the Côte-Nord region of Québec (Canada) with their selection coefficients (β) and associated 95% confidence intervals [CI], and the variance (VAR) and covariance (COV) for the random-effects. Availability was sampled within their annual 100% MCP for each individual. ... 83
Table 2.2. Candidate models explaining inter-patch movements of 23 caribou (n = 838
inter-patch movements) and 10 moose (n = 124 inter-patch movements) in winter in the Côte-Nord region of Québec (Canada), with the number of parameters (K), maximum log-likelihood (LL), quasi-likelihood under independence criterion differences (∆QIC), and QIC weight (wi). The top-ranked models are model 14 for caribou, and models 10 and 11 for moose. ... 86
Table 2.3. Best conditional logistic regression models explaining inter-patch movement
decisions of 23 caribou (n = 838 patch movements) and 10 moose (n = 124 inter-patch movements) during the winter in the Côte-Nord region of Québec (Canada). For each stratum, observed inter-node movements were compared to the set of alternative inter-node movements that could have been within the minimum planar graph from the same initial node. For caribou, the model is presented with its selection coefficients (β) and 95% confidence interval [CI] estimated from robust empirical
standard errors. For moose, the model is the average of the two top-ranked models (i.e., models 10 and 11, Table 2.2), and is presented with its model-averaged coefficients (β) and unconditional 95% confidence interval [CI] estimated from robust empirical standard errors. The of the k-fold cross-validation are indicated for the two models. ... 89
Table 2.4. Mixed-effects Cox models of predicting the risk of patch departure for caribou
(n = 23) and moose (n = 10) within network nodes, during the winter in the Côte-Nord region of Québec (Canada), based on the same model structure as for the best inter-patch movement model (Table 2.3). For caribou, the model is presented with its coefficients (β) and 95% confidence interval [CI] for the fixed-effects, and the variance (VAR) and covariance (COV) for the random effect, whereas, for moose, the average model is presented with its model-averaged coefficients (β) and unconditional 95% confidence interval [CI] for the fixed-effects, and the variance and covariance for the random-effect. ... 92
Table 2.5. Candidate models of habitat selection by seven wolves in four different packs in
winter in the Côte-Nord region, Québec (Canada), with the number of parameters (K), maximum log-likelihood (LL), Akaike information criterion differences (∆AIC), and AIC weight (wi). Model 5 is the top-ranked model. ... 95
Table 2.6. Mixed-effects logistic regression models of habitat selection of seven wolves
from four different packs in winter, Côte-Nord region of Québec (Canada), presented with their selection coefficients (β) and associated 95% confidence interval [CI], the variance (VAR) and covariance (COV) for the random-effects, and for the k-fold cross-validation and associated 95% confidence interval [CI]. Availability was sampled within the 95% MCP of the pack for each individual. ... 98
Table 2.7. Winter movements of wolves in the Côte-Nord region of Québec (Canada)
determined from step selection function of six wolves from three different packs (n = 4301 steps, ranging from 355 to 1308 steps per wolf). Each stratum of the conditional logistic regression included the observed steps, with each one being paired with 10 random steps sharing the same starting point. The model is presented with selection coefficients (β), 95% confidence interval [CI] based on robust empirical standard errors estimated from 139 independent clusters (11-34 clusters per wolf), and from k-fold cross-validation... 101
Table 3.1. Description of the six candidate resource selection functions used for assessing
habitat selection by woodland caribou and wolves in the Côte-Nord region, Québec (Canada). ... 127
Table 3.2. Candidate mixed-effects logistic regression models of habitat selection by
woodland caribou in the Côte-Nord region, Québec (Canada) with their number of parameters (K), maximum log-likelihood (LL), Bayesian Information Criteria (BIC), relative BIC values (Δ BIC), and BIC weight (wi). ... 131
Table 3.3. Mixed-effects logistic regression models of resource selection by 10 female
Table 3.4. Mixed-effects logistic regression models of resource selection by 3-4 wolves in
the Côte-Nord region, Québec (Canada) with their selection coefficients (β), standard error (SE) and for the k-fold cross validation. Availability was assessed within the 95% MCP of a given pack for each wolf. ... 136
Liste des figures
Figure I. Chaîne des conséquences de l‟altération du paysage sur les interactions
comportementales prédateur-proie. ... 12
Figure 1.1. Predator-prey encounters result from both predator and prey habitat selection
strategies displayed in a multi-scale process (i.e., Johnson‟s [1980] order selection). Habitat availability changes between spatial scales (i.e., landscape, home range, home-range-overlap, movement path), or between trophic levels (i.e., predator and prey). Third-order selection reflects the search tactic used by each species and can provide specific outcomes for the predator or the prey. For each species, the changes in habitat availability between the area of home-range-overlap of predator and prey and the movement-path determine the tactic efficiency. The differences in the tactic efficiency and the asymmetry of the tactic outcomes between predator and prey should help to improve our understanding of the predator-prey game. ... 37
Figure 1.2. The habitat features of the wolf-prey encounter sites in the Côte-Nord region of
Québec (Canada) were evaluated using resource selection functions at both home-range-overlap (HRO) and movement-path (MP) levels. The observed wolf-prey path intersections ( ), resulting from both wolf and prey movements, were identical for both species. In contrast, habitat availability was scale dependent and was separately assessed for each species at the MP level. Random sites were drawn within the overlap of 95% MCP for wolves and 100% MCP for prey ( ) at the HRO level, and along the movement paths of wolves ( ) or prey ( ) at the MP level. For each wolf-prey pair, RSFs evaluated where the intersection step non-randomly occurred given the species movement trajectory, the one of wolf (Wolf-Prey) or the one of prey (Prey-Wolf) and revealed the asymmetry of the outcomes of the tactic used. ... 38
Figure 1.3. Minimum convex polygons of wolves (95% MCP) and those of caribou and
moose (100% MCP) used in the wolf-caribou and wolf-moose step encounter analyses during summer-autumn period in the Côte-Nord region of Québec (Canada). Step encounter analyses only considered the home-range overlap (MCP overlap) areas between wolves and their prey. White areas represented the harvested areas (roads, regenerating and recent cuts). ... 45
Figure 2.1. Section of a least-cost network drawn within the graph-theoretic connectivity
framework, and showing potential inter-patch movements represented by links (black line) among network nodes (gray area). Nodes can be connected using several mathematical rules, thereby resulting in variety of graphs: (a) nearest neighbor, (b) minimum spanning tree, and (c) minimum planar graph. The nearest neighbor graph presents several non-connected sub-networks. The minimum spanning tree connects all sub-networks of the neighbor graph with a minimum total length of links. The minimum planar graph contains the maximum number of shortest links such that no two links cross and no other shortest links can be added without creating a cross. ... 73
Figure 2.2. Differences in habitat selection between caribou and moose translated into
caribou was based on nodes composed of conifer stands with lichens together with stands with sparse conifers with mosses, whereas the network of moose was based on nodes composed of mixed and deciduous stands. ... 75
Figure 2.3. Exponential relationship between the percentage of links in a given network
and the percentage of observed inter-patch movements of caribou (n = 23) and moose (n = 10) that occurred between connected nodes for that individual, in that year. Relationship is estimated using a non-linear mixed-effects model with the individual being considered as a random factor. From the smallest to the largest number of links in the network, we find: nearest neighbor (NN), minimum spanning tree (MST), minimum planar graph (MPG), and complete graph (CG). Means with their 95% confidence intervals are presented for each type of network. ... 84
Figure 2.4. Conditional risk of selecting a network node among those available, (a) as a
function of the interaction between Euclidean distance and proportion of cuts and roads within a 1-km radius around network nodes for moose (n = 10), and, (b) for caribou, as a function of the proportion of cuts and roads within a 1-km radius around network nodes and (c) the least-cost path value of the previous inter-patch movement (n = 23) in winter, Côte-Nord region of Québec (Canada). These relationships were estimated from the conditional logistic regression model presented in Table 2.3. ... 90
Figure 2.5. Relative risk of leaving a resource patch at a given time (a) as a function of
least-cost path value of the previous inter-patch movement for caribou (n = 23), and (b) as a function of the interaction between Euclidean distance of the previous inter-patch movement and the proportion of cuts and roads within a 1-km radius around the network nodes for moose (n = 10). These relationships were estimated from the Cox model presented in Table 2.4. ... 93
Figure 2.6. Relative risk of selecting a network node of caribou (cross) or moose (circle)
for wolves (n = 7) during winter in relation to the proportion of cuts and roads around the node, and the degree centrality of the network node, Côte-Nord region of Québec (Canada). These relationships were estimated from the resource selection function model presented in Table 2.6. Large cross and large circle represent the standard node used by wolves within network of caribou and moose, respectively. ... 100
Figure 2.7. Relative probability that a wolf (n = 6) made a step in winter with a given
average speed (step length / step duration) depending upon (a) the proportion of cutovers and roads along the step (Step length × P_CUT/ROAD, Table 2.7), (d) the distance from the nearest cut and road (Step length × DIST_CUT/ROAD, Table 2.7), and (g) the proportion of the step comprised of moose network nodes (Step length x P_NODE_M, Table 2.7). The 9% (a) and 14% (g) values corresponded to the median of the distribution for each covariable. Frequency histograms of turning angles (i.e., the angle between the current and previous direction) of wolf steps are presented with vector length (ρ) and mean turning angle (µ) with 95% confidence intervals. Frequency histograms of turning angles of steps are drawn depending on whether wolf steps were (b) entirely outside cuts or off roads, or (c) entirely inside cuts or on roads. Frequency histograms of turning angles of steps are also presented when the distance
coefficient of circular-linear correlation (r) with its P-value (P) between step length and turning angle of wolves is indicated for each histogram. * indicates that the turning angles have a unimodal distribution according to the Rayleigh test. ... 102
Figure 2.8. Relative probability that a wolf (n = 6) selected a step in winter that was
comprised of a given proportion of caribou network node, as a function of the distance from the nearest cut and road (DIST_CUT/ROAD × P_NODE_C in Table 2.7). Frequency histograms of turning angles of wolf steps are presented with vector length (ρ) and mean turning angle (µ) with 95% confidence intervals. Frequency histograms of turning angles of steps are drawn depending on whether wolf steps were less than (b, c) or at least 80% (d, e) inside of caribou network nodes. Frequency histograms of turning angles of steps are also presented when the nearest cut and road is [0-0.5] km (b, d) or ≥ 10 km (c, e). The coefficient of circular-linear correlation (r) with its P-value (P) between step length and turning angle of wolves is indicated for each histogram. * indicates that the turning angles have a unimodal distribution according to the Rayleigh test. ... 106
Figure 3.1. Study area (18900 km²) located in the Côte-Nord region of Québec, Canada.
The map shows study area boundaries (100% MCP of all caribou locations), wolf territories (95% MCP of all locations of a pack), caribou capture sites, as well as protected forest blocks and the forest harvesting area that were part of the caribou habitat management plan. ... 121
Figure 3.2. Example of net displacement (Euclidean distance between the first location and
the current location) by a female caribou from her capture site. Plateaus between circled zones indicate a low displacement, while sudden transitions indicate an important displacement (five circles). These transitions reflect caribou movements to specific sites such as calving or feeding sites, and the transitions were used to delineate six annual periods. In this example, there was no evident transition between summer and rut, and we used the mean date of all other individuals to set the transition date. ... 123
Figure 3.3. Proportions of the 10 land cover types comprising each of the three landscape
types (i.e., Continuous forest, Protected forest blocks and Forest harvesting area) of the study area. Continuous forest and Protected forest blocks are comprised of seven natural land cover types, whereas Forest harvesting area is also comprised of three anthropogenic land cover types (i.e., regenerating cuts, recent cuts and roads). ... 125
Figure 3.4. Relative probabilities of wolf-caribou co-occurrence in the Côte-Nord region of
Québec, Canada, estimated from Equation 3.3: ̂ ̂ ̂ , where ̂ and ̂ are the relative probability of occurrence estimated for wolf and caribou, respectively. The estimated probabilities of wolf-caribou co-occurrence ( ̂) were scaled between 0 and 1 following: ̂ ( ( )-
- ), where ( ) was the relative probability of co-occurrence at location x and and were, respectively, the smallest and largest values of all probabilities of co-occurrence for the study area. ... 138
À Carole, mon Amour. Tu fus mon étoile salvatrice dans la tempête doctorat. Sans toi j’aurais sombré. Merci.
À Chantal et Alain, mes parents bienveillants. Mon doctorat est le fruit de vos sacrifices.
À Jenny, ma sœur, et Momo, Liam et Tom, sa petite famille. Votre foyer martiniquais fut un havre de paix et une source de bonheur intarissable.
À Paul, un sage et un second père. Le vide laissé est immense. Tu me manques.
À Pierre-Christian Beaubrun. Tu fus bien plus qu’un directeur de recherche pour moi. Ta générosité, ta grandeur d’âme et ton intégrité sont désormais mes garde-fous.
« Admirons les grands maîtres, ne les imitons pas ».
Remerciements
Un doctorat est une longue et exaltante aventure parsemée de moments de doute et d‟épuisement devant la tâche à accomplir. Son aboutissement fut conditionné par la qualité de mon entourage. Mes premières pensées vont tout naturellement vers toi, Carole, qui a partagé le meilleur comme le pire de cette période de notre vie. Je mets enfin un terme à mon étude après sept années de travail. Merci pour ta patience, tes encouragements et ton amour indéfectible. Je ne l‟oublierai jamais. Je remercie également ma famille et mes proches pour leur soutien de tous les instants. Les naissances de Liam et Tom, mes neveux, pendant mes études doctorales, furent une source de motivation supplémentaire et seront associées à jamais à mon aventure canadienne.
J‟exprime toute ma reconnaissance à Daniel Fortin, mon directeur de recherche. En traversant l‟Atlantique, je recherchais un maître à penser. En Daniel, j‟ai trouvé un homme d‟une incroyable vivacité intellectuelle que j‟espère pouvoir atteindre un jour. Ses profondes connaissances théoriques en écologie, son savoir en statistique, sa grande disponibilité (y compris la nuit), son écoute et la rapidité de ses corrections sont autant de qualités qui m‟ont permis de mener à bien mon projet. Merci Daniel, ton exigence m‟a permis de repousser mes limites, même si cela fut parfois douloureux. J‟ai eu la chance d‟avoir un autre pilier pour m‟épauler lors de mes études, Christian Dussault, mon co-directeur de recherche. En Christian, j‟ai puisé ses connaissances du terrain et sa vision de l‟écologie appliquée et théorique me servant de piqûre de rappel lorsque mes recherches s‟en éloignaient. J‟ai apprécié nos conversations, son écoute et ses nombreux encouragements. Merci Christian. Daniel et Christian, vos points de vue différents mais complémentaires m‟ont permis de développer un esprit scientifique qui n‟en est que plus riche. Encore merci à vous deux.
Je tiens à remercier Réhaume Courtois pour son implication dans mon projet, sa grande connaissance du caribou, sa relecture de mes manuscrits et ses encouragements. Merci aussi aux membres de mon comité d‟encadrement de doctorat, Jean-Claude Ruel et Jean-Pierre Tremblay. Vos questionnements et vos conseils furent des plus appréciés et m‟ont permis d‟enrichir mes manuscrits. Je garde un excellent souvenir de nos rencontres
annuelles. Merci aux membres de mon comité d‟examen doctoral, Steeve Côté et David Pothier, qui m‟ont pointé mes lacunes dès le début du doctorat, me permettant ainsi de les combler rapidement et d‟accroître mes capacités de recherche. J‟aimerais également remercier Jean-Pierre Tremblay, David Pothier et Hervé Fritz qui ont accepté d‟arbitrer ma thèse.
Merci aux hommes de terrain sans qui le projet n‟aurait tout simplement pas pu exister : Bruno Baillargeon, Laurier Breton, Serge Couturier, Denis Guay, Dominic Grenier, Jean-Yves Lacasse et Bruno Rochette. Je me souviens des heures passées en votre compagnie au motel l‟Energie à attendre que les conditions météorologiques soient favorables pour sortir en hélicoptère, ou dans l‟hélicoptère à écouter les signaux VHF des animaux, ou en pleine forêt boréale sur la piste d‟un collier GPS à récupérer.
Une autre dualité que celle de mes professeurs a marqué le cours de mon doctorat, celle de deux étudiants en maîtrise que j‟ai accompagnés du début à la fin de leurs études et avec qui j‟ai tissé des liens étroits. Je tiens à remercier Mélina Houle pour sa gentillesse, sa joie de vivre, son énergie folle, pour m‟avoir permis de m‟intégrer à cette culture québécoise à grands pas, et pour m‟avoir initié au squash. Je n‟oublierai pas notre séjour passé en forêt boréale au mois d‟août 2006, lors duquel j‟ai rencontré mes premières mouches noires québecoises, pour le plus grand plaisir de Mélina. Son expérience du terrain me fut précieuse. Nos discussions sur les loups et nos premiers pas côte à côte dans le monde de la sélection d‟habitat et de la géomatique m‟ont permis de progresser très rapidement. Mélina, moi aussi, je suis très heureux d‟avoir eu la chance de travailler avec toi et de te connaître, milles mercis d‟être simplement toi. Guillaume Bastille-Rousseau (ô mon Géant) est une autre belle rencontre. Je le remercie pour sa générosité, sa gentillesse, sa simplicité et sa grande maturité. Je n‟oublierai pas nos parties de badminton, de beach volley et de squash, mon immersion dans le monde du gin qui perdure encore, mon initiation au tir aux pigeons d‟argile et nos visites de Pohénégamook et de Boston. Notre road trip en Californie restera gravé à jamais dans mon âme. Merci aussi pour son esprit critique très aiguisé. Guillaume, je pense sincèrement que tu m‟as aidé autant que je t‟ai aidé (mais pas plus ). Mes efforts à t‟endurer et à t‟aider, mon padawan, auront
Je suis très reconnaissant à tous les autres étudiants et post-docs qui ont traversé le laboratoire durant ces années : Ermias Azeria, Jean-Sébastien Babin, Mathieu Basille, Sabrina Courant, Karine Dancose, Pierre Etcheverry, Viviane Fargeot, Marie-Ève Fortin, Jacqueline Frair, James Hodson, Philippe Jansenn, Cheryl Johnson, Marie-Claude Labbé, Guillaume Latombe, Mélanie-Louise LeBlanc, Jérôme Lemaître, Aurore Malapert, Guillaume Moreau, Maïc Moulinot, David Pinaud, Kim Poitras, Carrie Roever, Marie Sigaud. Merci aussi à ceux que j‟ai découvert par courriel : Orphé Bichet, Chrystel Losier et Olivia Tardy. Nos conversations à caractère scientifique ou non, nos retrouvailles autour d‟une bonne bière de microbrasserie québécoise ou autour d‟un bon repas sont autant de bons souvenirs. Un merci spécial à Jérôme, James et Kim qui furent mes « frères d‟études » dès le début de l‟aventure. Merci à Viviane pour sa joie de vivre et son travail assidu qui a facilité la réalisation de mon premier chapitre. Pierre, Ermias, Philippe, Guillaume L., David, Kim, Jean-Gabriel Elie et Christine Casabon, j‟ai apprécié votre aide lors de mes relevés nivométriques sur la Côte-Nord, qui se firent souvent en motoneige. Merci aussi à James, Jérôme, Ermias, David, Cheryl et Mathieu pour m‟avoir aidé à améliorer mes manuscrits et pour vos conseils statistiques. Merci à Pierre qui a été là dès mon arrivée à Québec et a joué le rôle du « papa » protecteur, je ne l‟oublierai pas et je te suis redevable. Sans toi je ne serais peut être pas en train d‟inscrire ces dernières lignes. Enfin merci à Mathieu et Sabrina d‟avoir gardé mes affaires pendant ces trois dernières années (initialement 6 mois…) où j‟étais absent. James, Kim et LucBen, je me souviens de notre surf trip en Nouvelle-Ecosse ; Jean-Sébastien, LucBen et Guillaume, je me souviens de notre surf trip dans le Maine ; James et Guillaume, je me souviens de nos parties de squash et de badminton; Mathieu, mon bras se souviendra à jamais de nos parties de tennis acharnées. Merci aussi à tous les joueurs de hockey cosom, James, Mathieu, Guillaume…
Comment ne pas remercier aussi mes amis que j‟ai laissés en France et qui m‟ont soutenu pendant ce long périple et dans la dernière ligne droite. Merci donc à Brice Blanloeil, Hugues Perrier, Ashley Kibler, Julien Demotes-Mainard, Charlène Arnaud, Nicolas Téoulé, Antoine Lassort, David Dumon et Jean-Baptiste Dussault.
Je souhaiterais remercier Thierry Duchesne, Marc Mazerolle et Gaétan Daigle pour leurs conseils en statistiques et modélisation, Pierre Racine pour son aide en géomatique et
pour nos heures passées ensemble sur PostGIS, et enfin William Parsons pour la révision de l‟anglais de mes manuscrits.
Merci aussi aux partenaires financiers de cette étude : le Conseil de Recherches en Sciences Naturelles et en Génie (CRSNG)-Chaire en Sylviculture et Faune, le Ministère des Ressources naturelles et de la Faune du Québec, la Fondation Canadienne pour l‟Innovation (FCI), la Fondation de la Faune du Québec, la Fondation Anne Vallée et l‟Université Laval.
Avant-propos
Ce doctorat est présenté sous la forme d‟une thèse avec trois articles scientifiques. La thèse inclut une introduction générale et une conclusion générale qui lient l‟ensemble des articles, bien que chacun puisse être lu indépendamment.
En tant qu‟auteur principal des trois articles, j‟ai élaboré les objectifs de recherche, j‟ai planifié et réalisé en partie l‟échantillonnage sur le terrain, j‟ai effectué les analyses géospatiales et statistiques, et j‟ai rédigé les manuscrits. Mon directeur de recherche, Daniel Fortin, a largement contribué aux étapes d‟élaboration des objectifs et des protocoles d‟échantillonnage et de rédaction des manuscrits. Mon co-directeur de recherche, Christian Dussault, a également fortement participé à la rédaction des manuscrits. Je dois également souligner la contribution de Réhaume Courtois, qui a relu de façon assidue chacun de mes articles dont il est co-auteur.
Je tiens à souligner le travail de Vivianne Fargeot, une étudiante française de l‟Université de Rennes, que j‟ai codirigé avec Daniel Fortin au cours de son stage de master 1. Vivianne a beaucoup contribué à la préparation des données GPS en vue de leur traitement statistique, ainsi qu‟aux analyses statistiques. Le fruit de notre collaboration a aboutit à mon premier chapitre de doctorat pour lequel je suis l‟auteur principal. Le chapitre 1 a été publié dans la revue Journal of Animal Ecology en 2013, avec Daniel Fortin, Christian Dussault, Viviane Fargeot et Réhaume Courtois comme co-auteurs.
Le chapitre 2, pour lequel je suis l‟auteur principal, est actuellement sous presse dans la revue Ecological Monographs, avec Daniel Fortin, Christian Dussault et Réhaume Courtois comme co-auteurs.
Le chapitre 3, dont je suis le principal auteur, a été publié dans la revue Landscape
Ecology en 2009, avec Daniel Fortin, Christian Dussault et Réhaume Courtois comme
co-auteurs.
J‟ai également contribué à quatre articles hors thèse. J‟ai participé à la réalisation des analyses géospatiales de l‟article de Fortin et al. (2008a) publié dans Journal of Applied
Ecology, en tant que contractant. Dans l‟article de Duchesne et al. (2010) publié dans Journal of Animal Ecology, pour lequel je suis troisième auteur, j‟ai réalisé les simulations
de déplacements des individus, une partie des analyses statistiques, et j‟ai révisé le manuscrit. Dans un article de Fortin et al. (2013) publié dans The American Naturalist, je suis quatrième auteur et j‟ai réalisé une partie des analyses statistiques et géospatiales, et j‟ai participé à la révision du manuscrit. Je suis aussi quatrième auteur dans l‟article de Basille et al. (2011) publié dans Le Naturaliste Canadien pour lequel j‟ai révisé le manuscrit.
Finalement, j‟ai participé à la réalisation de deux chapitres de livre. Dans Fortin et
al. (2010) parut dans European bison conservation in Białowieża Forest. Threats and prospects of the population development, pour lequel je suis troisième auteur, j‟ai participé
aux analyses géospatiales et à la réalisation des figures, et j‟ai révisé le manuscrit. Je suis quatrième auteur du chapitre de Fortin et al. (2011) parut dans Woodlands: Ecology,
Management and Conservation, pour lequel j‟ai participé à la préparation des données
Introduction Générale
« … considering the infinite complexity of the relations of all organic beings to each other and to their conditions of existence, causing an infinite diversity in structure, constitution, and habits, to be advantageous to them… ».
Darwin, 1859.
1. Les interactions prédateur-proie
Les interactions entre les espèces représentent un thème d‟étude majeur en écologie depuis plus d‟un siècle et demi (Darwin 1859), notamment pour leur rôle structurant dans les communautés animales (Elton 1927, May 1973, Pimm et al. 1991, Polis et Strong 1996, Stouffer et Bascompte 2010) et leur impact sur les propriétés et les fonctions de l‟écosystème (Schmitz et al. 2008). Les interactions prédateur-proie adviennent entre les différents niveaux trophiques, lorsqu‟une espèce d‟un niveau trophique supérieur se nourrit d‟une espèce d‟un niveau trophique inférieur (Taylor 1984). Les relations prédateur-proie représentent l‟épine dorsale du fonctionnement des communautés animales (Elton 1927, Lindeman 1942) de par leurs impacts sur la dynamique des populations de proies (Lotka 1925, Volterra 1926, Hassell 1978, Turchin 2003, Barbosa et Castellanos 2005, Orrock et
al. 2008, Peckarsky et al. 2008). Par exemple, des études à long terme démontrent que la
densité d‟orignaux (Alces alces) est régulée par la population de loups gris (Canis lupus) dans le parc national de l‟Isle Royale (Michigan, USA) (McLaren et Peterson 1994). Les prédateurs affectent également la diversité spécifique d‟un écosystème (Paine 1966, May 1973, Barbosa et Castellanos 2005). Dans un système théorique comprenant un prédateur et deux proies, une pression de prédation moyenne accroit la diversité en permettant la coexistence des deux proies (Hastings 1978).
1.1. Complexité trophique et interactions prédateur-proie
Les chaînes trophiques linéaires qui ne considèrent que les relations prédateur-proie (Lindeman 1942) ne permettent pas d‟expliquer adéquatement la dynamique des relations
entre les espèces d‟un écosystème plus complexe (Polis et Strong 1996). En effet, les individus des espèces interagissent généralement entre eux au sein de véritables réseaux trophiques caractérisés, en plus des interactions prédateur-proie, par des interactions ayant cours au sein d‟un même niveau trophique comme la compétition (Pimm et al. 1991, Polis et Strong 1996, Linnell et Strand 2000, Barbosa et Castellanos 2005, Stouffer et Bascompte 2010) ou entre plusieurs niveaux trophiques lorsqu‟il y a prédation intraguilde et interguilde (Polis et Holt 1992, Barbosa et Castellanos 2005). Les interactions de compétition modulent les relations prédateur-proie (Werner et Peacor 2003). Par exemple, la présence des libellules (Anax junius) prédatrices modifie l‟activité et l‟utilisation de l‟espace de ses deux proies (Rana catesbeiana et Rana clamitans), leurs interactions de compétition (Werner 1991) et réduit le taux de prédation des autres espèces de libellules sur ces deux proies (Peacor et Werner 1997). Le lien entre prédation et compétition est un élément conceptuel central en écologie des communautés car sa considération permet de mieux expliquer les mécanismes sous-jacents à la dynamique des populations prédateur-proie (Werner et Peacor 2003).
1.2. Compétition apparente et interactions prédateur-proie
Un changement dans la dynamique des populations composant les systèmes prédateur-multi-proies peut survenir à cause d‟un type particulier d‟interaction indirecte entre les deux populations de proies, soit la compétition apparente (Holt 1977). La compétition apparente représente une situation selon laquelle l‟apparition ou l‟augmentation de la densité d‟une des population de proies à la suite de changements environnementaux dans un système prédateur-proies va induire une réponse numérique positive du prédateur, et indirectement créer une augmentation de la pression de prédation sur l‟autre population de proies, qui peut alors subir l‟extinction (Holt 1984, Abrams 1987, Abrams et al. 1998, Chaneton et Bonsall 2000, Bull et al. 2006, DeCesare et al. 2010). Par exemple, les populations de mouflons de la Sierra Nevada (Ovis canadensis californiana) ont décliné à la suite d‟une augmentation de la prédation par le puma (Puma concolor), dont les populations ont augmenté, conjoitement à celles de cerfs mulets (Odocoileus hemionus) qui ont été favorisées par les modifications anthropiques du paysage (Gibson
C‟est dans cette situation de compétition apparente que s‟organise ma thèse de doctorat. Les interactions entre le loup gris, le caribou des bois (Rangifer tarandus caribou) et l‟orignal subissent des modifications selon l‟hypothèse de la compétition apparente sous l‟impulsion de l‟altération de la forêt boréale par l‟humain (Seip 1992, Rettie et Messier 1998, Wittmer et al. 2005). Au vu de la résonnance entre compétition et prédation dans le système loup-caribou-orignal, considérer ces trois espèces simultanément devient indispensable pour mieux cerner les différents mécanismes comportementaux influençant les interactions loup-proies.
« Behavioural ecology is a meeting point for behaviour, ecology and evolution ».
Krebs et Davies, 1993.
2. Une approche comportementale
2.1. Importance des effets non létaux de la prédation
Les effets létaux de la consommation directe des proies (« consumptive effects ») affectent la densité de leurs populations (« density-mediated indirect interactions ») (Abrams et al. 1996) selon un effet numérique (Creel et Christianson 2008). Ces effets numériques de consommation ont été largement étudiés et représentent la base de notre compréhension des interactions prédateur-proie (Murdoch et al. 2003). En revanche, les effets non létaux (« nonconsumptive effects ») des prédateurs sur les proies sont beaucoup moins connus (Luttbeg et Kerby 2005, Creel et Christianson 2008). Ils affectent la densité des populations de proies, en induisant indirectement le développement de stratégies comportementales de défense coûteuses de la part des proies (« trait-mediated indirect interactions ») (Lima et Dill 1990, Werner et Peacor 2003, Schmitz et al. 2004, Barbosa et Castellanos 2005, Preisser et al. 2005, Prasad et Snyder 2006, Valeix et al. 2009). Pour diminuer leur risque de prédation, les proies adoptent tout un arsenal de réactions morphologiques, physiologiques, développementales et comportementales (Lima et Dill 1990, Lima 1998, Werner et Peacor 2003, Schmitz et al. 2004, Creel et al. 2007, Cresswell
en vigilance chez les impalas (Aepyceros melampus) et les zèbres des plaines (Equus
quagga) (Périquet et al. 2012) et la vie en groupe (Fryxell et al. 2007) chez le junco ardoisé
(Junco hyemalis) (Lima 1995) et le bison (Bison bison) (Fortin et al. 2009).
Ces effets non létaux ont un rôle tout aussi important, voire parfois plus important, que les effets létaux sur les populations prédateur-proie (Abrams 1984, revue dans Werner et Peacor 2003, Schmitz et al. 2004, méta analyse de Preisser et al. 2005, Trussell et al. 2006, Creel et Christianson 2008). Par exemple, les prédateurs peuvent induire une réaction des proies même lorsqu‟il n‟y a aucun risque de mourir de prédation (Creel et Christianson 2008, Sheriff et al. 2010). De plus, l‟importance relative des effets non létaux par rapport aux effets de consommation dépend du type de comportement prudent ou aggressif de la proie (Sih et al. 2012) et du prédateur (Preisser et al. 2007), et donc de leurs différentes stratégies comportementales.
Les effets non létaux affectent également la dynamique des communautés en intervenant dans la plupart des cascades trophiques (revue dans Schmitz et al. 2004). Les cascades trophiques représentent les effets indirects des prédateurs sur les plantes par l‟entremise des proies (Beckerman et al. 1997, Polis et al. 2000). La réintroduction du loup gris dans le parc national de Yellowstone en est un exemple. Avec le retour de leur prédateur, les wapitis (Cervus canadensis) ont modifié leur comportement et leur répartition spatiale (Fortin et al. 2005a), et la situation de surbroutement du peuplier faux-tremble (Populus tremuloides) par les wapitis a diminué, du moins à court terme, dans les zones présentant un fort risque de prédation (Ripple et al. 2001, mais voir Kauffman et al. 2010). Une cascade trophique similaire impliquant le loup, l‟orignal et le sapin baumier (Abies balsamea) a également été mise en évidence sur l‟Isle Royale (McLaren et Peterson 1994). La considération des modifications comportementales des différentes proies induites par les effets non létaux des prédateurs devient alors d‟une importance capitale pour mieux définir les relations trophiques dans les systèmes prédateur-multi-proies et, ultimement, pour mieux comprendre le fonctionnement des communautés (Lima 1998, Preisser et al. 2005).
2.2. Le comportement animal : un garde-fou des changements populationnels Les processus démographiques sont caractérisés par de nombreux décalages temporels qui font souvent des changements de la taille des populations un indicateur final des changements environnementaux (Morris 2003). Par contre, les réactions comportementales des animaux (p. ex., la sélection d‟habitat, les décisions de déplacements) apparaissent dans un très court intervalle de temps et sont souvent précurseurs des changements populationnels (Remeš 2000, Fryxell et al. 2005, Schick et al. 2008, Morales et al. 2010, Sih et al. 2012) et du fonctionnement de l‟écosystème (Morris 2003). Ainsi, l‟étude des réactions comportementales des animaux est un bon moyen d‟anticiper les conséquences des modifications anthropiques du paysage sur les tailles de populations (Festa-Bianchet et Apollonio 2003, Sih et al. 2012). Ma thèse s‟interessera donc à déceler les modifications de comportement du loup et de ses deux proies principales, le caribou et l‟orignal, à la suite de perturbations anthropiques, qui pourraient être révélatrices des changements s‟opérant dans leurs interactions trophiques en forêt boréale aménagée.
« Differential habitat selection is one of the principal relationships which permit species to coexist ».
Rosenzweig, 1981.
3. La répartition spatiale : une résultante comportementale
La répartition spatiale des animaux peut avoir une grande influence sur la dynamique de leurs populations (Tilman et Kareiva 1997, Turchin 1998, Morris 2006) et sur les interactions entre les espèces (McLaughlin et Roughgarden 1992, Lima et Zollner 1996, Tilman et Kareiva 1997, Turchin 1998, Turner et al. 2001). Une des principales conséquences des effets non létaux de la prédation est la modification de la répartition spatiale des proies. Par exemple, pour diminuer leur risque de prédation, les proies peuvent migrer (Bergerud et al. 1990, Hebblewhite et Merrill 2007), se disperser (Bergerud et al. 1990), altérer leur sélection d‟habitat (Gilliam et Fraser 1987, Lima et Dill 1990, Lima 1998, Brown et al. 1999, Mao et al. 2005, Dupuch et al. 2009), sélectionner un type de
milieu spécifique (Creel et al. 2005, Valeix et al. 2009), ajuster leurs décisions de déplacement (Mitchell et Lima 2002, Fortin et al. 2005a, Hodson et al. 2010a) et changer leur utilisation de l‟habitat de manière temporelle (Gude et al. 2006, Valeix et al. 2009). La répartition spatiale des animaux est largement dictée par deux processus comportementaux : les déplacements et la sélection d‟habitat (Figure I).
3.1. Le déplacement animal
Le déplacement est le processus par lequel les individus se répartissent dans l‟espace au cours du temps (Turchin 1998). Selon le principe de sélection naturelle, les animaux se déplacent dans leur environnement afin de survivre, de se développer et de se reproduire (Fahrig 2007, Nathan et al. 2008). Les déplacements des animaux reflètent donc en partie la nature des compromis coût/bénéfice entre le risque de prédation et l‟acquisition de nourriture (Lima et Dill 1990, Mitchell et Lima 2002, Brown et Kotler 2004, Fortin et
al. 2005a, Morales et al. 2005, Chetkiewicz et al. 2006, Fahrig 2007, Hodson et al. 2010b).
Les patrons de déplacements sont aussi influencés par la structure et la composition du paysage (Fryxell et al. 2008, Ovaskainen et al. 2008), comme chez le wapiti (Fortin et al. 2005a), le bison (Dancose et al. 2011), le caribou (Johnson et al. 2002) et le loup (Dyer et
al. 2002, Whittington et al. 2004, Bergman et al. 2006). Notre compréhension empirique du
déplacement animal dans le paysage est toutefois encore limitée (Fahrig 2007, mais voir Giuggioli et Bartumeus 2010, McClintock et al. 2012), de même que son impact sur les populations (Turchin 1998, Chetkiewicz et al. 2006). En effet, il existe relativement peu d‟études expliquant les mécanismes comportementaux sous-jacents aux décisions de déplacement (mais voir Fortin et al. 2005a, Morales et al. 2005, Fryxell et al. 2008, Dancose et al. 2011) et incluant plus d‟une espèce (Holyoak et al. 2008), possiblement à cause de la difficulté de quantifier les déplacements (Chetkiewicz et al. 2006). Ceci réduit largement notre capacité à comprendre les stratégies de déplacement de chaque espèce d‟un point de vue évolutif (Holyoak et al. 2008, Nathan et al. 2008).
3.2. La sélection d’habitat
prédation, partenaire sexuel, abri contre le stress abiotique) et des conditions écologiques (compétition, prédation, conditions abiotiques) qui lui permettent d‟occuper l‟espace (Hall
et al. 1997, Mitchell 2005). La sélection d‟habitat est un processus spatio-temporellement
hiérarchique (Johnson 1980, Senft et al. 1987, Arthur et al. 1996, Bailey et al. 1996, Boyce 2006, Gustine et al. 2006a, Courbin et al. 2009, Hebblewhite et Merrill 2009, Valeix et al. 2009, DeCesare et al. 2012) par lequel un animal utilise certaines composantes de l‟habitat disproportionnellement à leurs disponibilités (Johnson 1980). Ce processus reflète la manière dont l‟animal cherche à améliorer son aptitude phénotypique (McLoughlin et al. 2006, Morris 2006, Aldridge et Boyce 2007, McLoughlin et al. 2007, Fortin et al. 2008b, Dussault et al. 2012) en réalisant un compromis entre le risque de prédation et l‟acquisition de ressources (Fryxell et Lundberg 1997, Lima 2002, Gaillard et al. 2010). L‟importance relative du risque de prédation ou de la disponibilité de nourriture à une échelle donnée dépend des différences d‟hétérogénéité spatiale de chaque facteur (Gaillard et al. 2010). Par exemple, si la répartition de la nourriture est plus hétérogène à large qu‟à fine échelle spatiale et vice versa pour le risque de prédation, alors la sélection d‟habitat devrait être guidée en priorité par l‟acquisition de nourriture à large échelle et par l‟évitement des aires à fort risque de prédation à fine échelle (Gaillard et al. 2010). Dans les environnements présentant une très forte variation saisonnière (écosystèmes de montagnes, arctique), les proies devraient éviter les facteurs limitants à large échelle (Rettie et Messier 2000). La sélection pour un attribut de l‟habitat peut aussi varier en fonction de son abondance dans le paysage ou en fonction de l‟abondance d‟autres caractéristiqures du paysage (Fortin et al. 2008a, Hebblewhite et Merrill 2008, Godvik et al. 2009, Gillies et Cassady St. Clair 2010, Houle et al. 2010, Moreau et al. 2012); c‟est ce que l‟on appelle la réponse fonctionnelle dans la sélection d‟habitat (Mysterud et Ims 1998, Matthiopoulos et al. 2011).
Chez les ongulés, le compromis entre le risque de prédation et l‟acquisition de nourriture peut être observé durant les déplacements (Fortin et al. 2005b), à l‟intérieur du domaine vital saisonnier ou annuel (Bailey et al. 1996, Rettie et Messier 2000, Dussault 2002, Dussault et al. 2005a, DeCesare et al. 2012) et à l‟échelle du paysage (Dussault et al. 2005a, DeCesare et al. 2012). Certains facteurs abiotiques, comme la topographie, peuvent influencer la sélection d‟habitat des animaux à une échelle étendue (Bailey et al. 1996) comme chez le wapiti (Boyce et al. 2003), ou à une échelle plus fine chez le wapiti (Fortin
et al. 2005b) et le loup (Hebblewhite et al. 2005a). La sélection d‟habitat s‟organise selon des gradients géographiques chez le caribou (Fortin et al. 2008a) et varie également selon la saison chez le wapiti (Boyce et al. 2003), le caribou (Courbin et al. 2009), le loup (Houle
et al. 2010) et l‟orignal (Basille et al. 2012). Certains facteurs saisonniers sont susceptibles
d‟influencer la sélection d‟habitat à fine échelle, comme la neige (Dussault 2002, Fortin 2003, Fortin et al. 2005b, Dussault et al. 2005a, Mayor et al. 2007, Courbin et al. 2009), le harassement des insectes (Timmermann et McNicol 1988) et le stress thermique (Dussault
et al. 2004). Par exemple, la neige va restreindre les déplacements des orignaux lorsque sa
profondeur dépasse 60 cm, car les coûts énergétiques de locomotion augmentent exponentiellement au delà de cette limite (Renecker et Schwartz 1998).
3.3. Évaluer la sélection d’habitat et les déplacements
La sélection d‟habitat peut être évaluée en comparant le taux d‟utilisation de la ressource (c.-à-d., la quantité de ressource utilisée par un individu ou une population dans une période de temps fixe) avec sa disponibilité (c.-à-d., la quantité de ressource accessible à l‟individu ou à la population au cours de cette même période de temps). Il existe un vaste panel de méthodes statistiques relativement simples pour y parvenir (McClean et al. 1998), comme l‟analyse compositionelle (Aebischer et al. 1993). Cependant, la fonction de sélection des ressources (FSR, Manly et al. 2002) est actuellement l‟un des outils les plus utilisés et appropriés pour quantifier la sélection d‟habitat des animaux (McLoughlin et al. 2010). Une FSR lie la répartition des animaux aux patrons d‟hététogénéité spatiale de l‟habitat en comparant les composantes de l‟habitat caractérisant les sites fréquentés par l‟animal (p. ex., localisations GPS [Global Positioning System]) avec les composantes de l‟habitat de sites tirés aléatoirement (Boyce et McDonald 1999, Boyce et al. 2002, Manly et
al. 2002, Johnson et al. 2006, DeCesare et al. 2012). Les FSRs sont généralement
modélisées par une régression logistique (Gillies et al. 2006, Hebblewhite et Merrill 2008, Koper et Manseau 2009, Dussault et al. 2012) ou par une régression logistique conditionnelle (Whittington et al. 2005, Aarts et al. 2008, Craiu et al. 2008, Fortin et al. 2009, Koper et Manseau 2009, Duchesne et al. 2010). La définition de la disponibilité est un élément clé à considérer dans les études de sélection d‟habitat car elle influe directement
définition unique de la disponibilité car cela dépend avant tout de l‟objectif auquel on s‟intéresse (Boyce et al. 2003, Boyce 2006). La disponibilité peut être évaluée à de larges échelles spatiales, comme le paysage, le domaine vital et le domaine vital saisonnier, ou à des échelles plus fines, comme la parcelle d‟alimentation ou dans une zone tampon autour des localisations (revue dans Boyce 2006).
Les récents développements technologiques en matière de suivis GPS des animaux (c.-à-d., augmentation de la fréquence d‟échantillonnage) permettent généralement de localiser les individus plus fréquemment qu‟avec la télémétrie VHF (Very High Frequency), et donc de mieux caractériser leurs déplacements. Ceci facilite l‟utilisation d‟une approche Lagrangienne (c.-à-d., caractérisation des composantes du déplacement : magnitude, vitesse, directionnalité) (Turchin 1998, Chetkiewicz et al. 2006) qui permet de mieux comprendre les décisions de déplacement des animaux (Cagnacci et al. 2010). Dans cette optique, la fonction de sélection de pas (FSS, Fortin et al. 2005a, technique similaire aux FSRs) a été proposée pour caractériser les déplacements des animaux. Dans une FSS, on compare les caractéristiques de l‟habitat le long des pas de l‟animal (c.-à-d., ligne droite entre deux localisations consécutives, Turchin 1998) avec celles le long de pas tirés aléatoirement (Fortin et al. 2005a, Coulon et al. 2008, Forester et al. 2009, Roever et al. 2010).
3.4. Influence des activités humaines
En modifiant l‟habitat, les perturbations humaines peuvent changer la sélection de l‟habitat (Houle et al. 2010, Moreau et al. 2012) et les déplacements des animaux (Trombulak et Frissell 2000, Fahrig 2007, Roever et al. 2010). Les modifications de la répartition spatiale des espèces à la suite d‟activités humaines (Johnson et al. 2005) peuvent ainsi avoir des conséquences sur les interactions trophiques (Debinski et Holt 2000, Courbin et al. 2009, Figure I) et sur les populations animales (Trombulak et Frissell 2000, Aldridge et Boyce 2007, Fahrig 2007, Wasser et al. 2011, Dussault et al. 2012). Par exemple, dans la forêt boréale de l‟est du Canada, la présence des routes et des forêts décidues au sud dues en partie à la sylviculture favorisent les communautés d‟orignaux, de cerfs de Virginie (Odocoileus virginianus) et de loups, alors qu‟au nord la forêt de conifères matures non perturbée par l‟homme favorise les communautés de caribous et de
carcajous (Gulo gulo) (Bowman et al. 2010). Les mécanismes comportementaux affectant la réaction des animaux aux perturbations anthropiques de l‟habitat (Debinski et Holt 2000) restent peu étudiés à ce jour (Ryall et Fahrig 2006). Il existe notamment un manque de connaissances important sur les effets qu‟ont ces perturbations sur les interactions prédateur-proie, particulièrement sur le jeu prédateur-proie.
« All of life is a game and evolution by natural selection is no exception ».
Vincent et Brown, 2005.
4. Le jeu comportemental prédateur-proie
Certains aspects des tactiques de déplacement et de sélection d‟habitat des animaux peuvent être expliqués par la théorie des jeux. La théorie des jeux (Von Neumann et Morgenstern 1944) prédit que le choix fait par chacun des joueurs devrait devenir stable (p. ex., l‟équilibre de Nash) car aucun d‟entre eux ne peut modifier sa stratégie sans affaiblir sa position personnelle (Nash 1951). Le jeu prédateur-proie est une conséquence évolutionniste de la théorie des jeux (Maynard Smith 1982, Vincent et Brown 2005). Des travaux théoriques (Sih 1998, Sih 2005) se sont intéressés aux issues du jeu prédateur-proie en utilisant des modèles à trois niveaux trophiques (c.-à-d. : carnivore, herbivore et plante qui intéragissent) et en évaluant la répartition spatiale des espèces entre des parcelles de différentes qualités attendue sous un régime de distribution idéale libre (Fretwell et Lucas 1969). Les modèles trouvent des stratégies évolutivement stables (Maynard Smith 1979, Maynard Smith 1982) où ni le prédateur ni la proie ne pourraient améliorer leur aptitude phénotypique en changeant de parcelle (analogie évolutionniste de l‟équilibre de Nash). Le jeu prédateur-proie suit donc une dynamique darwinnienne où chaque stratégie est associée à une aptitude phénotypique particulière (Vincent et Brown 2005).
4.1. Le jeu spatial prédateur-proie
dynamique (Brown et al. 1999, Lima 2002, Sih 2005) avec chaque espèce qui s‟adapte aux comportements de l‟autre (Flaxman et al. 2011) et ultimement à la répartition spatiale de l‟autre (Brown et al. 1999, Bouskila 2001, Brown et al. 2001, Lima 2002, Sih 2005, Hammond et al. 2007, Dupuch et al. 2009, Flaxman et Lou 2009, Luttbeg et al. 2009, Valeix et al. 2009, Berger-Tal et al. 2010, Laundré 2010, Flaxman et al. 2011, Hammond
et al. 2012, Fraker et Luttbeg 2012, Williams et Flaxman 2012) (Figure I). Dans ce « jeu »
entre le prédateur et sa proie, chaque espèce vise à maximiser son aptitude phénotypique, compte tenu des différentes contraintes auxquelles elle fait face (Sih 1984, Sih 1998). Le prédateur fait un compromis entre la compétition, son taux de rencontre avec ses proies et ses chances de capturer une proie étant donné une rencontre. Au contraire, la proie cherche à acquérir ses différents types de ressource tout en minimisant la compétition et le risque de prédation. Pour rester imprévisibles et ainsi diminuer leur risque de prédation, les proies peuvent se déplacer fréquemment entre leurs parcelles de ressources (Mitchell 2009); c‟est le jeu de « passe-passe » (« shell game », Mitchell et Lima 2002). Dans un environnement où la proie est furtive et peu abondante, les prédateurs pourraient davantage rechercher leurs proies dans des secteurs où les caractéristiques de l‟habitat sont favorables à la proie plutôt que de rechercher les proies directement (« resource tracking », Flaxman et Lou 2009). Par exemple, durant la nuit, le puma passe 70 % de son temps près ou dans les parcelles de forêt riches en ressources pour les cerfs mulets, alors que ces derniers passent 60 % de leur temps dans les milieux ouverts qui sont pauvres en ressources alimentaires, mais présentent un faible risque de prédation (Laundré 2010). Malgré les coûts associés au comportement anti-prédateur (Bouskila 2001, Schmitz et al. 2004, Schmitz 2005), le jeu prédateur-proie peut être caractérisé par des réactions comportementales chroniques de la part des proies. Par exemple, les cerfs de Virginie sont plus abondants aux frontières des territoires de loups qui agissent comme des refuges pour diminuer leur risque de prédation (Mech 1977, Taylor et Pekins 1991, Lewis et Murray 1993, Mech et Boitani 2003). L‟asymétrie des bénéfices, des coûts et des contraintes entre le prédateur et sa proie peut être telle qu‟un effet saute-mouton (« leapfrog effect » Sih 1998) peut résulter du jeu spatial prédateur-proie. Les proies sous-utilisent alors les parcelles dans lesquelles leurs propres ressources abondent afin de diminuer leur risque de prédation (Sih 1998, Hammond et al. 2007, Laundré 2010), tandis que les prédateurs se concentrent dans ces parcelles riches en
Figure I. Chaîne des conséquences de l‟altération du paysage sur les interactions comportementales prédateur-proie.

![Figure 1.1. Predator-prey encounters result from both predator and prey habitat selection strategies displayed in a multi-scale process (i.e., Johnson‟s [1980] order selection)](https://thumb-eu.123doks.com/thumbv2/123doknet/7443386.220804/65.918.181.790.266.656/predator-encounters-predator-selection-strategies-displayed-johnson-selection.webp)






![Table 2.1. Mixed-effects logistic regression models of habitat selection of 24 caribou and 15 moose in winter, in the Côte-Nord region of Québec (Canada) with their selection coefficients (β) and associated 95% confidence intervals [CI], an](https://thumb-eu.123doks.com/thumbv2/123doknet/7443386.220804/111.918.152.781.277.1035/logistic-regression-selection-selection-coefficients-associated-confidence-intervals.webp)
