Reduced white matter integrity in sibling pairs discordant for bipolar disorder.
Texte intégral
Figure
Documents relatifs
Distribution of the mean FA values for the selected white matter tracts within the fronto-cerebellar circuit (mcp = middle cerebellar peduncle, scp = superior cerebellar peduncle)
Diffusion tensor imaging red-green-blue maps in a representative control subject and patients with frontal variant and temporal variant of frontotemporal dementia (fvFTD and
The golden rules for people being treated for bipolar disorder are: never stop taking medicines even if you feel completely recovered, contact your doctor immediately if you
Hou, Cleak and Peveler [12] focused more specifically on the impact of illness representation on adherence in bipolar disorder with the Revised Illness
Factors associated with adherence to medication according to linear regression models using MARS a total score, BIS-10 b nonplanning impulsivity sub-scale score, age, illness
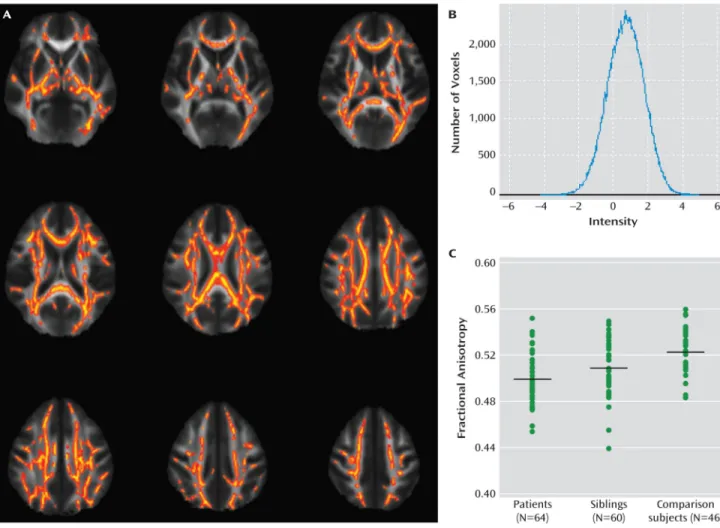
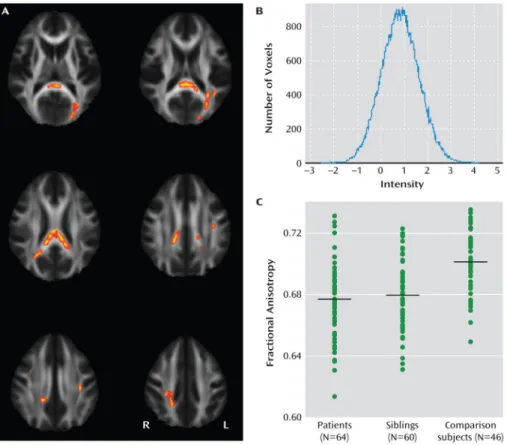
To study white matter abnormalities using whole-brain tractography in a large international multicenter sample of patients with bipolar disorder type 1 and to compare these
Bremer et al observed that polymorphisms in NTRK2 and INPP1 genes were associated with the response to lithium, and also with both suicidal ideation and post-traumatic
