Remotely-sensed changes in the primary productivity of
migratory caribou calving grounds and summer pasture:
the mixed influences of climate change and caribou
herbivory
Mémoire
Allen Campeau
Maîtrise en biologie
Maître ès sciences (M.Sc.)
Québec, Canada
© Allen Campeau, 2016
Remotely-sensed changes in the primary productivity of
migratory caribou calving grounds and summer pasture:
the mixed influences of climate change and caribou
herbivory
Mémoire
Allen Campeau
Sous la direction de :
Steeve D. Côté, directeur de recherche
iii
Résumé
Nous avons utilisé la télédétection pour examiner comment l’abondance du caribou migrateur pouvait influencer la quantité de ressources alimentaires, et comment ces changements pouvaient affecter la dynamique de population et les patrons d’utilisation de l’espace des caribous. Nous avons évalué les relations entre le caribou et ses ressources alimentaires pour l’aire de mise bas et l’aire d’estivage du troupeau Rivière-George (TRG) du nord du Québec et du Labrador (Canada) entre 1991 et 2011. Nous avons modélisé les relations entre la productivité primaire et des variables climatiques, nous permettant d’isoler les effets d’autres facteurs, comme la pression de broutement des caribous. Nous avons trouvé une relation négative entre la densité de caribous et la productivité primaire à grande échelle, suggérant que la pression de broutement par les caribous pouvait réduire l’abondance des ressources alimentaires et contribuer à la dégradation de l’habitat. Une forte tendance au réchauffement durant la période d’étude, couplée avec un déclin de la taille de population du TRG, a cependant entrainé une productivité primaire plus élevée. Cette hausse de la productivité primaire pourrait représenter un rétablissement de la
végétation suite à la réduction de la pression de broutement et/ou un effet du réchauffement climatique.
iv
Abstract
This thesis used remote sensing to examine how changes in caribou abundance can impact forage availability, and how these changes might in turn affect the population dynamics and space use patterns of migratory caribou. We investigated these relationships for the Rivière-George caribou herd (RGH) and its calving grounds and summer range in Northern Quebec and Labrador (Canada) from 1991-2011. We modelled the relationships between primary productivity and various climatic variables, allowing us to isolate the effects of non-climatic factors, such as caribou herbivory, on primary productivity. We found a negative relationship between caribou density and primary productivity at the range-scale, suggesting that caribou herbivory can reduce forage abundance and contribute to habitat degradation. However, a strong warming trend over the study period, coupled with a decline in RGH population size, meant that we observed a trend towards higher primary productivity. This greening trend may represent vegetation recovery following the reduction in caribou herbivory and/or an effect of climate warming.
v
Table of Contents
Résumé ... iii
Abstract ... iv
Table of Contents ... v
List of Tables ... vii
List of Figures ... viii
Acknowledgements ... x
Avant-propos ... xii
1. Introduction ... 1
1.1 Ungulate population dynamics, population size, and range size ... 1
1.1.1 The influence of density dependence ... 1
1.2 Ungulate range use, movement patterns, and habitat selection ... 2
1.2.1 Biotic controls on movement and space use: ecological trade-offs ... 3
1.2.2 Abiotic controls on movement and space use: topography and climate ... 3
1.3 Remote sensing for ungulate range assessment ... 4
1.3.1 The Normalized Difference Vegetation Index and other vegetation indices .. 5
1.4 Study context: migratory caribou on the Quebec-Labrador Peninsula ... 7
1.4.1 Migration patterns and seasonal changes in range use ... 8
1.4.2 Knowledge gaps: understanding interannual changes in range use ... 9
1.4.3 Knowledge gaps: understanding the role of caribou herbivory in habitat change ... 10
1.5 Objectives ... 12
1.5.1 General objectives ... 12
1.5.2 Specific objectives ... 12
2. Remotely-sensed changes in the primary productivity of migratory caribou calving grounds and summer pasture: the mixed influences of climate change and caribou herbivory ... 13
Résumé ... 14
Abstract ... 15
2.1 Introduction ... 16
vi
2.2.1 Study area ... 20
2.2.2 Caribou range delimitation ... 21
2.2.3 Caribou population size and density estimation ... 23
2.2.4 Remotely-sensed data processing ... 24
2.2.5 Assessing the caribou density influence on primary productivity ... 25
2.3.1 The cNDVI-climate models ... 26
2.3.2 Caribou range use and habitat characteristics ... 27
2.3.3 Caribou herbivory effects on primary productivity ... 31
2.4 Discussion ... 35
2.4.1 The model residual approach ... 36
2.4.2 The exploitation ecosystem hypothesis ... 38
2.4.3 Changes in calving ground use ... 40
2.5 Conclusion ... 41
Acknowledgements ... 41
3. Appendix 1: Remotely-sensed changes in lichen abundance on the Rivière-aux-Feuilles caribou herd winter range ... 43
4. General conclusion ... 50
4.1 Caribou herbivory impacts on habitat quality, population dynamics, and range use ... 50
4.2 The use of remote sensing tools to evaluate changes in lichen abundance ... 53
5. References ... 55
vii
List of Tables
Table 3.1. Land cover classes tested with supervised classifications for the barren-ground
winter range of the Rivière-aux-Feuilles caribou herd in Northern Quebec
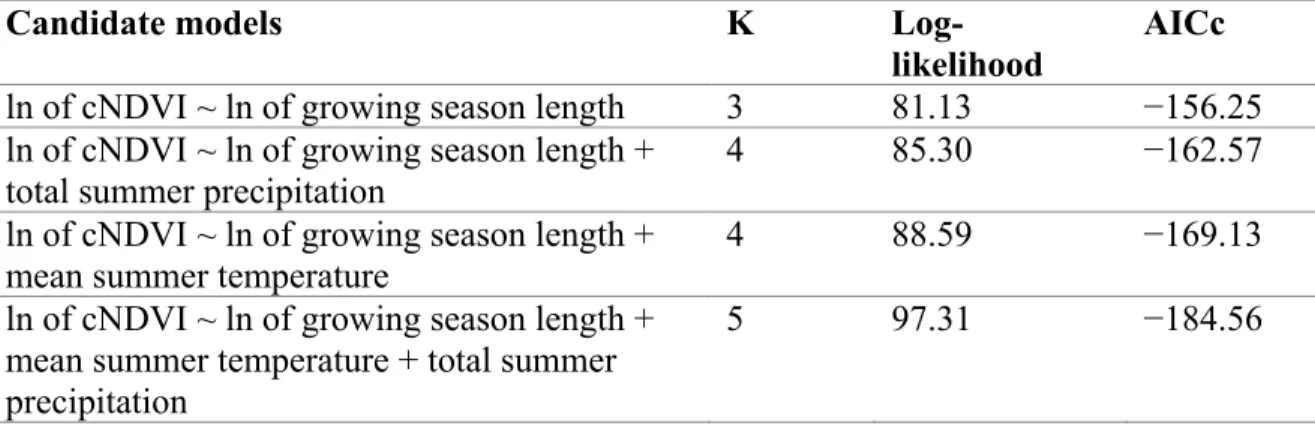
Table S1. Candidate models for cumulative growing season NDVI (cNDVI) in the
Rivière-George caribou herd global 1991-2011 calving grounds, defined by the 100% minimum convex polygon of adult female caribou locations during the calving period; growing season length, total summer precipitation, and mean summer temperature were tested as predictor variables.
Table S2. Candidate models for cumulative growing season NDVI (cNDVI) in the
Rivière-George caribou herd global 1991-2011 summer range, defined by the 100% minimum convex polygon of caribou locations during the summer period; growing season length, total summer precipitation, and mean summer temperature were tested as predictor variables.
Table S3. Trends in primary productivity (cNDVI), cNDVI-climate model residuals, and
climatic variables for the 1991-2011 period in the Rivière-George caribou herd global calving grounds, global summer range, and 1991-2000 and 2001-2011 core calving areas.
Table S4. Linear regression results for the relationships between caribou density (caribou
km-2) and mean cNDVI-climate model residuals for the different seasonal ranges and core
viii
List of Figures
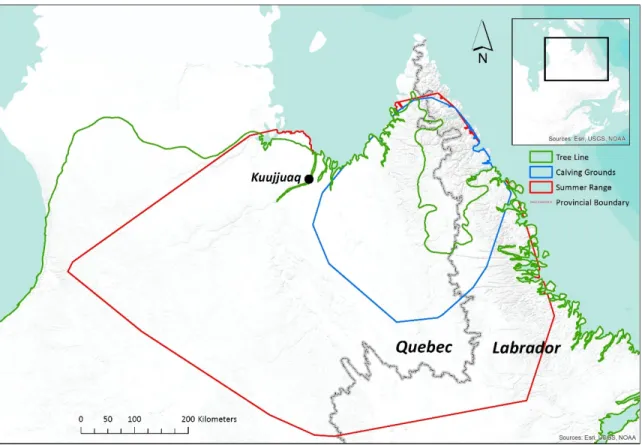
Figure 2.1. The global calving grounds and summer range of the Rivière-George caribou
herd over the 1991-2011 period, as defined by minimum convex polygons (MCPs), clipped to the coastline; the calving grounds MCP is based on the locations of collared females, whereas the summer range MCP is based on the locations of both collared males and females.
Figure 2.2. A loess smoothing spline fitted to aerial survey estimates of Rivière-George
caribou herd population size (black data points) to produce annual population size estimates. Error bars represent confidence intervals (α = 0.10) associated with the aerial survey data. The gray shaded area represents the 1991-2011 study period.
Figure 2.3. The distribution of the Rivière-George caribou herd in Northern Quebec and
Labrador during the calving period for (A) 1991-2000 and (C) 2001-2011, and during the summer period for (B) 1991-2000 and (D) 2001-2011. The gradient from cool to warm colours represents the number of overlapping individual Brownian Bridge (BB) 95% home ranges, with warmer colours representing areas with a greater degree of overlap. Each BB represents the seasonal home range of one collared individual; data from all collared individuals were used to delimit the summer range, but only data from adult females were used to delimit the calving grounds. The total number of BBs varied as follows: (A) 166, (B) 147, (C) 140, (D) 194. Core areas, shown in black crosshatch, represent areas with ≥ 70% of the maximum number of BB overlaps: (A) ≥ 13 BB overlaps, (B) ≥ 43, (C) ≥ 35, (D) ≥ 99. Note that areas with < 3 BB overlaps are excluded from the maps.
Figure 2.4. Interannual changes in cNDVI-climate model residuals, cNDVI, mean summer
temperature, and growing season length in (A) the global 1991-2011 calving grounds, (B) the global 1991-2011 summer range, (C) the 1991-2000 core calving area, and (D) the 2001-2011 core calving area of the Rivière-George caribou herd. All values represent means calculated from all non-water pixels within each area.
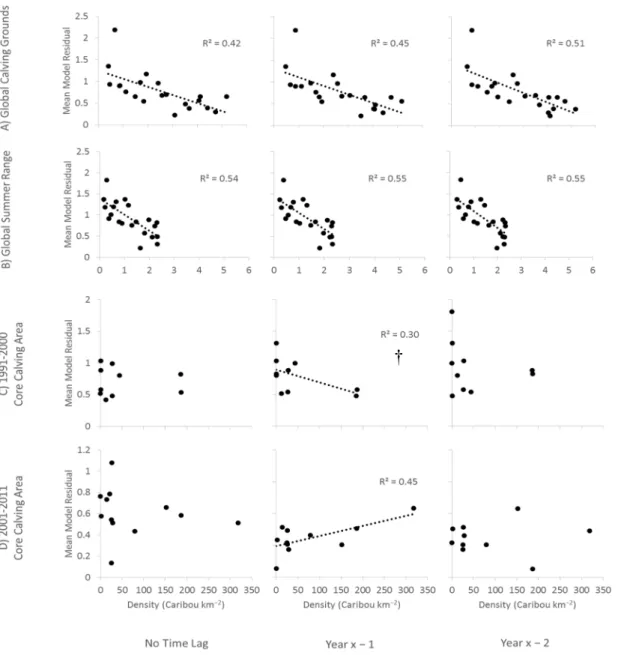
Figure 2.5. The relationships between mean cNDVI-climate model residuals and estimates
of Rivière-George herd caribou density, lagged by 0-2 years with respect to the model residual data, for (A) the global 1991-2011 calving grounds, (B) the global 1991-2011 summer range, (C) the 1991-2000 core calving area, and (D) the 2001-2011 core calving area. Sample size is consistently n = 21 for (A) and (B), with one data point for each year of the 1991-2011 period, and n = 10 for (C), with one data point for each year of caribou presence in the core area. Sample size varies depending on the time lag used for (D), from n = 12 with no time lag to n = 10 for a 2 year lag. The density estimates for the global 1991-2011 calving grounds refer only to adult females, but the other density estimates refer to all individuals. R2 values are provided for all significant relationships (α = 0.05) and
near-significant relationships (marked with †). These relationships were also tested for 3-6 year lags (Table S4 in appendix).
ix
Figure 2.6. Interannual changes in cNDVI-climate model residuals and estimates of
caribou density or Rivière-George herd population size for (A) the global 1991-2011 calving grounds and summer range, and (B) the 1991-2000 and 2001-2011 core calving areas. Caribou density estimates refer only to adult females. The time series for the two core calving areas can be separated into two periods based on the density estimates: caribou present and caribou absent. Model residual and density values represent means calculated from all non-water pixels within each area.
Figure 3.1. Scatterplot of pixel surface reflectance values for Landsat bands 3 and 4 on the
barren-ground portion of the Rivière-aux-Feuilles caribou herd winter range, illustrating the mean values and ellipses for two different lichen class training areas (green = more than 25% lichen cover, red = less than 25% lichen cover). The considerable overlap indicates that there is confusion between the classes in the final image classification.
Figure 3.2. The influence of phenological variation on surface reflectance values and
image classification for the 2012 Landsat composite of Ungava Peninsula in Northern Quebec, Canada (light grey = water or NoData, medium grey = more than 50% lichen cover, black = less than 50% lichen cover). Artificial banding is apparent in the subset image.
x
Acknowledgements
Firstly, I would like to thank Steeve Côté, my MSc thesis supervisor, for giving me the opportunity to take on this project. I joined the Caribou Ungava lab at a time of transition, with Phase 1 winding down and Phase 2 just beginning, so I consider myself fortunate to have been able to sneak in at all. I was probably something of a risky choice, having little background in biology, limited GIS and remote sensing experience, and poor French language skills. My project, as originally conceived, was also something of a risky
endeavor, owing to its heavy emphasis on remote sensing techniques and our need to seek outside assistance. I imagine these risks were clear to Steeve, but he decided to take me on anyways, for which I am very grateful. I had to overcome a steep learning curve and
contact several potential collaborators before real progress could be made, and my penchant for travelling and non-biology related extracurricular activities likely demanded no small amount of patience. Thank you Steeve for keeping me on board and letting me go catch caribou, travel to various conferences, go to Russia and Nunavut, and do my Model UN thing. My time in Quebec City was sometimes difficult because of my struggles with the French language, but ultimately I am happy to have studied here, and will look back on these years as being very formative.
Secondly, I would like to thank the students, postdoctoral fellows, and staff of the Caribou Ungava lab, and of the Côté-Tremblay lab, more broadly. As an introvert and the resident Anglophone, it was not always easy to mix with the group, but I am grateful for the friendship and kindness of many of you. Mael Le Corre, Sabrina Plante, Nicolas Houde, Edouard Bélanger, Mathieu Leblond, and Andrea Panagakis were particularly generous with their time.
Thirdly, I would like to thank Gregory Rickbeil and Nicholas Coops, my co-authors, whose assistance was absolutely essential to me in completing this work. The two months I spent working with them at the University of British Columbia in Vancouver were among the most productive and enjoyable of my degree program. I found their lab, the Integrated Remote Sensing Studio (IRSS), to be a very welcoming environment. Several IRSS graduate students, in addition to Greg, helped me in my work, but Yuhao (Bean) Lu was especially helpful. I am also very much indebted to Txomin Hermosilla, a postdoctoral fellow at IRSS, for his help with the work described in Appendix 1 of this thesis. He developed the Landsat composites used in our attempt to evaluate changes in lichen abundance on the winter range of the Rivière-aux-Feuilles herd.
The Rivière-aux-Feuilles lichen work was challenging and probably the riskiest element of our project, given the well-known difficulties associated with the large-scale mapping of lichen. The fact that I was able to make any progress at all is a testament to the generosity of the many remote sensing specialists and government collaborators who helped me
xi
pursue this work. In addition to the contributions of Txomin Hermosilla, I would like to acknowledge those made by Jérôme Théau, Alexandre Langlois, Claude Morneau, Antoine Leboeuf, and Line Couillard. Jérôme and Alexandre invited me to work in their remote sensing labs at the Université de Sherbrooke and offered their technical expertise. I spent a month working in Jérôme’s lab towards the end of my degree program. I met with Claude and Antoine, of Le ministère des Forêts, de la Faune et des Parcs, early in my studies, and we were able to identify potential avenues for cooperation. Line, of Le ministère du
Développement durable, de l'Environnement et de la Lutte contre les changements
climatiques, met with me several times and provided me with Quebec government data for lichen ground-truth. I regret that I was not able to produce something more substantial from our collaborations, but I am very grateful for your help.
I would also like to thank Jean-Pierre Tremblay and Stéphane Boudreau for evaluating my thesis proposal and serving on my thesis jury.
It would be remiss of me not to mention Peter Lafleur, my undergraduate thesis supervisor at Trent University, who gave me my first opportunity to work as a research assistant in the Arctic. I remember seeing my first caribou at our research camp in the Northwest
Territories and being struck by the beauty of the tundra. Thank you Peter for the chance to visit that place. Although I have since moved away from soil science and will probably move away from caribou biology, I expect I will remain committed to Arctic issues for the rest of my career.
Finally, I would like to thank my parents and grandparents, who have always been very supportive of me in my various academic pursuits. Thank you for everything.
xii
Avant-propos
Ce mémoire inclus un article que nous avons l’intention de soumettre et publier dans un journal scientifique. Cet article est intitulé Remotely-sensed changes in the primary productivity of caribou calving grounds and summer pasture: the mixed influences of climate change and caribou herbivory, mais ce titre est sujet à changement avant la
soumission finale au journal. Cet article a été écrit par A.B. Campeau en collaboration avec G.J.M. Rickbeil, N.C. Coops et S.D. Côté. A.B. Campeau a été responsable de réaliser les analyses et a écrit le manuscrit. M. Le Corre, mentionné dans les remerciements, a partagé les résultats de ses analyses du temps de premier passage provenant d’une étude
précédente; ces analyses ont permis l’identification des dates de départ et d’arrivée des caribous pour différentes saisons. Il a également aidé à préparer les images satellitaires utilisées pour cartographier la productivité primaire sur l’aire d’étude. G.J.M. Rickbeil a conseillé A.B. Campeau sur l’approche méthodologique, principalement pour les questions portant sur la télédétection et les analyses des résidus des modèles. Il a fourni à A.B.
Campeau les données utilisées pour identifier les tendances climatiques et qui ont permis de contrôler pour l’influence du climat sur la productivité primaire dans l’aire d’étude. Il a également partagé le code qu’il a développé lors de travaux antérieurs et a aidé A.B. Campeau à adapter le code pour son étude. Ce code a été utilisé pour créer les bases de données qui ont servies à développer les modèles reliant la productivité primaire aux variables climatiques. N.C. Coops a conseillé A.B. Campeau à propos des éléments de télédétection de l’étude. S.D. Côté a été le directeur de maîtrise de A.B. Campeau et a été fortement impliqué dans la conception et le développement de l’étude. Il a également fourni les bases de données à long terme des localisations de caribous, de la taille de population et du sexe ratio qui ont été utilisées pour les analyses. Tous les co-auteurs ont contribué en révisant le manuscrit final. J.P Tremblay et S. Boudreau ont été les membres du comité d’évaluation de A.B. Campeau.
A.B. Campeau a été supporté financièrement par la bourse Alexander Graham Bell Canada (CGSM) du Conseil de recherches en sciences naturelles et en génie du Canada (CRSNG). Le projet a été financé par les subventions de recherche du CRSNG et d’ArcticNet
décrochées par S.D. Côté, en plus du support financier reçu par les partenaires suivants : Le Ministère du Développement durable, environnement, faune et parcs; Fonds de
recherche sur la nature et les technologies du Québec; Hydro Québec; Glencore Xstrata; La Fédération des pourvoiries du Québec; La Fédération québécoise des chasseurs et pêcheurs; First Air; Société Makivik; The CircumArctic Rangifer Monitoring and Assessment
(CARMA) Network; L'Année polaire international; La Fondation canadienne pour l’innovation; L’Institut pour la surveillance et la recherche environnementales; La Fondation de la faune du Québec; Ouranos; La Fédération canadienne de la faune; Le Gouvernement de Terre-Neuve-et-Labrador; Azimut Exploration; La Conférence régionale
xiii
des élus de la Baie-James; Le Fonds vert du Québec; Tata Steel; Mine Raglan: Une compagnie Glencore; The Torngat Wildlife, Plants & Fisheries Secretariat; Air Inuit; The Grand Council of the Crees; Redevances Aurifères Osisko Ltée.
1
1. Introduction
1.1 Ungulate population dynamics, population size, and range size
Changes in the population size and range of migratory ungulates reflect variations in several intrinsic factors, such as age-specific reproduction and survival, which are in turn influenced by extrinsic, biotic and abiotic factors (Gaillard et al. 2000; Wang et al. 2009). A change in a particular intrinsic factor can influence the balance between the birth and death rates of a population, resulting in either an increase or decrease in population size. Changes in population size can then, in turn, influence range size. A growing population may outstrip available resources in its traditional range, requiring the movement of animals to new, often suboptimal areas, to meet the increase in resource demand, potentially decreasing individual fitness (Pérez-Barbería et al. 2013). Conversely, a shrinking population may require less area to meet its resource needs, allowing animals to occupy optimal areas, potentially increasing individual fitness (Hansen et al. 2009).
1.1.1 The influence of density dependence
Intrinsic factors respond to changes in population density through density-dependent processes and their influence on various biotic factors, resulting in changes to population dynamics. Several biotic factors mediate or exacerbate changes in ungulate population dynamics through negative or positive feedback, helping to either dampen or enhance oscillations in population size, respectively (Saether 1997). These factors include: predation and hunting; disease, parasitism and insect harassment; and forage availability, as exemplified by instances of overgrazing (Gaillard et al. 2000). High density ungulate populations can reduce the availability of forage resources (Côté et al. 2004). Indeed, several possible instances of overgrazing or overbrowsing have been documented in cervid populations (Manseau et al. 1996; Hansen et al. 2007). Mechanical damage through trampling can also occur, enhancing erosion and possibly leading to the development of barren ground areas (Allard 2003). According to the exploitation ecosystem hypothesis
2
(Oksanen et al. 1981), large herbivores can potentially regulate plant biomass, especially in low-productivity environments where predators do not occur in sufficient densities to limit herbivore abundance. Habitat degradation can decrease ungulate survival and reproductive success, ultimately leading to a decrease in population size that may in turn promote vegetation recovery (Klein 1987; Hansen et al. 2007). The situation is complicated by density independent factors, primarily climate and weather, which may interact with density-dependent factors to affect changes in ungulate population dynamics and space use (Pérez-Barbería et al. 2013). One example is the influence of snowfall on winter forage accessibility for northern ungulates, like caribou/reindeer (Rangifer tarandus) and musk-oxen (Ovibos moschatus); deep snow cover increases the energetic costs of cratering, exacerbating any density-dependent food restrictions that might contribute to population declines (Ihl & Klein 2001).
1.2 Ungulate range use, movement patterns, and habitat selection
The range use patterns of migratory ungulates reflect changes in the environment over time and the ability of ungulates to track these changes at various spatiotemporal scales. At a large scale, migratory ungulates move between seasonally changing environments to increase the quality of their habitat, often showing high fidelity to distinct seasonal ranges (Bolger et al. 2008; Mueller et al. 2011). These migratory movements normally occur for any of three primary reasons, as described by Bolger et al. (2008): (1) to track changes in forage abundance and/or quality; (2) to access critically limiting resources, such as salt or fresh water; and (3) to reduce predation pressure, parasitism, and/or insect harassment. When these benefits outweigh the high energetic costs of migratory movements, migration is an effective life history strategy (Hebblewhite et al. 2008; Bischof et al. 2012). At a smaller scale, within seasonal ranges, ungulates make similar compromises in their habitat selection decisions, often having to trade-off energy intake and mortality risk (Hebblewhite & Merrill 2009; Leclerc et al. 2012; van Beest et al. 2013). An examination of the tradeoffs that drive different movement and habitat selection strategies can provide insight into the causes of variability in space use among ungulate populations, leading to a better understanding of ungulate range use. A number of factors are known to drive ungulate range use decisions, but these factors
3
interact in complex ways, and there is limited knowledge regarding the perception and decision-making abilities of ungulates with respect to these interactions (Bolger et al. 2008).
1.2.1 Biotic controls on movement and space use: ecological trade-offs
Ungulates are forced to make certain tradeoffs in their movement and space use decisions to improve overall fitness. Migratory movements involve a tradeoff between the energetic costs of migration and the aforementioned benefits of migration. Other tradeoffs influence habitat selection at the landscape-scale and therefore influence seasonal range use. One major tradeoff is that between forage quality versus forage quantity (Holdo et al. 2009; Ryan et al. 2012). In temperate and northern regions, areas with high plant biomass tend to be at a later stage in their annual growth, and therefore contain less protein than areas of low plant biomass experiencing late onset and rapid growth (Pettorelli et al. 2007). According to the forage maturation hypothesis, the ideal point on the quality-quantity continuum should maximize nutritional gains, with optimal foraging areas having both intermediate plant biomass and high growth rates (Hebblewhite et al. 2008). However, spatial heterogeneity of vegetation community types may also be important in ungulate habitat selection, since ungulates may prefer certain forage species (Pérez-Barbería et al. 2013). Another major tradeoff influencing ungulate movement and space use is that between food reward and predation risk, parasitism or insect harassment (Hebblewhite & Merrill 2009). Optimal foraging areas that attract a high density of herbivores may also attract carnivores and insect pests. Ungulates may therefore be forced to choose between the best foraging areas and the safest ones, with many migratory species choosing to space-away from predators to limit predation risk (Hebblewhite & Merrill 2007).
1.2.2 Abiotic controls on movement and space use: topography and climate
Various abiotic factors can also influence the movement and space use decisions of migratory ungulates, as described here under two broad categories: (1) topography and landscape heterogeneity; and (2) climate. Topographic and climatic variability demonstrate universal
4
importance in their influence on migratory ungulate populations, either by providing the impetus for migratory movements or by influencing landscape-scale space use decisions (Weladji & Holand 2006; Sharma et al. 2009). Topographic variation can result in dramatic climatic differences between nearby areas, i.e. between lowlands and uplands, which can evoke short-distance ungulate migrations to exploit differences in the timing of forage maturation along elevational gradients (Hebblewhite et al. 2008; Bischof et al. 2012). Furthermore, topographic variation can result in intra-range variability in the timing of green-up due to small-scale differences in climate, allowing ungulates to exploit patches of highly productive vegetation throughout their range (Cagnacci et al. 2011; Mysterud et al. 2011). Spatial variation in climatic conditions, whether at the continental-scale or the landscape-scale, is thus a key determinant of ungulate movement and space use strategies. These strategies can also be influenced by inter-annual variation in regional weather (Pérez-Barbería et al. 2013), as well as by directional climate change (Sharma et al. 2009). The extent to which ungulates can adapt to climate change is currently a subject of intense research, with a particular focus on potential mismatches between migratory timing and spring green-up (Post & Forchhammer 2008; Kerby 2013; Middleton et al. 2013; Mysterud 2013).
1.3 Remote sensing for ungulate range assessment
Many of the factors considered important in driving ungulate space use can be evaluated using existing remote sensing tools. Indeed, these tools have already proved instrumental in many ecological studies, allowing researchers to link biological data to remotely-sensed measures of habitat characteristics, including such diverse factors as topography, land cover, climate, and vegetation productivity (Leyequien et al. 2007). Remotely-sensed vegetation indices, particularly the Normalized Difference Vegetation Index (NDVI), have perhaps shown the greatest utility in ungulate management and range assessment, as will be demonstrated in the following section.
5
1.3.1 The Normalized Difference Vegetation Index and other vegetation indices
Remote sensing technology has allowed for the indirect measurement of land cover characteristics, including vegetation productivity, through the analysis of reflectance spectra. Photosynthesis absorbs light in the photosynthetically active range (PAR) of the electromagnetic spectrum, resulting in a distinct reflectance spectrum with reduced PAR radiation relative to other wavelength bands (Pettorelli et al. 2005). The level of absorption in the PAR spectral region can thus be used as an index of photosynthetic activity or vegetation productivity. The commonly used NDVI works on this principle; it is calculated using the following expression, where NIR is reflectance in the near-infrared and VIS is reflectance in the visible (red) region of the electromagnetic spectrum, which closely approximates the PAR wavelength bands (Rouse et al. 1974; Pettorelli et al. 2005):
Satellites equipped with remote sensing radiometers, such as the Landsat and NOAA series satellite platforms, have been accumulating global multispectral data since the 1970s, allowing for the examination of large-scale spatiotemporal changes in environmental characteristics (Pettorelli et al. 2005). This technology has proven incredibly useful in long-term ecological studies, as reflected by a growing body of literature (Leyequien et al. 2007). Of particular importance, examined in greater detail here, is the use of NDVI data to track spatiotemporal changes in vegetation abundance. This approach is particularly valuable in examining vegetation changes spanning continental or regional scales that may be driven by global phenomena like climate change and nutrient deposition. At a large scale, Pouliot et al. (2009) used NOAA Advanced Very High Resolution Radiometer (AVHRR) data to calculate NDVI and examine vegetation changes in Canada between 1985 and 2006. In a similar study, Olthof et al. (2008) used both Landsat and AVHRR-derived NDVI data to examine vegetation changes in Northern Canada between 1986 and 2006, finding significant vegetation productivity increases in most Arctic and boreal regions, in accordance with many other authors (Fraser et al. 2011; McManus et al. 2012; Simms & Ward 2013). In their recent analysis of 1984-2012 Landsat data, Ju and Masek (2016) identified strong positive trends in
6
Arctic and subarctic NDVI, with the most intensive greening occurring in Northern Quebec and Labrador. The utility of NDVI in broad continental or regional-scale studies of vegetation change is thus very well demonstrated, especially in vast wilderness areas like the Canadian North, where huge distances and poor accessibility favor the use of remote sensing techniques.
In addition to the large-scale applications of global change studies, NDVI is also frequently used to examine small-scale vegetation changes and to infer habitat suitability for wildlife populations. NDVI can be used as an index of forage biomass or primary productivity to examine spatiotemporal variation in forage abundance, a key determinant of ungulate movement and space use strategies. Indeed, this index has already been used to study several migratory (or nomadic) ungulate species, including the saiga antelope (Saiga tatarica) (Singh et al. 2010), Mongolian gazelles (Procapra gutturosa) (Ito et al. 2013), and wildebeest (Connochaetes sp.) (Verlinden & Masogo 1997), and has shown promise with populations of reindeer and caribou in Arctic and boreal regions (Thomas et al. 2006; Hansen et al. 2009). Furthermore, NDVI can also be used as an index of forage quality, since the rate of change in NDVI can be linked to the rate of plant growth and subsequently to plant nutrient content, with areas exhibiting rapid increases in NDVI presumably containing rapidly growing, highly nutritious forage (Bischof et al. 2012). Migratory ungulates appear to prefer new plant growth, often tracking waves of green-up across the landscape, making an NDVI forage quality proxy a valuable tool to assess habitat quality (Pettorelli et al. 2007; Bischof et al. 2012). Ryan et al. (2012) found that NDVI can be used as an index of forage nitrogen content in a South African savannah ecosystem, and that correlations can be drawn between this index and African buffalo (Syncerus caffer) body condition. In a study on Bylot Island in Nunavut, Canada, Doiron et al. (2013) demonstrated that NDVI can be used as a proxy for peak nitrogen concentration in some tundra plants.
Beyond the calculation of NDVI for the aforementioned purposes, multispectral data can also be used to estimate the abundance of lichen, which has distinct spectral reflectance properties (Gilichinsky et al. 2011). There has been much interest in developing a remotely-sensed index of lichen abundance to guide northern ungulate management, since caribou/reindeer rely heavily on lichen as winter forage (Gilichinsky et al. 2011). However, several
7
methodological challenges have thus far restricted the accuracy of remotely-sensed lichen assessments (Gilichinsky et al. 2011). Lichen shows no reflectance peak in the green wavelengths (Rees et al. 2004), giving it a unique reflectance signal that excludes the use of NDVI. Areas where lichen co-occurs with green vegetation produce mixed reflectance signals (Gilichinsky et al. 2011). In lichen woodlands, for example, the lichen mat can be partially hidden by the forest canopy, with the level of canopy closure having an important influence on the overall reflectance signal of an area (Théau et al. 2005). Lichen assessment in boreal or tundra-forest ecotone environments is therefore more problematic than in open tundra. Although a number of authors have developed variably successful techniques for lichen classification, as reviewed by Gilichinsky et al. (2011), the remote sensing of lichen requires further methodological development.
1.4 Study context: migratory caribou on the Quebec-Labrador Peninsula
Populations of Rangifer, caribou or reindeer, represent a key component of Arctic and boreal ecosystems throughout their circumpolar range, including in Northern Quebec and Labrador (Vors & Boyce 2009; Festa-Bianchet et al. 2011). As the dominant ungulate grazers in the region, Rangifer represent both the main source of grazing pressure for vegetation communities and the main source of prey for large predators, giving Rangifer populations great importance in ecosystem functioning and in maintaining ecosystem integrity (Festa-Bianchet et al. 2011). Rangifer are also of great socioeconomic and cultural importance for human populations involved in either reindeer herding or caribou hunting activities (Vors & Boyce 2009), but these traditional uses may not be sustainable in the future, given the rapid pace of environmental change in northern regions (Anisimov et al. 2007). Indeed, most wild Rangifer populations have experienced declines in recent years, suggesting that global pressures, like climate change and human development activities, are negatively impacting Rangifer herds and their habitat (Vors & Boyce 2009). These declines will have serious consequences for Arctic and subarctic ecosystems and for traditional Rangifer hunting and herding cultures. It is for these reasons that recent Rangifer population declines are so troubling, providing further impetus for studies on Rangifer populations and their habitat.
8
The focal ungulate populations in this study will be the migratory Rivière-George (RGH) and Rivière-aux-Feuilles (RAFH) caribou herds, which range across much of Northern Quebec and Labrador on Canada’s Quebec-Labrador Peninsula (Bergerud et al. 2008). These two populations represent an important resource for the indigenous Cree, Inuit and Naskapi people, but variations in caribou population size have at times restricted their caribou harvest (Bergerud et al. 2008). At least two large oscillations in the RGH have occurred since the late 1800s, with population peaks followed by relatively lengthy periods of scarcity (Messier et al. 1988; Bergerud et al. 2008). Most recently, the RGH declined from a peak of approximately 776 000 animals in 1993 (Couturier et al. 1996; Couturier et al. 2004) to approximately 14 200 (± 710, 90% CI) in 2014 (Quebec aerial survey). This dramatic collapse occurred after a period of rapid population growth, a seemingly remarkable recovery from the low numbers of the early 1900s. These same population trends can also be inferred from tree-ring data, since caribou activity can be correlated with the extent of conifer root scaring caused by caribou trampling; an increase in the size of the RGH occurred between 1975 and the late 1980s, followed by a rapid decline in the 1990s (Boudreau et al. 2003). The RAFH, meanwhile, increased from about 56 000 animals in 1975 (Le Hénaff 1976) to a peak of at least 628 000 in 2001 (Couturier et al. 2004), with recent surveys suggesting a stable or slowly decreasing population of around 430 000 (± 99 000, 90% CI) animals in 2011 (Quebec aerial survey). The reasons for these changes and associated changes in range use are not well understood, inhibiting our ability to effectively manage the two herds.
1.4.1 Migration patterns and seasonal changes in range use
The RGH and RAFH migrate between different seasonal ranges to track changes in the quality, quantity and accessibility of their preferred forage species, as well as to space-away from apparent competition (i.e. with moose, Alces americanus), predation and insect harassment (Bergerud et al. 2008). In late winter, normally in April, parturient females begin a northward migration from their lichen-rich winter range to their calving grounds, trading the shelter of the boreal forest for the barren tundra plateaus where they traditionally give birth, usually in early June (Bergerud et al. 2008). The traditional calving grounds occur at
9
relatively high elevations for the RGH, most probably as a tactic to avoid predation, since wolves (Canis lupis) and black bears (Ursus americanus) are known to primarily utilize lower elevation areas (Apps et al. 2013). Caribou arrival on the calving grounds roughly coincides with the appearance of nutritious forage during spring green-up, with caribou mothers primarily feeding on new protein-rich graminoid growth to promote milk production (Bergerud et al. 2008). After a few days or weeks, the caribou leave their calving grounds for their broader summer range, often gathering in large aggregations to reduce individual predation risk and insect harassment (Bergerud et al. 2008). They feed on birch and willow shrubs, as well as many other plants, including graminoids (Crête et al. 1990; Bergerud et al. 2008). As winter approaches, caribou migrate south to their winter range in the lichen woodlands, where the partial forest canopy offers some shelter from harsh winter conditions (Bergerud et al. 2008). The lichen woodlands provide access to carbohydrate-rich lichen for winter grazing; caribou dig through the snow to feed on fruticose ground lichens and certain evergreen shrub species (Bergerud et al. 2008). Upon the return of spring, caribou again begin their annual migratory cycle, moving north to exploit the increase in summer productivity on the tundra.
1.4.2 Knowledge gaps: understanding interannual changes in range use
Over the last few decades, the RGH and RAFH have demonstrated considerable changes in their use of seasonal ranges, complicating efforts to manage the two populations. Both herds have undergone large changes in population size over this time, potentially explaining much of the observed variation in range use, but the mechanisms driving range use changes are largely unknown. Changes in forage availability and predation pressure interact with changes in population size through complex density-dependent interactions (Wang et al. 2009), with density independent forcing – such as climate change and extreme weather – adding further complexity (Tyler 2010). It has been well documented, however, that forage availability plays a key role in determining the ‘carrying capacity’ of migratory caribou habitat, with authors attributing variable importance to the summer and winter ranges in restricting population size (Messier et al. 1988; Manseau et al. 1996; Ferguson et al. 2001). For the
10
RGH, Manseau et al. (1996) found evidence to indicate that caribou overgraze their calving grounds and summer range during periods of peak population size, suggesting a need for large caribou herds to occupy larger or more productive areas. Indeed, Taillon et al. (2012) observed an increase in the size of the RGH calving grounds between 1974 and 1988 as the population expanded, followed by a range contraction after the population declined, suggesting overcrowding and overgrazing as probable causes for range expansion. Messier et al. (1988) also noted an increase in the size of the RGH annual range coincident with the 1971-1984 period of population increase. The RAFH, in contrast, has exhibited limited variability in calving ground use (Taillon et al. 2012) and appears more demographically stable than the RGH. It has, however, demonstrated an interesting change in its winter range use over the past few years, with individuals migrating further south to utilize a much larger winter range than they had previously (Le Corre 2016). This change could be a result of the RAFH moving to occupy the areas used by the RGH before its population crash and subsequent range contraction. However, it is also possible that the growing RAFH overgrazed its former winter range, reducing lichen abundance and thus necessitating the shift in range use.
1.4.3 Knowledge gaps: understanding the role of caribou herbivory in habitat change
Caribou herbivory can have transformative effects on vegetation communities subjected to grazing or browsing pressure, especially if caribou occur at high densities for several consecutive years (Newton et al. 2014). An obvious consequence of caribou herbivory can be a decrease in the biomass of preferred forage resources, like lichen or grasses and deciduous shrubs, with subsequent increases in the relative abundance of less palatable plant species (Manseau et al. 1996). The resulting changes in vegetation community composition and forage availability may reduce habitat quality for caribou and potentially precipitate declines in caribou abundance (Klein 1987; Hansen et al. 2007). However, the influence of caribou herbivory on habitat change can be difficult to distinguish from other factors, like climate change and nutrient deposition, which also influence Arctic and subarctic vegetation (Anisimov et al. 2007). Recent circumpolar increases in primary productivity appear to be
11
strongly associated with observed increases in the cover and biomass of deciduous shrubs, namely dwarf birch and willow species, in a process known as shrubification (Tremblay et al. 2012; Ropars & Boudreau 2012). This trend towards increases in shrub cover and biomass is most probably a consequence of climate warming (Sturm et al. 2001; Myers-Smith et al. 2011), but other changes, like regional declines in caribou abundance, could also have a positive effect on shrub growth. Shrubification and general increases in Arctic and subarctic primary productivity could improve green forage availability for caribou, but additional changes in plant nutritional quality or caribou diet may also prove important (Thompson & Barboza 2014). An improved understanding of the roles of caribou herbivory in influencing forage quality and quantity is needed to effectively manage caribou populations and respond to the challenges of climate change.
12
1.5 Objectives
1.5.1 General objectives
This study aims to evaluate the extent to which interannual variation in the range use of migratory caribou can be explained by demographic trends and spatiotemporal changes in forage abundance. It will also examine the extent to which caribou herbivory can influence primary productivity and impact the abundance of forage resources.
1.5.2 Specific objectives
1. Explain interannual variation in the size and location of the RGH calving grounds; 2. Examine the influence of demographic trends on green forage abundance to clarify
the impacts of caribou herbivory on the RGH calving grounds and summer range; 3. Evaluate the influence of lichen abundance in driving recent changes in the location
13
2. Remotely-sensed changes in the primary productivity of migratory caribou calving grounds and summer pasture: the mixed influences of climate change and caribou herbivory
A. Brett Campeau1, Gregory J.M. Rickbeil2, Nicholas C. Coops2 and Steeve D. Côté1 1Caribou Ungava, Département de Biologie, Université Laval, Québec, Québec, Canada 2Integrated Remote Sensing Studio, Department of Forest Resources Management,
14
Résumé
À haute densité, la pression de broutement des herbivores peut être un facteur suffisant pour réduire l’abondance des ressources alimentaires et potentiellement contribuer à la dégradation de l’habitat. Ces modifications pourraient engendrer des changements de taille de la population des herbivores ou de leur aire de distribution. L’hypothèse des écosystèmes exploités (EEH) suggère que l’herbivorie a un rôle régulateur dominant sur la biomasse végétale des environnements de faible productivité, comme la toundra et la taïga, où la disponibilité des ressources alimentaires pourrait être un facteur limitant la densité des herbivores. Le troupeau de caribous migrateurs Rivière-George (TRG) de la péninsule du Québec-Labrador (Canada) a subi un large déclin de sa taille de population depuis un pic au début des années 1990, parallèlement avec de grands changements d’utilisation des aires saisonnières. Ces changements démographiques sont suspectés avoir influencé la disponibilité des ressources alimentaires et les stratégies d’utilisation de l’habitat du caribou via des interactions dépendantes de la densité entre le caribou et son habitat. Nous avons utilisé le Normalized Difference Vegetation Index (NDVI) afin d’examiner les relations entre la densité de caribou du TRG et la productivité primaire entre 1991 et 2011. Nous avons utilisé une approche de modélisation liant le NDVI à des variables climatiques pour contrôler l’influence du climat et isoler le rôle du broutement sur la productivité primaire. La densité de caribous et la productivité primaire étaient négativement corrélées pour l’aire de mise bas et l’aire d’estivage du TRG. Les tendances temporelles de la productivité primaire semblent refléter le déclin de population du TRG, ce qui suggère un rétablissement de la végétation suite à la réduction de l’abondance du caribou. Ces résultats supportent l’EEH en démontrant que la pression de broutement peut réduire la productivité primaire dans les environnements de faible productivité. Le réchauffement climatique est le principal responsable des augmentations de la productivité primaire observées entre 1991 et 2011, mais la réduction de la pression de broutement du TRG pourrait également avoir contribuée à ces augmentations. Les changements démographiques et d’habitat du TRG pourraient avoir augmenté l’accessibilité aux ressources alimentaires pour les individus et ainsi expliquer l’amélioration de leur condition corporelle, ce qui en retour pourrait avoir accentué le rôle de l’évitement de la prédation en tant que mécanisme de sélection d’habitat à large échelle.
15
Abstract
At high density, herbivory may be sufficient to reduce forage abundance, potentially contributing to habitat degradation and driving changes in herbivore population size or range use. Indeed, the exploitation ecosystem hypothesis (EEH) proposes that herbivory may have a dominant role in regulating plant biomass in low-productivity environments, like tundra and taiga, with forage abundance potentially limiting herbivore density. The migratory Rivière-George caribou herd (RGH) of the Quebec-Labrador Peninsula (Canada) has experienced a large decline in population size since the population peaked in the early 1990s, with similarly large changes in seasonal range use. Demographic changes are suspected to have influenced forage abundance and caribou range use through density-dependent interactions between caribou and their habitat. We used the Normalized Difference Vegetation Index (NDVI) to examine the relationships between RGH caribou density and range productivity over the 1991-2011 period. A modelling approach relating NDVI to climatic variables was used to control for the climate signal and isolate the influence of caribou herbivory on primary productivity. Significant negative relationships were identified between caribou density and primary productivity, after controlling for climatic variation, for the global RGH calving grounds and summer range. Positive temporal trends in primary productivity appeared to reflect the decline in RGH population size, suggesting vegetation recovery following reductions in caribou abundance. These results demonstrate that caribou herbivory can reduce primary productivity, providing support for the EEH prediction of top-down herbivory effects in low-productivity environments. Climate warming was clearly most responsible for the strong positive trends in primary productivity observed over the 1991-2011 period, but decreases in RGH herbivory also contributed to the increases in range productivity. Demographic and habitat changes likely allowed for improved forage access and subsequent increases in caribou body condition, which may have influenced RGH range use by increasing the relative importance of predation avoidance in driving large-scale habitat selection.
16
2.1 Introduction
Arctic terrestrial ecosystems exhibit slow vegetation dynamics because of harsh climatic and soil conditions, giving Arctic herbivores like caribou (or reindeer, Rangifer tarandus) an important role in governing ecosystem functioning (Post & Pedersen 2008). These low-productivity ecosystems may function according to the exploitation ecosystem hypothesis (EEH), which postulates that vegetation is regulated by top-down herbivory in the absence of significant predation pressure (Fretwell 1977; Oksanen et al. 1981). The extent to which low-productivity terrestrial ecosystems are regulated by herbivory is debatable, however, because of the varying degrees of predation intensity and the confounding effects of climate change on vegetation productivity (Legagneux et al. 2012). It is therefore necessary to consider the influences of climatic and environmental variation when assessing plant-herbivore relationships in low-productivity environments. For migratory caribou herds, migration to relatively predator-free calving grounds and summer areas helps to reduce predation risk (Fryxell & Sinclair 1988; Heard et al. 1996; Bergerud & Luttich 2003), but the low primary productivity and slow vegetation dynamics of these areas make them susceptible to overgrazing, especially when caribou are abundant (Allard 2003). These environments thus represent appropriate test areas for the EEH prediction of a top-down regulatory role for herbivory in low-productivity areas. A large-scale and long-term examination of herbivory effects can help to shed light on complex plant-herbivore
relationships, perhaps linking herbivore population fluctuations with vegetation degradation and recovery (Hansen et al. 2007; Newton et al. 2014).
Most migratory caribou populations in North America are currently in a state of decline, possibly suggesting a general deterioration of habitat conditions and subsequent reductions in caribou survival and fecundity (Vors & Boyce 2009; Festa-Bianchet et al. 2011). The decline phase is arguably part of natural population fluctuations, which for many herds appear to follow the same, predictable pattern: a low-abundance phase followed by rapid growth, a high-abundance plateau, and, when habitat conditions deteriorate, a subsequent collapse of the population (Messier et al. 1988; Couturier et al. 1990). These fluctuations are thought to be driven by several interacting factors, including predation by gray wolf (Canis lupus) (Heard et al. 1996; Bergerud & Luttich 2003), climatic variability and
17
extreme weather (Klein & Shulski 2009; Tyler 2010), and perhaps most importantly, changes in caribou forage availability (Couturier et al. 2009). Caribou herbivory likely contributes to habitat degradation and thus leads to declines in caribou abundance, which could in turn promote vegetation recovery and an eventual return to high caribou density. The length of time necessary for vegetation recovery will vary depending on the intensity and duration of caribou herbivory and the resilience of the affected vegetation community, potentially introducing temporal fluctuations in forage availability (Newton et al. 2014). Density-dependent feedback between caribou and their preferred forage species may therefore have important consequences for caribou population dynamics.
Changes in the size of a caribou population can affect changes in its range use through density-dependent interactions between caribou, predators, and their habitat. Migratory caribou travel between different seasonal pastures to avoid predation and access forage, often congregating in large numbers and demonstrating high fidelity to specific areas or movement corridors (Fryxell & Sinclair 1988; Bergerud et al. 2008). Parturient females migrate from boreal winter pastures to small, well-defined calving grounds on the tundra portion of their range, typically giving birth in early spring to coincide with snowmelt and the emergence of nutrient-rich vegetation (Post et al. 2003; Post & Forchhammer 2008). Caribou also aggregate in large groups during the summer period to reduce insect
harassment (Toupin et al. 1996). These behaviours can result in caribou occurring at very high densities, potentially reducing local forage abundance through acute foraging and trampling pressure (Manseau et al. 1996). Intense herbivory may ultimately degrade caribou habitat and force caribou to move to areas where forage is more readily accessible, but where predation risk might also be higher (Bergerud & Luttich 2003). The relative importance of predation avoidance and forage access in driving habitat selection may therefore vary in response to caribou population dynamics and associated changes in forage availability.
Forage availability has been shown to limit caribou populations, with different authors attributing primary importance to either the availability of lichens on the winter range (Skogland 1986; Ferguson et al. 2001) or the availability of green forage (i.e. shrubs, herbs, and grasses) on the summer range and calving grounds (Crête & Huot 1993; Crête &
18
Doucet 1998). Lichen degradation has been observed on reindeer winter pastures in Fennoscandia (Colpaert & Kumpula 2012; Kumpula et al. 2014) and on the winter ranges of wild insular Rangifer herds (Klein 1987; Hansen et al. 2009), suggesting a strong
relationship between caribou density and winter forage availability. Lichen degradation can also be observed on the summer ranges of migratory caribou herds (Boudreau & Payette 2004; Théau & Duguay 2004), but lichen is considered less important than green forage for parturient and lactating females during the critical calving and summer foraging periods (Crête et al. 1990; Crête & Huot 1993). Indeed, observational and fecal analysis studies from Alaska (USA) and the Yukon (Canada) have shown that deciduous shrubs, namely dwarf birch and willow species, amount to between 45 and 50% of caribou diets in the spring and summer months (Trudell & White 1981; Boertje 1984; Russell et al. 1993). The hypothesis of green forage limitation is thus highly tenable for continental migratory caribou herds with relatively small calving grounds or low-productivity summer areas. Earlier work has demonstrated calving ground and summer range degradation during periods of high caribou population size, with localized field measurements showing decreases in green forage biomass (Manseau et al. 1996) in association with observed changes in caribou population dynamics (Messier et al. 1988; Couturier et al. 1990). The negative effects of caribou herbivory on forage species have also been demonstrated using experimental studies; caribou exclosures exhibit higher levels of green forage abundance than nearby unenclosed areas (Pajunen et al. 2008; Zamin & Grogan 2013). These field studies have helped to define the immediate link between caribou and vegetation dynamics, but our understanding of long-term and large-scale interactions remains limited. Remote sensing approaches may be better suited to assessing the relationship between caribou and vegetation dynamics over broad spatiotemporal scales relevant to migratory caribou. Satellite-derived indices of vegetation abundance, such as the Normalized Difference Vegetation Index (NDVI), have proven extremely valuable in studies of global change and ungulate habitat selection. A general greening trend has been observed in the Arctic, reflecting the positive effects of climate warming and nutrient deposition on primary productivity and shrub growth (Pouliot et al. 2009; Fraser et al. 2011; Ju & Masek 2016). Other remote sensing studies have demonstrated the influence of forage abundance on ungulate movement patterns, showing a preference by ungulates for highly productive or
19
nutrient-rich forage patches (Mueller et al. 2008; Hansen et al. 2009). However, the extent to which ungulates influence forage abundance is considerably more difficult to assess and has received less attention (but see Boudreau et al. 2003; Côté et al. 2004; Hansen et al. 2007). In recent studies, Newton et al. (2014) and Rickbeil et al. (2014) were able to identify caribou herbivory effects on forage abundance using remotely-sensed indices of vegetation productivity. Remote sensing can thus play a valuable role in evaluating
herbivory effects on vegetation, as well as the influence of vegetation changes on ungulate range use and population dynamics.
We investigated the relationships between primary productivity, caribou population size, and range use by the Rivière-George caribou herd (RGH) in Northern Quebec and Labrador, Canada. This herd experienced a 93% decline in population size between 1991 (~774 265 individuals) and 2011 (~53 980 individuals), and has demonstrated a similarly large decline in the size of its calving grounds over the same period; Taillon et al. (2012) observed a >85% decrease in RGH calving grounds size from 42 800 (SE: 7 500) to 5 930 (SE: 730) km2
between the early 1990s and the 2006-2010 period. These changes likely reflect density-dependent linkages between caribou and their habitat. The RGH has access to a very large winter range compared to its relatively small calving grounds and summer range, leading some authors to propose green forage abundance as the main limiting factor for RGH population growth (Manseau et al. 1996; Crête & Doucet 1998). These same authors provide evidence of habitat degradation on the RGH calving grounds and summer range, but the range-scale relationship between caribou density and green forage abundance remains poorly quantified. We used remote sensing tools and a long-term dataset of caribou locations and population size to assess this relationship for the RGH calving grounds and summer range. We also aimed to clarify the role played by habitat degradation in the RGH population collapse and observed range use changes, as well as the potential for vegetation recovery following a reduction in caribou herbivory. This knowledge will help inform range-scale management responses to future changes in caribou abundance.
20
2.2.1 Study area
Our study area covered the global calving grounds and summer range of the RGH between 1991 and 2011, an area straddling the Quebec-Labrador border in northeastern Canada (Figure 2.1). The area was primarily located within the Taiga Shield Ecozone and included a portion of the Arctic Cordillera Ecozone in the Torngat Mountains (Wiken 1986). This area encompassed a mosaic of land cover types from open forests and wetlands at lower elevations to shrub heaths and bare uplands at higher elevations. The tree line ran north-south along the eastern and western edges of the Torngat Mountains, with trees penetrating into higher areas along major river valleys (Payette, 1983; CAVM Team, 2003). Black spruce (Picea mariana) was the dominant tree species, while white spruce (P. glauca) and eastern larch (Larix laricina) were also common. Betula glandulosa was the dominant shrub species and Cladina stellaris was the dominant lichen species. Vegetation dynamics were slow because of the harsh climate and poor soil conditions (Auger & Payette 2010). Average annual temperature for the 1971-2000 period, reported from the Kuujjuaq climate station (58°06' N, 68°25' W), was −5.7 °C, with average July and January temperatures of 11.5 °C and −24.3 °C, respectively (Environment Canada). Average annual precipitation was 526.8 mm, including 249.6 mm of snow (expressed as snow water equivalent) (Environment Canada). Northern Quebec and Labrador (north of 55°N) has experienced significant climate warming since temperature records began; by the early 2000s, mean seasonal temperatures (for all seasons) had increased by ~2 °C relative to the 1961-1990 reference period (Allard & Lemay 2012). The study area was underlain by sporadic and discontinuous permafrost in the lower-lying, southerly areas and by continuous permafrost in the Torngat Mountains (Hachem et al. 2009).
21
Figure 2.1. The global calving grounds and summer range of the Rivière-George caribou
herd over the 1991-2011 period, as defined by minimum convex polygons (MCPs), clipped to the coastline; the calving grounds MCP is based on the locations of collared females, whereas the summer range MCP is based on the locations of both collared males and females.
2.2.2 Caribou range delimitation
We defined the seasonal ranges of the RGH using the locations of animals equipped with Argos satellite-tracking collars (Telonics, Argos platform, Mesa, Arizona, USA). Caribou were captured using a net-gun fired from a helicopter, with most captures occurring on the RGH calving grounds in early June. Capture and collaring efforts were focused on areas of known caribou use based on existing telemetry or aerial survey data. The capture process followed the guidelines published by the Canadian Council on Animal Care and is described in greater detail by Bookhout (1996). A total of 284 individuals (54 males and 230 females) were equipped with Argos collars and tracked over the 1991-2011 period for a total of 740 caribou-years of location data, with data from an average of 35 individuals in any given year.
22
The satellite collars reported locations every 6 days, on average. The locations had an average precision of better than 700 m. Anomalous locations were excluded using an algorithm similar to that used by Austin et al. (2003), as described by Le Corre et al. (2014). The most accurate locations for a given transmission period were selected based on signal quality, and all locations representing displacements greater than 50 km per day were excluded (Boulet et al. 2007).
We identified the dates of arrival and departure to and from the calving grounds and summer range for each collared individual using first-passage time (FPT) analysis (Le Corre et al. 2014). This method examines the time required for an individual to move from the centre to the exterior of a circle of a predetermined radius, with shorter times indicating migratory movements and longer times indicating more localized, foraging movements. We excluded individuals that did not show clear arrival or departure dates from our analyses (10% and 17% of individuals for the calving grounds and summer range, respectively). We used data from all collared RGH individuals (both males and females) to define the RGH summer range. However, we only used data from collared females to define the calving grounds, since only parturient females are known to reliably migrate to the calving grounds in the spring (Gunn & Miller 1986; Bergerud et al. 2008). For the 1991-2011 period, we successfully identified calving grounds and summer ranges for 306 and 341 caribou-years, respectively, for an average of 15 individual calving grounds and 16 individual summer ranges in each year.
We delimited the annual calving grounds and summer ranges of the RGH for the 1991-2011 period using minimum convex polygons (MCPs) and Brownian Bridge Movement Models (Horne et al. 2007; hereafter Brownian Bridges, or BBs). We created 100% MCPs and 95% Brownian Bridge home ranges using the adehabitat package (Calenge 2006) in R (R Foundation for Statistical Computing, Vienna, Austria; R Core Team 2014). The MCPs were used to conservatively define the global 2011 calving grounds and the global 1991-2011 summer range based on the locations of all collared animals over the 1991-1991-2011 period. The Brownian Bridges were used to identify areas of intensive caribou use, or core areas, based on the extent of Brownian Bridge overlap occurring in an area over a given time period. Annual calving period and summer period Brownian Bridges were created for each collared
23
animal with identifiable calving and summer periods. Core calving and summer areas were then defined as those areas with ≥ 70% of the maximum number of Brownian Bridge overlaps from all seasonal ranges in the time period of interest. Core areas were examined for different multiyear periods based on observed multiyear tendencies in seasonal range use.
2.2.3 Caribou population size and density estimation
We produced annual RGH population size estimates by fitting a loess smoothing spline to existing population data, collected using aerial survey techniques (Quebec and Newfoundland-Labrador government aerial counts), and then extracting the predicted values from the spline (Figure 2.2); the loess function in R was used. Annual density estimates for the global 1991-2011 calving grounds and summer range MCPs were then produced by dividing the annual RGH population size estimate by the land area (km2) of the global MCP.
Densities reported for the global calving grounds refer only to adult females, whereas densities for the global summer range refer to all individuals. Sex ratio data were used to estimate the abundance of adult females (Caribou Ungava, unpublished data). We also estimated caribou densities in the RGH core calving and summer areas for each year of the 1991-2011 study period based on the number of Brownian Bridge overlaps occurring in a given area. For the core summer areas, we assumed that areas with complete overlap of all Brownian Bridges from all collared individuals were visited by all RGH caribou in that season, for a density equivalent to the RGH population size divided by the size of the area. Our approach was the same for the core calving areas, except the population estimate was adjusted to only consider adult females. The core area density calculations were conducted in ArcGIS (Esri, Redlands, California, USA) on a by-pixel basis according to Equation 2.1, where Proportion refers to the proportion of the number of Brownian Bridges overlapping the pixel relative to the total number of collared caribou in that season.
24
Figure 2.2. A loess smoothing spline fitted to aerial survey estimates of Rivière-George
caribou herd population size (black data points) to produce annual population size estimates. Error bars represent confidence intervals (α = 0.10) associated with the aerial survey data. The gray shaded area represents the 1991-2011 study period.
2.2.4 Remotely-sensed data processing
We used 10-day Advanced Very-High Resolution Radiometer (AVHRR) composites to calculate the NDVI and evaluate interannual changes in the primary productivity of the study area over the 1991-2011 study period. The AVHRR composites consist of cloud-free or nearly cloud-free images, available at 10-day intervals, with a nominal spatial resolution of 1 km. They were created using the Earth Observation Data Manager (EODM) AVHRR data processing system described by Latifovic et al. (2005). NDVI has proven to be a reliable index of primary productivity in a wide variety of habitats, including tundra and taiga environments, making it well-suited to this study (Stow et al. 2004; Pettorelli et al. 2005). We used cumulative annual growing season NDVI (cNDVI) as an index of primary productivity. This index allowed us to examine the snow-free period in its entirety, removing any influence of phenology on interannual comparisons of primary productivity. We calculated cNDVI on a by-pixel basis by summing all NDVI values over 0.05, providing a single value for each pixel in each year. We chose the 0.05 threshold after examining the