Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 270
Research Article
Anthropisation et effets de lisière : impacts sur la
diversité des rongeurs dans la Réserve Forestière de
Masako (Kisangani, R.D. Congo).
Léon Iyongo Waya Mongo
1,2,5*, Marjolein Visser
1, Charles De
Cannière
1, Erik Verheyen
3, Benjamin Dudu Akaibe
4, Joseph Ulyel
Ali-Patho
5et Jan Bogaert
21
Service d’Ecologie du Paysage et Systèmes de Production Végétale, Université Libre de Bruxelles, Av. Franklin Roosevelt 50, CP 169, Bruxelles 1050, Belgique.
2
Unité Biodiversité et Paysage, Gembloux Agro-Bio Tech, Université de Liège, Passage des Déportés 2, B-5030 Gembloux, Belgique.
3
Vertebrate Department, Royal Belgian Institute of Natural Sciences, Vautierstraat 29, 1000 Brussels, Belgium. 4
Laboratoire d’Ecologie et de Gestion des Ressources Animales, Faculté des Sciences, Université de Kisangani, B.P. 2012, R.D. Congo.
5
Section des Eaux et Forêts, Institut Supérieur d’Etudes Agronomiques de Bengamisa, BP 202 Kisangani, R.D. Congo. *Corresponding author ([email protected])
Résumé
Nous avons évalué l’impact anthropique sur la richesse, la diversité, l’abondance et la densité relative des Rongeurs dans quatre habitats de la Réserve de Masako au nord-est de la République Démocratique du Congo. Une forêt primaire à Gilbertiodendron dewevrei et trois habitats d’origine anthropique (forêt secondaire, jachère et zone de contact jachère/forêt secondaire (lisière)) ont été investigués de mai 2009 à avril 2010. 1275 spécimens répartis entre 24 espèces et 5 familles inventoriées ont permis de quantifier des différences de composition et d’abondances entre un habitat non perturbé et des milieux anthropisés, ainsi qu’entre une lisière et des habitats adjacents. Nos résultats montrent une distribution non uniforme des Rongeurs dans les quatre milieux. La richesse et la diversité sont élevées en lisière etfaibles en forêt primaire. Chaque milieu héberge une densité relative différente. Les plus fortes densités sont observées en forêt secondaire et jachère ; la plus faible est observée en forêt primaire. Les abondances diffèrent significativement entre les habitats, à l’exception de la paire forêt secondaire - jachère. Les paramètres dans la lisière diffèrent de ceux des habitats adjacents, traduisant l’existence d’un effet de lisière. Les équitabilités faibles obtenues dans tous les habitats traduisent l’instabilité relative de cet écosystème forestier. Le test de χ² atteste l’existence de l’effet « saison » sur les abondances des Rongeurs. De même, ce test confirme l’existence de l’effet « activités anthropiques » sur les abondances dans les milieux anthropisés, mais pas en forêt primaire.
Mots clés : Anthropisation, effets de lisière, diversité, Rongeurs, Masako. Abstract
Anthropogenic effects on rodent richness, diversity, abundance, and relative density were studied in 4 habitats of the Masako Reserve, situated in the northeastern part of the Democratic Republic of the Congo. A primary forest dominated by Gilbertiodendron dewevrei as well as 3 anthropogenic habitats (secondary forest, fallow land, and the edge zone situated between the fallow land and the secondary forest) were sampled for rodents between May 2009 and April 2010. 1275 specimens (24 species and 5 families) were captured to quantify the differences in composition and abundance between the non disturbed habitat and the anthropogenic habitats, and between the edge zone and its adjacent habitats. Results indicated a non uniform rodent distribution across the four habitats. Richness and diversity were high in the edge habitat and low in the primary forest. Every habitat was characterized by a different relative density, of which the highest values were observed for the secondary forest and for the fallow habitat; the lowest value was observed for the primary forest. Edge zone characteristics differed from the adjacent habitats which suggested the existence of edge effects. Abundances differed significantly between habitats except between the secondary forest and the fallow land. The low evenness values observed in all habitats reflected the relative instability of the ecosystems at study. A chi-squared test confirmed the existence of seasonal effects on rodent abundance; the impact of anthropogenic activities on rodent presence was shown in the same way for the anthropogenic habitats but not for the primary forest habitat.
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 271
Introduction
Les écosystèmes forestiers tropicaux n’échappent pas à la déforestation malgré leur grande complexité, leur remarquable diversité et leur importante richesse de formes biologiques. Ils sont très menacés de dégradation suite aux divers facteurs tels que le changement climatique régional [1] et les types d’activités anthropiques qui y sont pratiquées, notamment l’agriculture itinérante sur abattis-brûlis [2], l’exploitation du bois sous diverses formes (bois de chauffe, bois d’œuvre et d’industrie), la cueillette et la chasse, l’urbanisation, l’installation des infrastructures de développement et l’exploitation minière [3]. Face aux menaces que les activités anthropiques font peser sur ces écosystèmes, une préoccupation croissante se développe autour des problèmes de perte et de dégradation de la biodiversité [3, 4]. C’est ainsi que les études liées au phénomène de déforestation se multiplient, en vue d’évaluer l’impact de la dégradation et de la fragmentation accélérée des écosystèmes sur la biodiversité des habitats et son évolution dans l’espace et dans le temps. Cette fragmentation se traduit par l’augmentation de la proportion des lisières qui sont des zones situées à l’interface d’habitats contrastés et sur lesquelles s’exercent les « effets de lisière » [5, 6].
Les Rongeurs sont parmi les petits Mammifères, ceux qui contribuent le plus à la diversité spécifique d’un écosystème. Ils interagissent intensivement avec leur environnement et ont des effets complexes sur les autres organismes (végétaux et animaux) [7]. Ils forment un maillon important dans les réseaux trophiques et constituent une ressource alimentaire représentant une proportion non négligeable du gibier dans plusieurs contrées africaines [3]. Vu leur abondance et leur dépendance vis-à-vis des plantes et des insectes dont ils se nourrissent, ils sont des victimes directes des actions anthropiques [8]. Leur forte association à des habitats particuliers dont témoignent certaines évidences paléontologiques et écologiques fait d’eux d’intéressants indicateurs du changement des conditions du milieu et de la fragmentation des habitats [9, 10]. De même, il est admis que des changements rapides dans les écosystèmes naturels conduisent à la dominance de quelques espèces opportunistes [11].
La présente étude a pour but de vérifier les hypothèses selon lesquelles les activités humaines pratiquées à Masako auraient un impact sur la distribution de la diversité spécifique, des abondances et des densités relatives des Rongeurs entre les différentes taches d’habitat dans cet écosystème forestier. Le milieu non perturbé compterait une diversité, une abondance et une densité relative différentes de celles de chaque milieu anthropisé et présenterait une très faible similarité avec ces derniers.Comme l’ont montré certains auteurs [5, 6], la zone de lisière hébergerait une richesse et une diversité plus élevées. Cette zone se caractériserait par des abondances et des densités de Rongeurs différentes de celles des habitats adjacents. Face aux conditions changeantes du milieu [10], les espèces les mieux adaptées à un type d’habitat y deviendraient dominantes.
Received: 26 January 2012; Accepted: 20 february 2012; Published: 17 September 2012.
Copyright: © Léon Iyongo Waya Mongo, Marjolein Visser, Charles De Cannière, Erik Verheyen, Benjamin Dudu Akaibe, Joseph Ulyel Ali-Patho and Jan Bogaert . This is an open access paper. We use the Creative Commons Attribution 3.0 license
http://creativecommons.org/licenses/by/3.0/ - The license permits any user to download, print out, extract, archive, and distribute the article, so long as appropriate credit is given to the authors and source of the work. The license ensures that the published article will be as widely available as possible and that the article can be included in any scientific archive. Open Access authors retain the copyrights of their papers. Open access is a property of individual works, not necessarily journals or publishers.
Cite this paper as: Iyongo Waya Mongo, L., Visser, M., De Cannière, C., Verheyen, E., Dudu Akaibe, B., Ulyel Ali-Patho, J., and Bogaert, J. 2012. Anthropisation et effets de lisière : impacts sur la diversité des rongeurs dans la Réserve Forestière de Masako (Kisangani, R.D. Congo).
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 272
C’est pourquoi, nous avons entrepris une étude sur les effets des activités anthropiques durant une période de 12 mois dans la Réserve Forestière de Masako localisée à Kisangani en République Démocratique du Congo. La Réserve est l’un des blocs forestiers importants des environs de Kisangani et abrite une gamme importante de petits Mammifères [3]. Ceux-ci sont particulièrement reconnus comme sensibles aux transformations de leurs habitats [10]. La présente étude établit des interactions entre les activités humaines et la distribution de la biodiversité des Rongeurs.
Méthodes
La présente étude, effectuée sur autorisation de la Coordination Provinciale de l’Environnement et Conservation de la Nature en Province Orientale (RD Congo), a été conduite de mai 2009 à avril 2010 dans la Réserve Forestière de Masako (0°36’N et 25°13’E), d’une superficie de 2.105 ha et située à 15 km de la ville de Kisangani au nord-est de la République Démocratique du Congo [12, 13]. Trois types de pièges appâtés à la pulpe de noix de palme et au manioc cru ont permis de capturer les animaux : les clapettes du type « Lucifer Rat Traps », les pièges Shermans du type « Life folding traps » en aluminium et les pièges traditionnels fabriqués à l’aide de bâtonnets et de ficelles.
Le dispositif d’échantillonnage était constitué de quatre grilles de capture installées respectivement dans la jachère (J), la zone de lisière (L), la forêt secondaire (Fs) et la forêt primaire à
Gilbertiodendron dewevrei (Fp) (Fig. 1). Chaque grille (1 ha) comptait 120 pièges (50 clapettes, 50
Shermans et 20 pièges traditionnels). Une distance de 300 m était maintenue entre les grilles J, L et Fs dans le but de disposer d’observations indépendantes. La grille Fp, qui sert de témoin à la
comparaison avec les milieux anthropisés, était installée à 1,5 km en raison du recul de la forêt vierge face à la pression anthropique.
Fig. 1. Réserve Forestière de Masako (0°36’N ; 25°13’E), Kisangani, République Démocratique du Congo avec indication des 4 dispositifs d’échantillonnage chacun constitué d’une grille (1 ha/grille ; 120
pièges/grille avec orientation N-S ; 10 m de distance entre les pièges Sherman (a) et Lucifer (b) placés avec alternance; 20 m de distance entre les pièges traditionnels (c) intercalés entre les lignes des Sherman et Lucifer. (Fp) forêt primaire à
Gilbertiodendron dewevrei ; (Fs) forêt
secondaire ; (L) lisière (jachère/forêt secondaire) ; (J) jachère. Adapté de Dudu (1991), avec permission.
La végétation de la grille Fp est caractérisée par la dominance de Gilbertiodendron dewevrei (Caesalpiniaceae) accompagnée de quelques essences comme Polyanlthia mavelolens, Strombosia
glaucescens, Annonidium mannii, Diospyros melocarpa, Staudtia gabonensis, Strombosia grandiflora, Isolonona hexaloba, Garcinia epunctata, Cola griseiflora, Uapaca guineensis et Monodora angolensis.
La grille Fs est principalement caractérisée par les espèces Petersianthus macrocarpus, Trichilia gilgiana, Hannoa klaineana, Entandrophragma angolense, Uapaca guineensis, Funtumia africana,
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 273
Dichostema glauscens, Pycnanthus angolense, Staudtia kamerunensis, Trilepisium madagascariense, Entandrophragma cylindricum, Coelocaryon botryoides et Ricinodendron heudelotii. Dans la grille J,
les espèces fréquemment présentes sont Elaeis guineensis, Pteridium aquilinum, Alchornea
cordifolia, Rauvolfia vomitoria, Manihot esculenta, Triumfetta cordifolia, Caloncoba welwitschii, Costus lucanusianus, Palisota ambigua, Ananas comosus, Aframomum sanguineum, Harungana madagascariensis, et Macaranga monandra. La végétation de la grille L est dominée par Musanga cecropioides, Dichostema glauscens, Petersianthus macrocarpus, Pycnanthus angolensis, Uapaca guineensis, Macaranga spinosa, Trichilia gilgiana, Hannoa klaineana et Coelocaryon botroides (Fig.
2).
Fig. 2. Habitats explorés dans la Réserve Forestière de Masako (Kisangani, RDC) : Forêt à Gilbertiodendron dewevrei (1) ; Forêt secondaire (2) ; Jachère (3) ; Zone de lisière jachère (à gauche) et forêt secondaire (à droite) (4). Photo L. Iyongo, 2010.
Les captures se faisaient durant cinq jours chaque mois [3, 12]. Les relevés ont eu lieu chaque jour autour de 9h00. Les individus ont été identifiés sur la base d’une combinaison de données morphométriques externes (tous les spécimens) [3, 14-16] mesurées à l’aide d’un pied à coulisse et d’analyses phylogénétiques des séquences du cytochrome de l’ADN mitochondrial obtenues (tous les individus des genres Praomys, Hylomyscus, Graphiurus, Grammomys et Mastomys ainsi que quelques représentants des autres genres) [17], qui ont été comparées avec la base de données des séquences d’ADN disponible au Département des Vertébrés de l’Institut Royal des Sciences Naturelles de Bruxelles. Les individus morts lors de la capture (33 spécimens) ont également été utilisés pour des analyses d’ADN et finalement, incorporés comme spécimens dans la collection de l’Institut Royal de Bruxelles et du Laboratoire de Biologie Evolutive (Evolutionary Biology Group) de l’Université d’Anvers.
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 274
Les données récoltées ont permis de déterminer les richesses spécifiques des habitats en se servant de l’indice de Margalef (RMg) dont les valeurs s’obtiennent par la formule ;
) ln( 1 N S RMg
où N est le nombre d’individus et S est le nombre d’espèces capturées. La diversité (D) et l’équitabilité (E) des Rongeurs dans les habitats ont ensuite été déterminées en utilisant les indices de Simpson :
S i ip
D
1 2 et S D E (1/ )où D est l’indice de diversité et pi la proportion d’individus de l’espèce i. Les similarités de la
composition faunistique des habitats ont également été calculées à l’aide du coefficient de Jaccard (Cj) : c b a a Cj
où a est le nombre d’espèces communes pour les habitats comparés ; b et c équivalent au nombre d’espèces présentes seulement dans un des deux habitats [18]. Finalement, les densités relatives (T) ont été déterminées : 100 ) ( (%) t n N T
avec T la densité relative ou indice de piégeage, n le nombre de pièges utilisées et t le nombre de nuits de capture effectives [9].
Pour tester la distribution des abondances, les moyennes mensuelles d’individus capturés dans les différents habitats ont été comparées à l’aide du test t de Student. Pour vérifier l’existence d’un effet « saison », les effectifs observés durant les différentes saisons (septembre à novembre ; décembre à février ; mars à mai ; juin à août) ont été comparés aux valeurs attendues pour une distribution uniforme à l’aide du test de χ². De même, χ² a permis de tester les effets anthropiques sur l’abondance de rongeurs en comparant les effectifs de la période (de juin à août) d’intenses activités anthropiques (incinération des champs et récolte des chenilles) aux valeurs mensuelles attendues selon une distribution uniforme. L’analyse de l’impact de l’anthropisation a été complétée par une comparaison des données de la forêt primaire à Gilbertiodendron dewevrei avec celles de la jachère et de la forêt secondaire. Enfin, pour vérifier l’existence d’un effet de lisière, celle-ci a été comparée aux deux habitats qu’elle sépare (jachère et forêt secondaire).
Résultats
Durant une année de capture, un total de 457 (35.8%), 286 (22.4%), 477 (37.4%) et 55 (4.3%) individus regroupés en 24 espèces et 5 familles a été réalisé respectivement dans les grilles J, L, Fs et
Fppour 7200 nuits-pièges/grille (Appendix 1).
Les Nesomyidés ont été les plus représentés (17 espèces), suivis des Sciuridés (3 espèces), Gliridés (2 espèces) ; les Thryonomyidés et Anomaluridés comptent 1 seule espèce (Appendix 2).
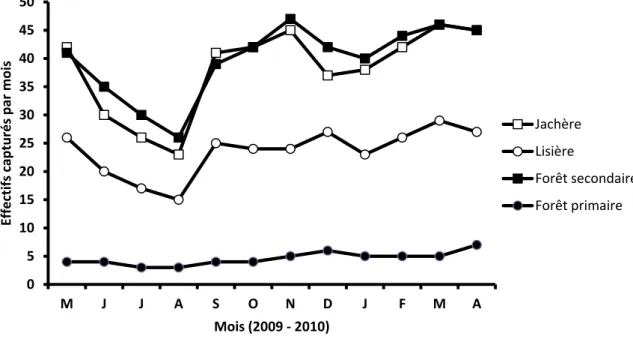
L’évolution annuelle des captures est donnée dans la Fig. 3.
La comparaison des effectifs observés et ceux attendus par saison dans les quatre habitats donne des différences significatives pour la jachère (χ²=15.67 ; p<0.01 ; ddl=3), la lisière (χ²=8.00 ; p<0.05 ; ddl=3) et la forêt secondaire (χ²=9.08 ; p<0.05 ; ddl=3). Par contre, ces effectifs ne diffèrent pas
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 275
significativement en forêt primaire (χ²=1.80 ; p>0.05 ; ddl=3). De même, les effectifs obtenus durant les mois de brûlis des champs et récolte des chenilles (juin à août) comparés aux valeurs mensuelles attendues donnent des différences significatives pour la jachère (χ²= 11.52 ; p<0.01 ; ddl=2), la lisière (χ²=6.63 ; p<0.05 ; ddl=2) et la forêt secondaire ; (χ²=7.72 ; p<0.05 ; ddl=2). A l’inverse, ces effectifs ne diffèrent pas en forêt primaire (χ²= 1.17 ; p>0.05 ; ddl=2).
Les moyennes mensuelles des captures sont très significativement différentes entre la jachère et la lisière (t=5.72 ; p<0.001 ; ddl=22), la jachère et la forêt primaire (t=14.89 ; p<0.001 ; ddl=22), la lisière et la forêt secondaire (t=-7.27 ; p<0.001; ddl=22), la lisière et la forêt primaire (t=14.99 ; p<0.001 ; ddl=22), la forêt secondaire et la forêt primaire (t=18.61 ; p<0.001 ; ddl=22). Par contre, ces moyennes ne montrent pas de différence significative entre la jachère et la forêt secondaire (t=-0.58 ; p>0.05 ; ddl=22).
Fig. 3. Evolution mensuelle des captures (durant 12 mois) dans les 4 habitats étudiés de la Réserve Forestière de Masako (Kisangani, République Démocratique du Congo) à l’aide de 4 grilles de capture (1 ha/grille ; 120 pièges/grille).
La zone de lisière est l’habitat le plus riche (grande valeur de RMg) et le plus diversifié (faible valeur de
D), alors que la forêt primaire à Gilbertiodendron dewevrei est moins riche (petite valeur de RMg) et
moins diversifiée (grande valeur de D) en espèces de Rongeurs. La jachère et la forêt secondaire présentent des richesses similaires. Les équitabilités sont relativement faibles dans les quatre habitats avec la plus grande valeur enregistrée en forêt primaire et la plus faible valeur en forêt secondaire. La densité relative la plus élevée est obtenue en forêt secondaire et la plus faible en forêt primaire ; des valeurs intermédiaires caractérisent la lisière et la jachère (Tableau 1).
Sur la base de Cj, on observe que la forêt primaire montre une similarité de 58% avec la forêt secondaire et de 36% avec la jachère. La zone de lisière partage 77% de similarité avec la forêt secondaire et 70% avec la jachère. La jachère et la forêt secondaire sont similaires à 58%.
0 5 10 15 20 25 30 35 40 45 50 M J J A S O N D J F M A Effectifs capturés par mois Mois (2009 - 2010) Jachère Lisière Forêt secondaire Forêt primaire
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 276
Discussion
Une augmentation des captures est observée durant les périodes de février à mai et de septembre à novembre dans la jachère, la lisière et la forêt secondaire. La même tendance est observée en décembre mais uniquement dans la lisière. Il s’agit des périodes pluvieuses, principalement caractérisées par une faible intensité des activités champêtres, une forte diminution de la perturbation anthropique [13, 19], et la fructification de la plupart des plantes sauvages de la région [20]. Ceci conduirait à un accroissement de la reproduction des rongeurs [21] étant donné que la majorité des espèces présentes à Masako sont insectivores et frugivores. Elles exploitent mieux la nourriture végétale au moment de la reproduction [3]. Par ailleurs, on note qu’il existe un lien entre les variations saisonnières des abondances relatives des rongeurs et les précipitations qui favorisent la fructification des plantes et l’abondance des insectes [22]. Une forte diminution des captures a été détectée de juin à août dans les trois milieux perturbés (jachère, lisière et forêt secondaire). Nous pouvons attribuer ce constat à la saison (grande saison sèche) et aux intenses perturbations anthropiques [13, 23] liées principalement aux brûlis des champs et à la récolte des chenilles [19, 24] durant cette période comme l’attestent les analyses statistiques. En effet, une forte relation a déjà été constatée entre l’intensité du feu, la saison sèche et l’abondance des Rongeurs dans les écosystèmes forestiers tropicaux [1, 25].
Tableau 1. Richesses (RMg ), diversités (D), équitabilités (E) et densités relatives ou indices de
piégeage (T (%)) des rongeurs récoltés de mai 2009 à avril 2010 dans la Réserve Forestière de Masako (Kisangani, République Démocratique du Congo). N = Nombre d’individus capturés ; S = Nombre d’espèces capturées.
Habitats N S R Mg D E T(%)
Jachère 457 19 2,94 0,20 0,26 6,30
Lisière 286 20 3,36 0,15 0,33 3,90
Forêt secondaire 477 19 2,92 0,24 0,22 6,60
Forêt primaire 55 11 2,49 0,24 0,38 0,80
Les captures dans la forêt non perturbée restent quasi uniformes durant toute l’année (capture inférieure à 7 individus/mois). Ceci est vraisemblablement dû aux conditions macroclimatiques régionales quasi constantes [21] qui maintiennent la flore et la faune à un niveau plus ou moins régulier [14, 15], ainsi qu’à l’absence d’activités agricoles [23] dans cette forêt, comme démontrée par les analyses statistiques.
De manière générale, cette étude montre une différence dans la distribution des abondances et des richesses spécifiques entre les habitats concernés ; la même observation a été faite par d’autres études [25], et s’explique par les différences écologiques et structurelles entre ces milieux, y compris les différences de régime de perturbation [7, 26].
La zone de lisière est le milieu le plus riche et le plus diversifié en Rongeurs. Cette grande diversité est liée au fait que la lisière réunit les conditions écologiques des milieux qu’elle sépare [12, 27]. Par conséquent, elle détient, en plus de ses espèces spécifiques, une grande partie des espèces des habitats adjacents [28-30]. La faible richesse et la faible diversité de la forêt primaire à
Gilbertiodendron dewevrei peuvent être attribuées à son sous-bois ouvert, qui fait d’elle un milieu de
prédilection d’un nombre réduit d’espèces de grande forêt [21, 27].
L’abondance et la densité de Rongeurs diffèrent entre la lisière et les habitats adjacents. Nous estimons que la lisière est une entité qui diffère de façon marquée des habitats qu’elle sépare [31] avec des caractéristiques écologiques, voire des espèces, propres [32]. Les faibles abondances et densités de la lisière, comparativement aux habitats qu’elle sépare, seraient attribuées (i) au taux de
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 277
mortalité plus élevé généralement observé dans les lisières [33], (ii) à une forte présence des prédateurs [34, 35], (iii) à son rôle de corridor propice au déplacement des espèces [36] qui y transitent sans y être strictement inféodées [31, 37], et finalement (iv) à sa nature de barrière écologique [38].
La jachère et la forêt secondaire partagent de fortes ressemblances du point de vue leur richesse spécifique, abondance relative, densité relative et moyenne de captures. Ceci s’explique par le fait que ces deux biotopes sont caractérisés par des richesses et des abondances quasi égales toute l’année à Masako [21] et se montrent favorables à la majorité des Rongeurs de forêt [12, 27]. Les équitabilités faibles obtenues dans ces habitats seraient un signe de l’instabilité de la Réserve de Masako liée aux diverses perturbations anthropiques [3] ; ce qui entrainerait une dominance mais aussi une raréfaction de certaines espèces [27].
Sept espèces ont été recensées dans les quatre biotopes exploités (Praomys cf. jacksoni, Hybomys cf.
lunaris, Deomys ferrugineus, Lophuromys dudui, Hylomyscus stella, Malacomys longipes et Praomys misonnei). Ceci est probablement dû à leur mobilité et à la connectivité entre ces habitats qui sont
supposés héberger des métapopulations [30, 39, 40].
La forêt non perturbée partage une faible similarité avec la jachère, traduisant ainsi des différences écologiques et microclimatiques significatives entre ces deux habitats [32, 41]. Par contre, la forêt à
Gilbertiodendron montre une similarité élevée avec la forêt secondaire, ce qui laisse entrevoir une
évolution vers un stade climacique de ce dernier habitat [42]. L’étude met aussi en évidence la présence d’Anomalurus derbianus en forêt, avec deux individus récoltés. Bien qu’elle passe ses journées dans les creux des arbres [43, 44], cette espèce descend par moment à terre, en quête probable de nourriture.
Thryonomys swinderianus a été capturée dans la jachère. La présence de cette espèce savanicole en
milieu forestier des environs de Kisangani est une preuve de son expansion liée à la déforestation croissante en République Démocratique du Congo. Sa présence dans la Réserve de Masako s’explique aussi bien par la dégradation des forêts de cet écosystème que par la forte présence du couvert graminéen dans les jachères, avec des espèces comme Pennisetum purpureum [45].
En conclusion, cette étude a permis de mettre en évidence les effets des activités humaines sur la distribution des rongeurs à Masako. L’existence d’effets liés aux activités anthropiques et aux saisons sur la distribution des Rongeurs a été démontrée dans les habitats anthropisés mais pas en forêt primaire. Les richesses, les abondances, les densités et les moyennes de capture varient, de manière générale, en fonction des habitats, ce qui témoigne d’un effet des changements d’occupation du sol. Lesgrandes valeurs de similarité observées entre la lisière et les deux habitats qu’elle sépare (jachère et forêt secondaire) traduisent son rôle de zone de transition ou écotone. Les équitabilités observées au sein des habitats sont plutôt faibles, ce qui est un signe de l’instabilité de l’écosystème forestier de Masako, favorisant la dominance de quelques espèces de rongeurs à haute valence écologique [3]. La zone de lisière présente des différences de diversité, d’abondance, de densité et de moyenne de captures comparativement aux habitats qu’elle sépare. Ceci signifie que la dynamique de l’occupation du sol sous la pression des activités humaines à Masako induit un réel effet de lisière sur les Rongeurs. La lisière est le milieu le plus riche et le plus diversifié en termes de Rongeurs, à l’inverse de la forêt primaire. En définitive, la plus grande diversité observée dans la lisière ne doit pas être considérée comme un argument en faveur de la création de lisières par fragmentation des forêts, mais plutôt comme une confirmation que la plus grande diversité des espèces au niveau du paysage peut être attendue lorsque le mélange des habitats de lisière et de l’intérieur est optimal [23].
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 278
Implications pour la conservation
La présente étude met en évidence l’influence de l’anthropisation sur la diversité des Rongeurs. Etant donné que chaque système écologique est caractérisé par une interdépendance entre sa configuration, sa composition et son fonctionnement [46], les changements d’occupation du sol à Masako ont induit des effets clairs sur le fonctionnement des unités paysagères et leur composition en Rongeurs. La richesse, la diversité et les abondances de rongeurs varient en général en fonction des biotopes.
Etant donné que certaines espèces sont inféodées au cœur de la forêt, évitant ainsi toute influence de la lisière [29], l’augmentation de la proportion de lisière suite aux activités humaines reste un élément à considérer comme très négatif dans la conservation de la diversité des Rongeurs de forêt, et ce d’autant plus que certaines espèces de Masako manifestent moins de préférence pour cet habitat [12, 24, 27]. Les faibles effectifs de ces espèces dans la lisière en restent une preuve incontestable. La présence de Mastomys natalensis, un rat anthropophile [21, 47] dans le milieu naturel de Masako témoigne de cette forte anthropisation dont souffre cet écosystème. Il y a lieu, partant de cette étude, de songer à surveiller l’évolution des populations de cette espèce et d’établir une gestion responsable de l’écosystème forestier de Masako.
Anomalurus derbianus, capturée puis relâchée dans le contexte de cette étude, figure sur la liste
rouge de l'UICN à cause de données biologiques insuffisantes [43]. Son étude et sa protection passe donc obligatoirement par l’enrayement de la dégradation de l’habitat forestier de Masako.
La fragmentation des forêts congolaises a conduit à l’expansion de Thryonomys swinderianus, une des espèces nuisibles pour l’agriculture, causant de sévères dégâts aux récoltes. En général, ces espèces sont caractérisées par des taux de reproduction et des capacités de dispersion très élevés qui contribuent grandement à leurs impacts négatifs [48]. Il est donc indispensable de surveiller l’augmentation des populations de cette espèce qui semble déjà être bien installée dans les milieux naturels des environs de Kisangani.
Remerciements
Nous remercions la Coopération Technique Belge (C.T.B.) pour la bourse de doctorat accordée à Mr. Léon IYONGO ; la Wallonie Bruxelles Internationale (WBI) pour la subvention complémentaire ; l’Institut Royal des Sciences Naturelles de Bruxelles pour l’identification des échantillons, et la Coordination Provinciale de l’Environnement en Province Orientale (RD Congo) pour avoir autorisé la réalisation de la présente recherche au sein de la Réserve Forestière de Masako.
Références Bibliographiques
[1] Van der Werf, G.R., Randerson, J.T., Giglio, L., Gobron, N. and Dolman, A.J. 2008. Climate controls on the variability of fires in the tropics and subtropics. Global Biogeochemical Cycles 22: 1-13 [2] Mate, M. 2001. Croissance, phytomasse et minéralomasse des haies des légumineuses
améliorantes en cultures en allées à Kisangani (R.D. Congo). Thèse de doctorat. Université Libre
de Bruxelles, Belgique.
[3] Katuala, G.B. 2009. Biodiversité et biogéographie des rongeurs Myomorphes et Sciuromorphes
(Rodentia: Mammalia) de quelques blocs forestiers de la région de Kisangani (R.D. Congo). Thèse
de doctorat. Université de Kisangani, RDC.
[4] Querouil, S. 2001. Intérêts et limites de l’approche moléculaire pour aborder la biographie et la
speciation: l’exemple de quelques mammifères d’Afrique tropicale. Thèse de doctorat. Université
de Rennes, FRANCE.
[5] Fagan, W. F., Fortin, M. J. and Soykan, C. 2003. Integrating edge detection and dynamic modeling in quantitative analyses of ecological boundaries. Bioscience 53: 730-738.
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 279
[6] Harper, K. A., Macdonald, S. E., Burton, P. J., Chen, J. Q., Brosofske, K. D., Saunders, S. C., Euskirchen, E. S., Roberts, D., Jaiteh, M. S. and Esseen, P. A. 2005. Edge influence on forest structure and composition in fragmented landscapes. Conservation Biology 19: 768-782.
[7] Gascon, C., Malcolm, J.R., Patton, J.L., Da Sylva, M.N.F., Bogart, J.P., Lougheed, S.C., Peres, C.A., Neckel, S. and Boag, P.T. 2000. Riverine barriers and the geographic distribution of Amazonian species. PNAS 97: 13672-13677
[8] Tarrant, K.A., Johnson, I.P., Flowerdew, J.R. and Greig-Smith, P.W. 1990. Effects of pesticide applications on small mammals in arable fields and the recovery of their populations. Thornton
Heath 1: 173-182.
[9] Nicolas, V., Barriere, P. and Colyn, M. 2003. Impact of removal pitfall trapping on the community of shrews (Mammalia: Soricidae) in two African tropical forest sites. Mammalia 67: 133-138. [10] Duivenvoorden, J.F., Svenning, J.C. and Wright, S.J. 2002. Beta diversity in Tropical Forests.
Science 295: 636-637.
[11] Makundi, R.H., Massawe, A.W. and Mulungu, L.S. 2005. Rodent population fluctuations in three ecologically distinct locations in north-east, central and south-west Tanzania. Belgian journal of
zoology 135 (Supplement): 159-165.
[12] Iyongo, W.M., Visser, M., Verheyen, E., Leirs, H., Dudu, A., Ulyel, A. et Bogaert, J. 2009. Etude préliminaire des effets de lisière sur quatre espèces de rongeurs dans la Réserve Forestière de Masako (Kisangani, RD Congo). Annales des Instituts Supérieurs d’Etudes Agronomiques 4 : 156-167.
[13] Bogaert, J., Barima, Y.S.S., Iyongo, W.M.L., Bamba, I., Mama, A., Toyi, M. and Lafortezza, R. 2010. Forest fragmentation: causes, ecological impacts and implications for landscape management. In:
Landscape Ecology in Forest Management and Conservation. Li, C., Lafortezza, R. and Chen, J.
(Eds.), pp.273-296. Springer book, New York.
[14] Amundala, D., Bapeamoni, A., Iyongo, W., Kennis, J., Gambalemoke, M., Kadange, N., Katuala, P.G.B. and Dudu, A. 2005. The population structure of four rodent species from a tropical region (Kisangani, D.R. Congo). Belgian Journal of Zoology 135 (supplément): 127-131.
[15] Katuala, G.B., Hart, J.A., Hutterer, R., Leirs, H. and Dudu, A. 2005. Biodiversity and ecology of small mammals (Rodents and Shrews) of the “Réserve de Faune à Okapi”, Democratic Republic of the Congo. Belgian Journal of Zoology 135 (supplement): 191-196.
[16] Mukinzi, I., Katuala, G.B., Kennis, J., Gambalemoke, M., Kadange, N., Dudu, A.M., Colyn, M. and Hutterer, R. 2005. Preliminary data on the biodiversity of rodents and insectivores (Mammalia) in the periphery of Kisangani (D.R. Congo). Belgian Journal of Zoology 135 (supplément): 133-140. [17] Terryn, L., Wendelen, W., Leirs, H., Lenglet, G. and Verheyen, E. 2007. African Rodentia. (Web
address: http://projects.biodiversity.be/africanrodentia).
[18] Magurran, A.E. 2004. Measuring biological diversity. Blackwell Publishing, UK, Oxford.
[19] Juakaly, M.J.L. 2007. Résilience et écologie des araignées du sol d’une forêt équatoriale de basse
altitude (Réserve Forestière de Masako, Kisangani, RD Congo). Thèse de doctorat. Université de
Kisangani, RD Congo.
[20] Dechuvi, N. 2010. Etude socio-économique et chimique des fruits alimentaires spontanés de la
région de Bengamisa. Mémoire de DES. Université de Kisangani, RD Congo.
[21] Dudu, A.M. 1991. Etude du peuplement d’insectivores et de rongeurs de la forêt ombrophile de
basse altitude du Zaïre (Kisangani, Masako). Thèse de doctorat. Antwerpen, Belgique.
[22] Nicolas, V. and Colyn, M. 2003. Seasonal variations in population and community structure of small rodents in a tropical forest of Gabon. Canadian Journal of Zoology 6: 1034-1046.
[23] Vranken, I., Djibu, K.J.P., Munyemba, K.F., Mama, A., Iyongo, W.M.L., Bamba, I., Laghmouch, M. and Bogaert, J. 2011. Ecological impact of habitat loss on African landscapes and biodiversity. In:
Advances in Environmental Research. Daniels, J.A. (Eds.), pp 365-388. Nova Science Publishers,
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 280
[24] Iyongo, W.M., Visser, M., Verheyen, E., Leirs, H., Iyongo, B., Ulyel, A. et Bogaert, J. 2009. Etude préliminaire des effets de la fragmentation des forêts sur la similarité des habitats et leurs richesses en espèces de rongeurs (Masako, RD Congo). Annales des Instituts Supérieurs d’Etudes
Agronomiques 4 : 177-186.
[25] Decher, J. and Bahian, L.K. 1999. Diversity and structure of terrestrial small mammal communities in different vegetation types on the Accra Plains of Ghana. Journal of Zoology 247: 395-408.
[26] Makundi, R.H., Massawe, A.W., Mulungu, L.S. and Katakweba, A. 2009. Species diversity and population dynamics of rodent in a farm-fallow field mosaic system in Central Tanzania. African
Journal of Ecology 48: 313-320.
[27] Iyongo, W.M. 2008. Etude des effets de lisière sur les populations des rongeurs dans la Réserve
Forestière de Masako (Kisangani, R.D. Congo). Mémoire DEA. Université Libre de Bruxelles,
Belgique.
[28] Anderson, C.S., Cady, A.B. and Meikle, D. 2003. Effects of vegetation structure and edge habitat on the density and distribution of white-footed mice (Peromyscus leucopus) in small and large forest patches. Canadian Journal of Zoology 81: 897-904.
[29] Mc Donald, R.I. and Urban, D.L. 2006. Edge effects on species composition and exotic species abundance in the North Carolina Piedmont. Biology Invasions 8: 1049-1063.
[30] Alignier, A. 2010. Distribution des communautés végétales sous l’influence des lisières forestières
dans des bois fragmentés. Thèse de doctorat. Université de Toulouse, France.
[31] Cadenasso, M. L., Pickett, S. T. A., Weathers, K. C. and Jones, C. G. 2003. A framework for a theory of ecological boundaries. Bioscience 53: 750-758.
[32] Imbeau, L., Drapeau, P. and Mokkonen, M. 2003. Are forest birds categorised as "edge species" strictly associated with edges? Ecography 26: 514-520.
[33] Ewers, R. M. & Didham, R. K. 2006. Confounding factors in the detection of species responses to habitat fragmentation. Biological Reviews 81: 117-142.
[34] Arrignon, F. 2006. Un modèle multi-agent pour simuler la dynamique hivernale d'un insecte
auxiliaire des cultures (Episyrphus balteatus, Diptera: Syrphidae) dans un paysage hétérogène.
Université Paul Sabatier, Toulouse. France.
[35] Le Tacon, F., Selosse, M. A. et Gosselin, F. 2001. Biodiversité, fonctionnement des écosystèmes et gestion forestière. Deuxième partie: interventions sylvicoles et biodiversité. Revue Forestière
Française 53 : 55-80.
[36] Ries, L. and Fagan, W. F. 2003. Habitat edges as a potential ecological trap for an insect predator.
Ecological Entomology 28: 567-572.
[37] Ries, L. and Sisk, T. D. 2010. What is an edge species? The implications of sensitivity to habitat edges. Oikos 1636-1642.
[38] Cadenasso, M. L. and Pickett, S. T. A. 2001. Effect of edge structure on the flux of species into forest interiors. Conservation Biology 15: 91-97.
[39] Kindlemann, P., Aviron, S. and Burel, F. 2005. When is landscape matrix important for determining animal fluxes between resource patches? Ecological Complexity 2: 150-158
[40] Kindleman, P. and Burel, F. 2008. Connectivity measures: a review. Landscape Ecology 23: 879-890.
[41] Yarrow, M. M. & Marin, V. H. 2007. Toward conceptual cohesiveness: A historical analysis of the theory and utility of ecological boundaries and transition zones. Ecosystems 10: 462-476.
[42] Bangirinama, F. 2010. Processus de la restauration écosystémique au cours de la dynamique
post-culturale au Burundi: mécanismes, caractérisation et séries écologiques. Thèse de doctorat.
Université Libre de Bruxelles, Belgique.
[43] Corbin, C.E. and Cordeiro, N.J. 2006. Gliding characteristics of Lord Derby’s Anomalure (Anomalurus derbianus) in Tanzania. African Journal of Ecology 44: 106-108.
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 281
[44] Schunke, A.C. and Hutterer, R. 2005. Geographic variation in the West African scaly-tailed squirrel Anomalurus pelii (Schlegel and Müller, 1845) and description of a new subspecies (Rodentia: Anomaluridae). In: African Biodiversity. Huber, B.A. et al. (Eds.), pp.321-328. Springer, Netherlands.
[45] Jori, F. 2001. La production de rongeurs en milieu tropical. Bois et Forêts des Tropiques 269(3) : 31-42.
[46] Bogaert, J. et Mahamane, A. 2005. Ecologie du paysage: cibler la configuration et l’échelle spatiale. Annales des Sciences Agronomiques du Bénin 7 : 39 - 68
[47] Fichet-Calvet, E., Koulémou, K., Koivogui, L., Soropogui, B., Sylla, O., Lecompte, E., Daffis, S., Kouadio, A., Kouassi, S., Akoua-Koffi, C., Denys, C. and Meulen, J. 2005. Spatial distribution of commensal rodents in regions with high and low Lassa fever prevalence in Guinea. Belgian
Journal of Zoology 135(Supplement): 63-67.
[48] Brooks, T.M., Mittermeier, R.A., Mittermeier, C.G., Da Fonseca, G.A.B., Rylands, A.B., Konstant, W.R., Flick, P., Pilgrim, J., Oldfield, S., Magin, G. and Hilton-Taylor, C. 2002. Habitat loss and extinction in the hotspot of biodiversity. Conservation Biology 16: 909-923.
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 282
A B
C D
E F
Appendix 1. Quelques espèces capturées vivantes (durant 12 mois) dans la Réserve Forestière de Masako (Kisangani, République Démocratique du Congo) : Anomalurus derbianus (A), Hylomyscus spp (B), Praomys spp (C et D), Mastomys natalensis (E) et Lemniscomys striatus (F) (Photo L. Iyongo)
Tropical Conservation Science | ISSN 1940-0829 | Tropicalconservationscience.org 283
Appendix 2. Espèces capturées (durant 12 mois) dans la Réserve Forestière de Masako (Kisangani, République Démocratique du Congo).
Effectifs dans les habitats
Familles Espèces Jachère Lisière Forêt
secondaire
Forêt Primaire
Nesomyidae Praomys cf. jacksoni DE WINTON, 1897 153 67 192 22
Hybomys cf. lunaris THOMAS, 1906 101 54 99 14
Deomys ferrugineus THOMAS, 1888 28 29 74 4
Lophuromys dudui VERHEYEN et al., 2002 79 50 30 2
Hylomyscus stella THOMAS, 1911 30 18 17 3
Stochomys longicaudatus TULLBERG, 1893 26 27 25 0
Hylomyscus aeta THOMAS, 1911 0 4 8 1
Malacomys longipes MILNE-EDWARDS, 1877 2 12 13 4
Oenomys hypoxanthus PUCHERAN, 1855 6 3 1 0
Lemniscomys striatus LINNAEUS, 1758 7 2 0 0
Nannomys cf. grata THOMAS & WROUGHTON, 1910 7 4 4 0
Praomys misonnei VAN DER STRAETEN & DIETERLEN, 1987 3 1 2 1
Praomys mutoni VAN DER STRAETEN & DUDU, 1990 1 2 4 0
Grammomys kuru THOMAS & WROUGHTON, 1907 2 1 1 0
Hylomyscus parvus BROSSET, DUBOST & HEIM DE BALSAC, 1965
Cricetomys emini WROUGHTON, 1910
2 0 0 1 1 2 1 2
Mastomys natalensis SMITH, 1834 2 0 0 0
Gliridae Graphiurus lorraineus DOLLMAN, 1910 0 6 1 0
Graphiurus surdus DOLLMAN, 1912 4 2 1 0
Sciuridae Funisciurus anerythrus THOMAS, 1890 1 1 0 0
Funisciurus pyrropus CUVIER, 1833 1 1 0 0
Paraxerus boehmi REICHENOW, 1886 0 1 1 0
Thryonomyidae Thryonomys swinderianus TEMMINCK, 1827 2 0 0 0
Anomaluridae Anomalurus derbianus GRAY, 1842 0 0 1 1