Copyright© 1983, AmericanSociety forMicrobiology
Action of Patulin
on
a
Yeast
Z. LEZISUMBU,1 P.THONART,1* AND J. BECHET2
DepartementdeTechnologieAgro-AlimentaireetForestiere,Faculte desSciences Agronomiques, B5800 Gembloux, Belgium,1and Institutde Recherches et Institut des Industries de Fermentation, Ceria, B1070
Anderlecht, Belgium2
Received23 February 1982/Accepted1 October 1982
The action ofpatulinonSaccharomyces cerevisiaewasstudied. At weakdoses, the druginhibitedgrowth, but inhibitionwastransient. After 10
min,
synthesesof rRNA,tRNA,andprobablymRNAwereblocked;thiswasshownbyradioactiveprecursorincorporationassaysandgel electrophoresis ofRNAs. Afterrecovery
ofgrowth, patulin disappearedfrom the medium. It seemedthat thisdegradation resulted fromtheactivityofaninducibleenzymatic system.Induced cells resisted
veryhigh patulin concentrations.
Patulin isamycotoxin produced bynumerous molds, especially Penicillium patulum and
Bys-sochlamysnivea.Thismycotoxinhas been iden-tified in various fruit juices and principally in applejuice andsilage (1, 4, 11, 13). The biosyn-thesis of patulin by Penicilliumurticaehasbeen studied (6),and a greatdealof datahave shown that itmaybe produced inveryabundantyields
(up to50% ofthebiomass increase [14]);
howev-er, nofunction has been discerned for thistoxin. The action spectrum ofpatulin is very broad,
including mostbacteria and animal and vegeta-ble cells. Neither the primary target nor the
mechanism of action of the toxin has been
specified (9, 10, 12). Manyenzymes having the
sulfhydryl group in their active sites are
sensi-tivetopatulin, butenzymes withoutthe
sulfhy-drylgroup canbe equally inhibited(12). Mouley andHatey (10) reported the inhibition of RNA
polymeraseAactivity in isolatedratliver nuclei.
Our purpose in this study was to specify the
action ofpatulinon Saccharomyces cerevisiae. We chose this yeast because of the
well-docu-mented fact that patulin disappears during fer-mentation of apple juice; so far, no model to explain this degradation hasbeen proposed.
MATERIALS AND METHODS
Strain. The S. cerevisiae strain used in this work
(1278b [a]) is a haploid strain and has been described by Grensonetal.(5).
Medium. We used minimal medium 149, which contains 3% glucose and has been previously de-scribed (5).
Incorporation test. Protein synthesis was measured by determining the incorporation of [3H]leucine and
[3H]lysine.
Yeast cells were grown at 29°C in minimal medium with(per milliliter) 0.4 ,uCi of[3H]leucine,0.4p.Ciof[3H]lysine, 100 .gof coldL-leucine, and 100 ,ug of L-lysine.Atintervals,1mlof culture was boiled for 30min in 5 mlof10%trichloroacetic acid; precipitates
werecollectedonHAmembrane filters(0.45-p.mpore size; Millipore Corp.), and radioactivity was
mea-sured.
For determining RNA synthesis, the culture was grown at29°C in minimal medium149;at logphase,
[3H]uracil (0.4 ,uCi/ml; specific activity, 53 Ci/mmol)
or[14C]adenine(0.4p.Ci/ml; specific activity,53
Ci/m-mol)wasadded. Atintervals,theamountof
radioac-tivityincorporated wasdetermined aspreviously de-scribed(16).
Labeling of cells for RNAanalysis,RNAextraction,
anddiscontinuouspolyacrylamide gelelectrophoresis
(2.4 to7%) were performed as described previously
(16).
Patulin.Patulinwasproduced byP.patulumNRRL 2159Aandpurifiedaspreviouslydescribed(14).It was identifiedbyhigh-performanceliquid chromatography
andinfrared spectrum andmelting point studies; com-mercialpatulin (P 1639; SigmaChemical Corp.) was used asthe reference. Patulin wasassayed by
high-performanceliquidchromatography.A Hewlett-Pack-ard model 1084 B chromatograph with a variable-wavelength UV detector, operating at 277 nm, was used. All solventswere ofspectrophotometricgrade. Analysis was carried out with a 10-p.m Lichrosorb (Hibar) silica column (E. Merck AG). The mobile phase was
chloroform-cyclohexane-ethanol-acetoni-trile(25:7.5:1.5:1[vol/vol/vol/vol]),thesamplevolume was 10p.l,the solvent flowrate was2ml/min,and the patulin retention time was 2.91 min. The culture sampleswerecentrifuged, filtered(0.45-p.mHAfilters [Millipore]), and then injected without previous ex-tractionorpurification (17).
RESULTS
Action ofpatulin on RNA synthesis. In the first setof experiments, patulin (0to 100 ,ug/ml)and
[3H]uracil
(0.4 ,uCi/ml) were added simulta-neously to alog-phase culture. At various times,sampleswerewithdrawn as described above. A decrease ofat least 50% in the uracil incorpo-ration rate was observed after 10 min of
20
01
C)0-10
0
I~~
0 ,-IO-- +-+0
10
20
minutes
FIG. 1. Uracilincorporation into I ence ofpatulin at 0 ,ug/ml (0),50
R.
,ug/ml (+).tion in the presence of the dru
p.g/ml)
(Fig. 1).Inthe second set ofexperimer ,ug/ml)wasadded at zero
time,
artive precursor
([14C]adenine;
0.2added after0,4,9,and 17min. T(
ity inthe cells(adenine pool plus ration) and RNA incorporation v
1i
a-i
C)
lular pool indicated that nucleotide uptake was less affected by patulin than was the RNA
incorporation.
Analysis of RNAsynthesized in the presence of patulin. An S. cerevisiae culture grown at29°C
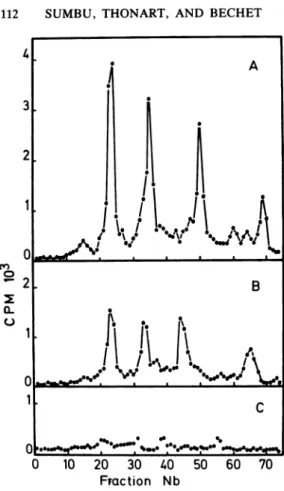
was divided into several portions. Various amounts of patulin (0, 25, 50 ,ug/ml) and a constant amount [3H]uracil(10 ,uCi/25 ml) were added.Cellswereharvestedafter 3 h, and RNAs were extractedand analyzed bypolyacrylamide
gel electrophoresis (16) (Fig. 3). It is clearthat
-_-,o_-
thesyntheses
ofrRNA and tRNAspecies
weretotally inhibited in the presence of 50
jig
of ~+-+ +~ patulin per ml. At 25jig
of patulin per ml,inhibition was not complete, but there was no preferential inhibition ofeither RNA species.
Protein synthesis was slightly affected at 25
30 40 ,jig/ml. At 50
jiglml,
the reduction of the aminoacid incorporation rate was consistent with an
RNA inthepres- mRNA half-life of 30
min
(15).g/ml (0), or 100 Degradationof patulin by S. cerevisiaein vivo. Weadded 50jigofpatulinper ml to alog-phase culture; after a transient inhibition period,
growth and RNA synthesis resumed at a rate Lg (50 and 100 slightly slower than that seenin the absenceof patulin (Fig. 4). During growth, there was an its, patulin (50 important decrease in the patulin concentration
nd the radioac- in the medium(Fig. 5). Thisdecrease, although 5 ,uCi/ml) was not strictlyparallelwithgrowth,wasassociated
)talradioactiv- with it; cells killedby heat(10 minat
800C)
did RNAincorpo- notinduce the reduction of the patulin concen-vere measured tration (Fig. 6). Inhibition ofprotein synthesis,0
2
4
6
8
min
utes
0
2
4
6
8
FIG. 2. Influence ofpatulinon
["4C]adenine
incorporation
as afunction of time. (A)Total radioactivityofcells(adeninepoolplus RNAincorporation);(B) RNA
incorporation
(trichloroaceticacid-precipitablemateri-als). Patulinwasaddedat zerotime,andadeninewasaddedat0min(0),4min(0),9min(+),or17(x)minafter
B
L
Ili*I..J.
17
0~~
ft be.so 0 10 20 30 40 50 60 70 Fraction NbFIG. 3. Influence of patulin at 0 ,g/ml (A), 25
,Lg/ml (B), or 50 ,ug/ml (C) on rRNA and tRNA syntheses. Thetechniqueof discontinuous polyacryl-amidegelelectrophoresis (2.4to7%)andthe
correc-tion procedure for RNAlossesduringextraction have been describedpreviously (16).
effectedbyadding2,ug of cycloheximide perml at the same time that patulin was added,
pre-vented the disappearance of the mycotoxin. Nevertheless, if cycloheximide was added 3 h
after theaddition ofpatulin, the degradationrate wasslowed down but notstopped (Fig. 6).
Effect of patulin concentration. Log-phase cells andcells preincubated for 3 h in the pres-ence of 50,ugofpatulin per mlwere incubated
with 12.5 to 500 ,g of patulin per ml. Patulin
degradation was measured after 6, 12, 18, 24,
and 48 h(Fig. 7). At 12.5, 25, and 50 ,ug/ml, the patulin degradation rate was slightly higher in
preincubated cells;at200 and 500,xg/ml, inacti-vation of patulinwas noteworthy in preincubat-ed cells, but no or nearly no degradation
oc-curred in the reference culture. This was a
consequence of the fact that preincubated cells
wereresistant andgrewin thepresenceofhigher
dosesofpatulin,whereasnon-preincubated cells didnotgrow.
6 5 43
8
25c
04.
4E
3< z 2 1 2 3 4 5 6 7 hoursFIG. 4. Influence of patulin (50 ,ug/ml) on yeast growth (0) and RNA incorporation(0). Patulin was added at 90 min. Yeast growth was measured by optical density determinations (540 nm), and incorpo-ration in RNA wasmeasured asdescribed in the text.
DISCUSSION
Althoughpatulin was discovered in the1940s, its inhibition mechanism is not yet known. Some facts have been demonstrated, but the primary
target of the toxin remains undiscovered. Patulin reacts with sulfhydryl groups (2, 3, 8); thus, many enzymeshavingasulfhydrylgroup at their active sites areinhibited by patulin, but this is not a general rule, and there are sensitive en-zymeswhich do not possess such a group.
Another hypothesis has been proposed by
Mouley and Hatey (10); these authors have shown the inhibition of RNA polymerase A in isolated rat liver nuclei. RNA polymerase B
(responsible formRNA synthesis) is less sensi-tive; nevertheless,thedose ofpatulin used was very high (200,ug/mlfor50% inhibitionofRNA
polymerase A). Using this system, Hatez and
Gaye have also shown inhibition of protein synthesis (7). Inhibition of S. cerevisiae growth by patulin is transient, and theduration is pro-portional to the dose of the toxin (15).
There are thus two stages in the action of patulin on yeast cells: first, the action of the
toxin on yeast metabolism and the subsequent inhibition ofgrowth; second, the resumption of
growth, indicating the appearance of a resist-ance mechanism probably associated with the
previouslyreporteddisappearanceofthe myco-toxinduring apple juice fermentation.
Uracilincorporation into trichloroacetic
acid-insoluble material (ca. 10 min) was rapidly stopped at patulin concentrations of >12.5
,ug/ml. Thisrapidcessation obviously indicated
an arrest ofthe synthesis ofrRNA, the major component of the total RNA (>80%). To draw a
o~~~~~~~~~~~~~~~~0- LUO(-0i 10
06
6
0 -~~~~~~~~~~~~~~~~~~~~~4-LQ2
/
0~10
20
30
hours
FIG. 5. Patulindegradation(0)during growth(0).Patulin(50,ug/ml)wasaddedtoalog-phase cultureat90 minandmeasuredasdescribedinthetext.Growthwasmeasuredbyoptical density determinations(540 nm).
of the resultsof thisincorporationtestwould be hazardous. The problem has been analyzed by polyacrylamide gel electrophoresis and, indi-rectly, by studying the kinetics of radioactive amino acid incorporation. The syntheses of rRNAs(25S, 17S,and5S)and tRNAs(4S)were
equally inhibited (Fig. 3). The kinetics of radio-active amino acid incorporation (into tricloroa-cetic acid-precipitable materials) afterthe
addi-tion of 50 ,ug of patulin per ml were consistent
0 0) 0) 0._ Q Q a h. a L-L C a-e 6 0 12
with an mRNA half-life of 30 min (data not
shown). This behavior is similar to the pattern
observed forthermosensitive S. cerevisiae
mu-tantstrains4572,4472, and 1564 (16) and qualita-tively similartothe behaviorinduced by dauno-mycine and lomofungin, which are known to
inhibit RNA synthesis. However, the modes of action oflomofungin and patulinarenotsimilar: lomofungin is a chelating agent with an action
whichcanbe reversed by Mg2+. Patulin's action
24
36 hours 48FIG. 6. Patulindegradation. Symbols: 0,50 ,ugofpatulinpermladded;0,50,ugofpatulin permladded;2
,ugofcycloheximideperml added3 h afteraddition ofpatulin(arrow);+, 50,ugofpatulin perml and 2,ugof cy-cloheximideperml addedat thesametime; x,50,ugofpatulinpermladded; cells killedbyheat (10minat80°C).
0 S~~~~~~
30~~~~~~
0~~~~~~~~~ 0-1-
+ 4 20i o- 11.0 E o 00
-0.j.6.
0Q2I
0 24 480 24 48 0 24 48 hoursFIG. 7. Patulindisappearance.Symbols: 0,cellspreincubatedfor 3 h with 50 ,gof
patulin
perml;
*, non-preincubated cells withpatulinat50,ug/ml(A),100,ug/ml
(B),or500,g/ml
(C).is not reversed by 20 ,ug ofMg or Mn per ml
(data not shown). These arguments strongly favor the hypothesis that the
primary
target ofpatulin is the
transcription
mechanism.Disappearance of
patulin
during
fermentation isawell-known phenomenon: neithera mecha-nism for it nor a lagperiod
preceding
growth resumption has everbeenreported. Our results show thatpatulindegradation
isanactive proc-ess and a consequence of yeast metabolism: pasteurized yeastswereincapable
ofprovoking
the
disappearance
of the toxin(Fig. 6).
Theinducible character of the
phenomenon
wasfur-therprovenbythe resultsobtained for
patulin
in the presence ofcycloheximide,
anantifungal
agent which blocks peptide bond formation.When patulin and cycloheximide were added
simultaneously
to a culture, protein synthesiswasblocked andno toxinwas degraded(Fig.6).
These results indicate that the wild-type strain was incapable of spontaneous patulin inactiva-tion and that detoxicainactiva-tion requires protein
syn-thesis.
When cycloheximide wasadded 3 h after the addition ofpatulin, degradationcontinued but at aslower rate(Fig.6),possiblyindicatingthatthe
newly synthetized proteinshavea catalytic rath-erthan
stoichiometric
role.At patulin concentrations of >200
,ug/ml,
growth was definitely blocked and resistance inductionwasimpossible.This could be a conse-quence of complete inhibition of proteinsynthe-sisby patulin athigh concentrations. However,
ifthe cells are preinduced for 3 h at 50
,ug/ml,
they become resistant todoses of >200
jxg/ml.
Patulin-sensitivemutants in which patulin
degra-dation isnolonger inducible havebeenisolated (P. Thonart and J. Bechet, Veme Int. Symp.
Yeast 1980, p. 225-230). The present
experi-ments did not enable us to determine if the induced protein(s) acts directly on patulin or permits the synthesis of a detoxifying sub-stance(s). This question is beingstudied.
ACKNOWLEDGMENTS
We thank M.GregoireandJ.Maquet for excellenttechnical assistanceand A. Pierard and F.Hilgerforvaluable discus-sionsandforreadingthemanuscript.
This workwassupported byNorth AtlanticTreaty Organi-zation research grantno.1139,bythe Fonds delaRecherche FondamentaleCollective,andbyCANIRA.
LITERATURECITED
1. Bechet, J.,and P.Thonart.1978. Lesprincipales myco-toxinesetleurimportance dans lesproduits alimentaires, p. 75-87. In Serie syntheses bibliographiques, no. 20. Apria, Paris.
2. Brachett, R. E., and E. H. Marth. 1979. Ascorbic acid and ascorbate cause disappearance of patulin from buffer solutions andapple juice. J. Food Prot. 42:864-866. 3. Ciegler, A., A.C. Bechwith, andL. K. Jackson. 1976.
Teratogenicity of patulin adducts formed with cysteine. Appl. Environ.Microbiol. 31:664-667.
4. Escoula, L. 1974. Moisissures toxigenes des fourrages ensiles. I. Presencedepatulinedans les frontsdecoupe. Ann. Rech. Vet.5:423-432.
5.Grenson, M., M.Mousset, J.M.Wiame, and J. Bechet. 1966.Multiplicityoftheamino acid permease in Saccha-romyces cerevisiae.Biochim. Biophys. Acta 127:325-338. 6. Grootwassink, J.W.D., and G. M. Gaucher. 1980. De novo biosynthesis of secondary metabolism enzymes in homogeneous cultures of Penicillium urticae. J. Bacteriol. 141:443-455.
7. Hatez, F.,andP.Gaye. 1978. Inhibition oftranslation in reticulocyte by the mycotoxin patulin. FEBS Lett. 95:252-256.
8. Lindroth, S.,and A.VonWright. 1978.Comparison ofthe toxicities ofpatulin and patulin adducts formed with cysteine. Appl. Environ. Microbiol. 35:1003-1007. 9. Mayer, W.M., andM.S. Legator. 1969. Production of
petitemutantsofSaccharomycescerevisiaeby patulin.J. Agric.FoodChem.17:454-456.
10. Mouley, G., andF.Hatey. 1977. Mechanism of the in vitro inhibition oftranscriptionby patulin, amycotoxin from
toxin ofpotential concern in foods. J. MilkFoodTechnol. 38:695-705.
14. Sumbu,Z.L., P. Thonart, J.P.Wathelet, P.Blanpain, andJ.Bechet.1981. Lapatuline.Production, purification, degradation etdosage par CPG. Rev. Ferment. Ind. Alim. 36:122-129.
Thermosensitive mutations affecting ribonucleic acid polymerases in Saccharomyces cerevisiae. J. Bacteriol. 125:25-32.
17. Ware, G., C.W.Thorpe, and A. E. Pohland. 1974.Liquid chromatography method for determination of patulin in apple juice.J. Assoc. Anal.Chem. 57:1111-1113.