Serotonin type 4 receptor dimers
Texte intégral
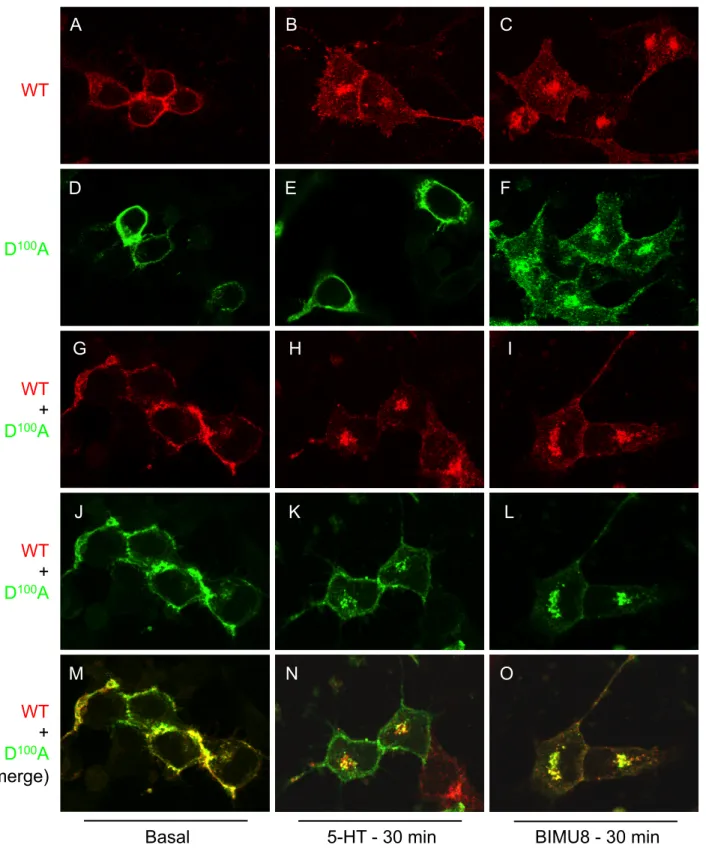
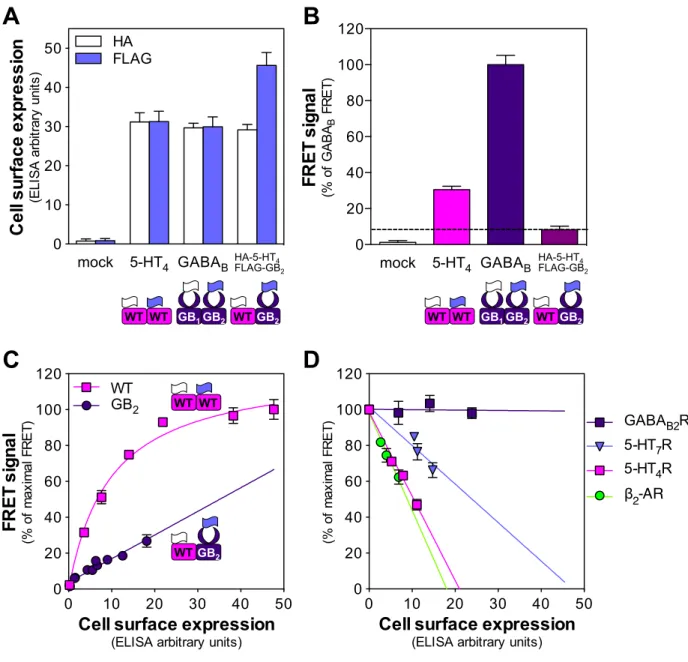
Figure
Documents relatifs
For instance, UNC EFC’s report from 2017 details a case where the City of Phoenix reported spending over $600,000 in advisor fees “for assistance carrying out highly
However, if a training procedure can be created that improves the alignment of the actual motions experienced with the desired motion, even when motor errors are large, such a
subtracted signal. The standard deviation formula is shown below for sample-size K and values xi,.. It is clear that better statistics are possible with an increase
Moreover, it has been shown within the layered superconductor model under the applied in-plane magnetic field in the FFLO phase that the experimentally observed
Here, we took advantage of lysolcithin- induced demyelination to investigate the expression dynamics of the oligodendrocyte specific connexin 47 (Cx47) and to some extent that
one or two of the labile monodentate ligands can be substituted, we focused our attention on the influence of the alkyl‐substituted pyridine ligand on magnetic and
Abstract Approximately half of all newborns with con- genital heart disease are asymptomatic in the first few days of life. Early detection of ductal-dependant cardiac mal-
