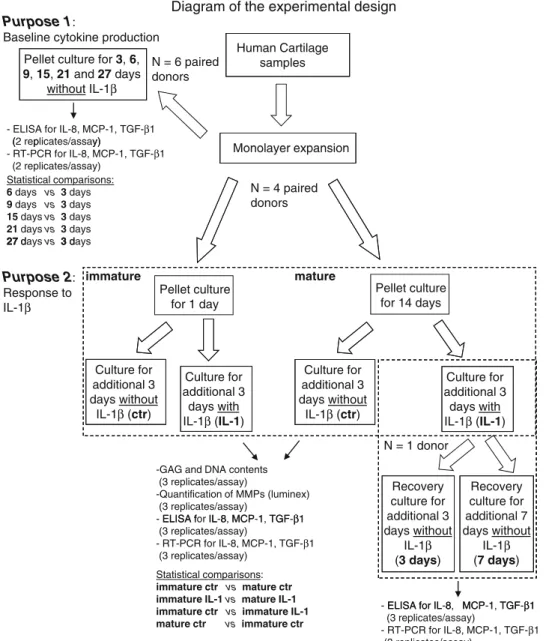
Engineered Cartilage Maturation Regulates Cytokine Production and Interleukin-1β Response
12
0
0
Texte intégral
Figure


Documents relatifs