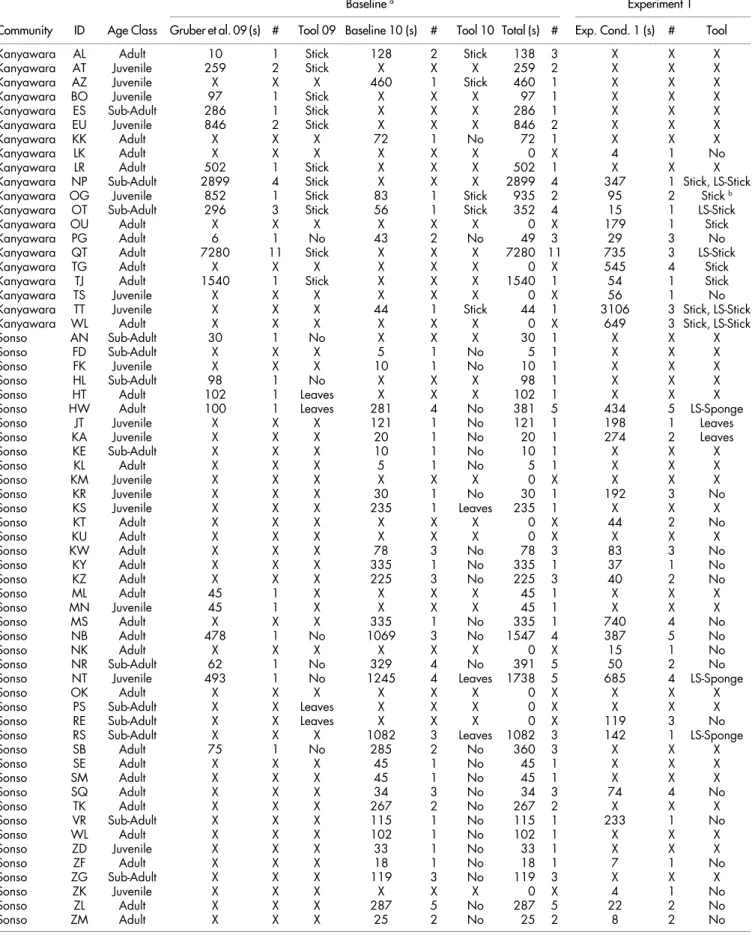
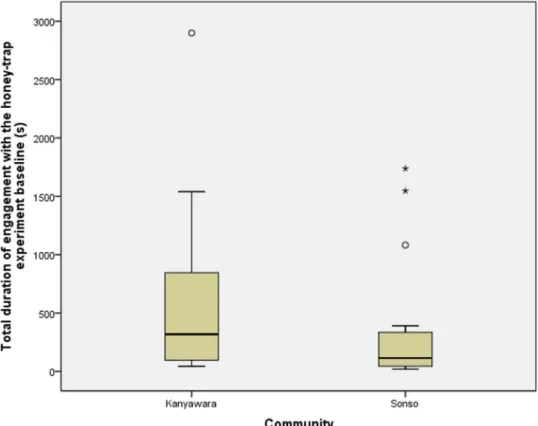
Community-specific evaluation of tool affordances in wild chimpanzees
Texte intégral
Figure

Documents relatifs
By considering Brochard–de Gennes slip model and molecular chain dynamics at the wall, we propose an expression for the critical shear stress at the onset of stick–slip as a
Effect of epidermal growth factor receptor tyrosine kinase domain mutations on the outcome of patients with non-small cell lung cancer treated with epidermal growth factor
The degree of abstraction is considerably reduced in the sense that there could be no one model of the market process but plenty of models of market processes according to the
In the second part of the article Comme Appel´e du N´eant – As If Summoned from the Void: The life of Alexandre Grothendieck by Allyn Jackson, pub- lished in the Notices of the
Regardless of the early period, current studies are sufficient for the comparisons of experimental uncertainties between the low-pile-up measurement and the W-mass measurement at 7
In this case, the volume and weight packing factors are assumed to be 1.0 because 100 percent of the volume in the tank can be used and the tank weight is included as a
Les pharmaciens de ce groupe ont également reçu une formation sur les soins pharmaceutiques, mais plus générale que celle du groupe « soins pharmaceutiques », et ils n’avaient
La Revue Francophone de Recherche en Ergothérapie est publiée par CARAFE, la Com- munauté pour l’Avancement de la Recherche Appliquée Francophone en
