HAL Id: hal-02796822
https://hal.inrae.fr/hal-02796822
Submitted on 5 Jun 2020HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Effet de la fertilisation NPK sur la production et les
traits d’une prairie permanente fauchée
Paul Moreau
To cite this version:
Paul Moreau. Effet de la fertilisation NPK sur la production et les traits d’une prairie permanente fauchée. Biodiversité et Ecologie. 2016. �hal-02796822�
Rapport de stage effectué du 4 avril au 4 juin 2016 au sein de
l’INRA, Unité de Recherches sur l’Ecosystème Prairial (UREP)
Soutenance le 13 juin 2016.
Effet de la fertilisation NPK sur la production et les traits
d’une prairie permanente fauchée
Moreau Paul
Université de Rennes 1
Master 1 Biodiversité-Ecologie-Environnement
Mention Patrimoine Naturel et Biodiversité 2015-2016
Photo : Paul Moreau, Theix.
Encadrement
RESPONSABLE DE STAGE :
MME NATHALIE VASSAL (MAITRE DE CONFERENCE) MME FREDERIQUE LOUAULT (INGENIEURE DE RECHERCHE) MME CATHERINE PICON-COCHARD (CHARGEE DE RECHERCHE)
TUTEUR UNIVERSITAIRE :
Dans un premier temps, je tiens à remercier Pascal Carrère, ingénieur de recherche et directeur de l’UREP, pour m'avoir accueilli dans son unité de recherche
Pour m’avoir encadré, l’accueil, le temps accordé, les conseils, les nombreuses relectures de mon rapport et pour leur gentillesse, un énorme merci pour mes maîtres de stage, Nathalie Vassal (VetAgroSuP), Frédérique Louault (UREP) et Catherine Picon-Cochard (UREP).
A tous ceux avec qui j’ai été amené à travailler, pour les moments partagé autour d’un repas, d’une pause-café, pour des conseils, en statistiques notamment, des services et j’en passe, titulaire comme non titulaire, Nedjla, Geoffrey, Elisa, Camille, Iris, Pauline, Pierre, Ludo, David x 2, Alexandre, Éric, Vincent, Laurine, Jean-Noël, Olivier, Gaël, Julien, Sandrine, Raphael, Gianni, Annick, Laurence, Sylvie…Merci !
Merci à Michel Frain, Botaniste, pour m’avoir appris quelques rudiments de détermination des graminées !
Enfin, un énorme Merci à Magali pour tout !
Table des matières
I/ Introduction ... 1
II/ Matériel et méthodes ... 3
2.1. Prélèvement des racines et lavage ... 4
2.2. Mesure des traits racinaires ... 5
2.3. Trait et production de biomasse des parties aériennes ... 5
2.4. Analyses statistiques ... 6
III/ Résultats ... 7
2.1. Dynamique temporelle de la production de biomasse racinaire ... 7
2.2. Effet de la température du sol sur la production racinaire ... 8
2.3. Effet de la fertilisation NPK sur les productions racinaires et aériennes ... 9
2.4. Analyses des traits agrégés ... 10
2.5. Relations entre les traits et masses sèches racinaire et aérienne ... 11
IV/ Discussion ... 12
4.1.Dynamique saisonnière de la production racinaire ... 12
4.2. Production et allocation de biomasse entre les parties aériennes et racinaires ... 13
4.3 Effet des traitements sur les traits moyens des communautés ... 13
4.4 Lien entre trait fonctionnel et production de biomasse ... 14
4.5 Regard critique ... 15
V/ Conclusion ... 15
VI/ Bibliographie ... 16
I/ Introduction
Le sol des écosystèmes terrestres est le support du développement des végétaux, et il a une importance majeure dans les agroécosystèmes en agissant sur la production végétale par ses caractères physiques, chimiques et biologiques (Gobat, 2010). Mais le sol rend d’autres services aux activités humaines : la rétention et la dégradation des polluants, la capacité à accumuler des matières organiques et donc du carbone en grande quantité. Augmenter le stockage du carbone dans les systèmes prairiaux permet de compenser en partie les émissions liées aux pratiques agricoles qui représentent 21% des émissions totales françaises (www.developpement-durable.gouv.fr). Généralement, les stocks de carbone organique les plus élevés sont observés dans les sols de prairies, dans les forêts et les milieux à végétation arbustive et/ou herbacée. Ils résultent principalement des allocations de carbone au sol par l’exsudation racinaire (libération de composés organiques par les racines), les tissus végétaux morts (racines, feuilles et tiges sénescentes) et les activités microbiennes associées (Jackson et al., 1997, cité par Herfurth, 2015). Selon leur nature (prairie permanente ou temporaire) et leur mode de gestion (pâturage, fauche, pratiques de fertilisation), les prairies peuvent être des sources ou des puits de carbone (Klumpp et al., 2013). La production de biomasse racinaire est modulée par les pratiques de gestion (Loiseau et al., 1995), qui influent donc elles-mêmes sur la dynamique de stockage du carbone. Une meilleure compréhension des effets des pratiques de gestion en prairie permanente sur la production racinaire permettrait de mieux comprendre la dynamique de stockage du carbone en prairie.
Au niveau aérien, la composition botanique, la dynamique de végétation mais aussi le niveau de production dépendent de facteurs de gestion des prairies tels que la gestion de la disponibilité en nutriments (fertilisation) et du niveau de perturbation appliqué (Louault et al., 2005).Par exemple, la richesse floristique diminue face à l'augmentation de la fertilisation azotée (Muller, 2003), tandis que la biomasse aérienne augmente (Pontes et al., 2007). Picon-Cochard et al, 2009, ont montré que la production de biomasse racinaire de graminées en monoculture est stimulée par une faible fréquence de fauche et un faible apport azoté en comparaison à une plus forte fréquence de fauche et des apports azoté. Ces résultats contrastés de réponse des parties aériennes et racinaires peuvent s’expliquer par le concept « d’équilibre fonctionnel » de l’allocation de biomasse (Brouwer, 1963; Thornley, 1972; Iwasa et Roughgarden, 1984, cité par Poorter et al., 2011). Les plantes vont allouer plus de biomasse vers les racines si le facteur limitant pour la croissance est souterrain (e.g. nutriment, eau), ou au contraire vers les parties aériennes si le facteur limitant est aérien (e.g. lumière, CO2) (Poorter et al., 2011).
La caractérisation d’un couvert végétal de prairie permanente est souvent complexe, de fait notamment de la diversité des espèces qui peuvent la composer. Les approches de caractérisation des couverts et des espèces à travers une approche fonctionnelle des traits permet de caractériser l’état d’une espèce face à un facteur environnemental, tel que le mode de gestion agricole. On regroupe sous le terme de traits fonctionnels « toute caractéristique morphologique, physiologique ou phénologique mesurable au niveau de l’individu, de la cellule à l’organisme dans son ensemble (Violle et al., 2007). Les « traits de réponse » sont définis comme étant les traits dont les mesures diffèrent en réponse à un changement environnemental et les « traits d’effet » comme étant liés à des modifications de propriété et de fonctionnement des plantes ou de l’écosystème (Keddy 1992, Diaz et Cabido 2001, Lavorel et Garnier, 2002, cité par Violle et al., 2007). Les valeurs de traits fonctionnels marquent généralement des compromis entre les différentes fonctions pouvant être accomplies par une plante, déterminant des « stratégies » (Lavorel et al., 2007). Ainsi, les espèces à croissance rapide (stratégie d’exploitation) sont notamment caractérisées par des valeurs élevées de surface spécifique foliaire (SLA) et les espèces à croissance lente (stratégie de conservation) présentent quant à elles des caractéristiques opposées avec notamment des valeurs de matière sèche des limbes (LDMC) élevées (Wright et al., 2004 ; Diaz et al., 2004, cité par Chanteloup, 2013). De même, au niveau racinaire, une forte densité racinaire (RTD) et un grand diamètre racinaire favorise la longévité des racines, caractéristiques retrouvées chez les espèces à croissance lente (Roumet
et al., 2006). A l'inverse, des espèces adoptant une stratégie d'acquisition rapide des ressources
ont une longueur spécifique des racines (SRL) plus élevée, une forte concentration en azote dans les tissus (RNC) et un taux de respiration plus élevé que chez les espèces à croissance lente (Picon-Cochard, et al., 2012). Cependant, il existe des réponses contrastées entre traits foliaires et traits racinaires lorsque les nutriments du sol sont modifiés. En faible apport d’azote dans le sol, les traits foliaires portent des valeurs liées à la stratégie de conservation des ressources (Pontes et al., 2007), tandis que les traits racinaires portent des valeurs liées à la stratégie d’acquisition rapide des ressources (Picon-Cochard et al., 2009). Cependant, peu d’études ont été réalisées sur l’effet combiné des facteurs environnementaux et les modes de gestion sur les traits morphologiques des systèmes racinaires des espèces prairiales (Roumet et al., 2006). La plupart des études ont été réalisées en condition de monoculture, permettant difficilement de simuler des processus tels que les dynamiques de compétition souterraine ente espèces pour les ressources en eau et nutriments.
Au-delà de l’individu dans le couvert, la caractérisation fonctionnelle d’une communauté végétale peut se faire à l’aide du calcul d’une valeur de trait agrégé à partir de la valeur des
traits des différentes espèces et de leurs abondances relatives dans la communauté (Violle et
al., 2007). Cette étude a pour finalité de comparer la dynamique de croissance racinaire et
aérienne, pour différents niveaux de fertilisation en prairie permanente de fauche, et de voir si cette disponibilité en nutriments détermine une allocation différente de biomasse entre les compartiments aérien et racinaire. Pour cela, nous allons tester l’hypothèse 1 : en gestion de fauche sans restitution minérale (F-), la production de biomasse racinaire sera plus forte et la production de biomasse aérienne sera plus faible qu'en gestion de fauche avec restitution minérale (F+). Ensuite, nous testons l'hypothèse 2 : en gestion F-, les traits agrégés aériens reflètent une stratégie de conservation des ressources et les traits racinaires reflètent une stratégie d'acquisition des ressources. Ensuite nous allons déterminer les valeurs de traits agrégés au niveau aérien et souterrain pour analyser l'effet des traitements sur les communautés. Nous voulons identifier si les traits étudiés se différencient et correspondent à des stratégies vis à vis des ressources et s’ils sont en lien avec les performances de production.
II/ Matériel et méthodes
L’étude a été réalisée sur le Système d’Observation et d’Expérimentation sur le long terme pour la Recherche en Environnement – Agroécosystèmes, Cycles Biogéochimiques et Biodiversité (Commune de Saint-Genès Champanelle, Puy de Dôme, France ; 45°43' N, 3°1' E), à 870 m d’altitude, avec une température moyenne annuelle de 8,7°C et un niveau de précipitation annuelles de 800 mm. Avant l'installation du dispositif, en 2005, le site de prairie permanente était utilisé en gestion mixte combinant fauche et pâture. Deux blocs ont été définis par les caractéristiques du sol, avec pour un premier bloc, un sol de type cambisol eutric et pour un deuxième bloc, un sol de type cambisol colluvique (Plan annexe 1). Des analyses physico-chimique du sol ont été effectués en 2005 et 2012 sur chaque parcelle étudiée (Annexe 2). Les parcelles expérimentales de prairie permanente ont été délimitées puis gérées à partir de 2005 de manière différenciée selon différents modes de gestion afin d'analyser l’évolution du fonctionnement de l'écosystème sur le long terme. Les modes de gestions affectent le milieux selon deux gradients : un gradient de perturbation (intensité de pâturage) et un gradient de fertilisation (restitution minérale après fauche). Sur le gradient de fertilisation, les parcelles sont fauchées trois fois par an, les dates moyennes sur 10 ans étant les 27/05, 20-30/07 et 15-20/10. L'étude a été conduite sur ce gradient de fertilisation, en comparant un traitement sans restitution minérale (F-) après la fauche et un niveau de fertilisation avec restitution d'azote, phosphore et potassium après la fauche (F+). Les mesures ont été faites sur la période du
5/12/2014 au 29/05/2015. On définira ensuite les périodes de croissance par la période 1 (5/12/14 au 2/03/15), la période 2 (2/03/15 au 30/03/15), la période 3 (30/03/15 au 24/04/15) et la période 4 (24/04/15 au 29/05/15). Sur chaque bloc, deux traitements (F-, F+) sont répétés deux fois, et quatre emplacements par parcelle ont été déterminés aléatoirement pour les relevés de végétations et les racinaires, soit un total de 32 prélèvements répartis sur 8 parcelles : 4 en F- et 4 en F+. Chaque parcelle est équipée d'une sonde permettant un suivi journalier de la température du sol à 20 cm de profondeur.
2.1. Prélèvement des racines et lavage
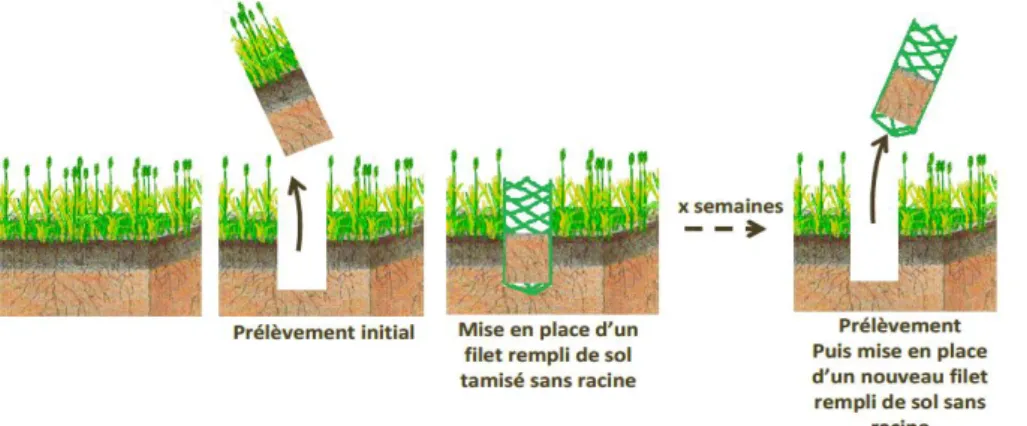
Pendant la période de repos hivernal (5/12/2014), quatre Ingrowth-core (IGC) par parcelle ont été implantés. Ce sont des dispositifs permettant de récupérer les racines qui ont poussé dans un sol sans racines pendant un laps de temps donné. Ainsi on peut mesurer la production de biomasse racinaire en un temps donné pour un volume connu (Filet en plastique à maille de 8mm, 20 cm de profondeur, 8 cm de diamètre). Les filets sont introduits dans un trou préalablement creusé avec un carottier ( 8 cm de diamètre et 20 cm de profondeur) puis rempli de terre provenant du même milieu, stockée au préalable et tamisée à 5 mm. Après leur mise en place, les IGC ont été extraits une première fois en fin d’hiver (2/03/15) puis remplacés chaque mois (30/03/15, 24/04/15, 29/05/2015) (figure 1). Une fois récupérés sur le terrain, les échantillons sont conservés au congélateur à - 18°C en attente du lavage. Le lavage et le tri des racines se réalisent par un lavage à grande eau et trempages successifs dans lequel on utilisera deux tamis superposés (1mm puis 0,2 mm). Dans un récipient, les racines vivantes et les rhizomes sont séparés. Les racines fraîches sont pesées puis séchées à 60°C pendant 48h afin de peser la masse sèche et de déterminer la teneur en matière sèche (RDMC).
Fig.2- Lavage des racines. a) Tamis superposés pour le lavage. b) Échantillon lavé. (Photo, Moreau Paul)
2.2. Mesure des traits racinaires
Pour les échantillons prélevés le 29/05/2015, la préparation est effectuée selon la méthode décrite par Revaillot et Picon-Cochard 2005. Un sous-échantillonnage représentatif de l’ensemble des classes de tailles des racines est effectué, puis les racines sont colorées au bleu de méthylène, rincées dans l’eau pour enlever l’excès de colorant, démêlées et étalées dans une cuve en verre remplie d'un fond d'eau. 3 et 5 scans ont été réalisés par sous échantillon correspondant à un emplacement par classes de taille des racines (grosse, moyenne, fine), ce qui permet de minimiser l’écart de contraste. Ensuite, elles sont recouvertes d'un film transparent permettant de fixer la préparation, puis numérisées avec un scanner (Epson perfection V700, Fig. 3) équipé d'un capot lumineux permettant de faire passer la lumière dans les deux directions et limiter les effets d’ombrage et à une résolution de 800 dpi. Les images sont analysées avec le logiciel WinRhizo (2012b, Regent Instruments, Québec, Ca), permettant de calculer la longueur racinaire (mm), le diamètre (mm) des racines par classe de taille (classe de 0,1 mm) et le volume (cm3). Après le scan, les échantillons sont séchés 48h à 60°C et pesés. Sont alors déterminées la longueur spécifique des racines (SRL, longueur par unité de masse sèche, m g-1), la surface spécifique racinaire (SRA, surface racinaire par unité de masse sèche, cm² g-1), la densité des tissus racinaires (RTD, masse sèche par unité de volume, g cm-3) et le diamètre racinaire moyen (Diam, mm). Pour l’analyse de la longueur par classes de diamètre, les racines ont été regroupées en deux classes de taille, fines : < 0.2 mm et grosses : > 0.2 mm.
Fig.3 Etapes de l’analyse morphologique : disposition des racines dans une cuve étanche contenant de l’eau (a), Positionnement de la cuve sur le scanner (b) et traitement des images scannées (c). (Source, rapport M1 Axel Bartout)
2.3. Trait et production de biomasse des parties aériennes
La production de biomasse aérienne est mesurée lors de la première fauche de l’année le 27/05/2015. Les prélèvements de biomasse sont effectués sur la couronne, autour de l’IGC, un périmètre délimité par un cercle d'un diamètre de 20 cm, centré sur IGC. La hauteur de coupe est de 5.5cm, pour ne pas prélever les collets des graminées (zone de transition entre les tiges et les racines). Ces prélèvements sont triés par espèces, séchés à 60°C puis pesés, ce qui permet d’obtenir une contribution par espèce à la biomasse récoltée autour de l’IGC (couronne). Pour chaque couronne, la contribution (%) de chaque espèce a été déterminée, et seules les espèces dominantes, c’est à dire celles dont la somme des contributions atteint 85%, ont été prises en compte pour le calcul des traits agrégés. Les traits agrégés (moyenne pondérée) de chaque couronne ont été calculés à partir d'une base de données locale de valeur de trait par espèce et par parcelle acquise en 2006, 2007 et 2008 sur le dispositif SOERE de Theix pour les traits suivants : le taux de matière sèche des limbes (LDMC= Leaf Dry Matter Content, masse sèche du limbe divisée par la masse fraîche, g.g-1), la surface spécifique des limbes (SLA= Specific Leaf Area, surface foliaire par unité de masse sèche, cm².g-1) et la hauteur reproductive (H, cm), mesurée selon la méthode de Pérez-Harguindeguy et al., 2013. Ces valeurs de traits agrégés des parties aériennes permettront de les associer à celles de l'échantillon racinaire. Un tableau des contributions (pourcentage) de la composition spécifique en fonction des deux traitements est disponible en annexe 3.
Fig.4- Couronne délimitant les prélèvements de biomasse autour de l’ingrowth-core (20 cm de diamètre). (Photo, Moreau Paul)
2.4. Analyses statistiques
Les analyses statistique ont été effectuées avec le logiciel R (Rstudio, integrated development environment for R ; Version i386 3.2.4). Pour l’analyse des données de biomasse racinaire des IGC mesurée 4 fois, ont été testé l'effet des facteurs traitement (F+, F-), date, bloc et leurs interaction par une ANOVA selon un modèle linéaire à effets fixes sur la variable masse racinaire sèche. Le même modèle a été utilisé sur la variable température du sol. Les donnée masse racinaire et température du sol ont été transformées en log pour la normalité des résidus. Le
bloc est volontairement considéré comme un facteur fixe car des différences de composition pédologique, et notamment de fertilité sont connues. Les effectifs analysés sont n= 4 parcelles par date et par traitement (moyenne des quatre répétitions par parcelle). Pour l’analyse des données sur la production des masses sèche aérienne, racinaires et du ratio racine/aérien, l'effet des facteurs traitement et bloc et de l’interaction traitement x bloc ont été testés par une ANOVA selon un modèle linéaire à effets fixes, de même pour l’analyse des traits agrégés aérien et racinaires. Pour les traits agrégés, le diamètre, les classes de taille C1 et C2, la RTD, LDMC et la RDMC ont été transformés en arc-sinus car les valeurs sont comprises entre 0 et 1. Pour chaque analyse, la normalité et l'homoscédasticité ont été vérifiées par les tests de Bartlett et de Shapiro–Wilk sur les résidus. Les tests Post-hoc de Tukey ont été utilisés avec la fonction « lsmean » (package lsmeans, Russell, 2016). La valeur des traits agrégés aériens a été estimée sur la fauche du 27/05/15, et les traits agrégés racinaires ont été estimé sur la période de croissance de mai 2015 (IGC du 29/05/15). Le cumul des 4 masses de racines récoltées dans les IGC aux 2/03/15, 30/03/15, 24/04/15 et 29/05/15 permet d’estimer la production racinaire, et la mesure des parties aériennes récoltée une fois au 27/05/15 permet d’estimer la production aérienne sur la même période.
Une analyse multivariée en composante principale (ACP) a été effectuée sur les variables des traits agrégés racinaires, aériens, la masse sèche racinaire (IGC du 29/05/15) et la masse sèche aérienne (package ade4, Dray S et al, 2016).
III/Résultats
3.1. Dynamique temporelle de la production de biomasse racinaire
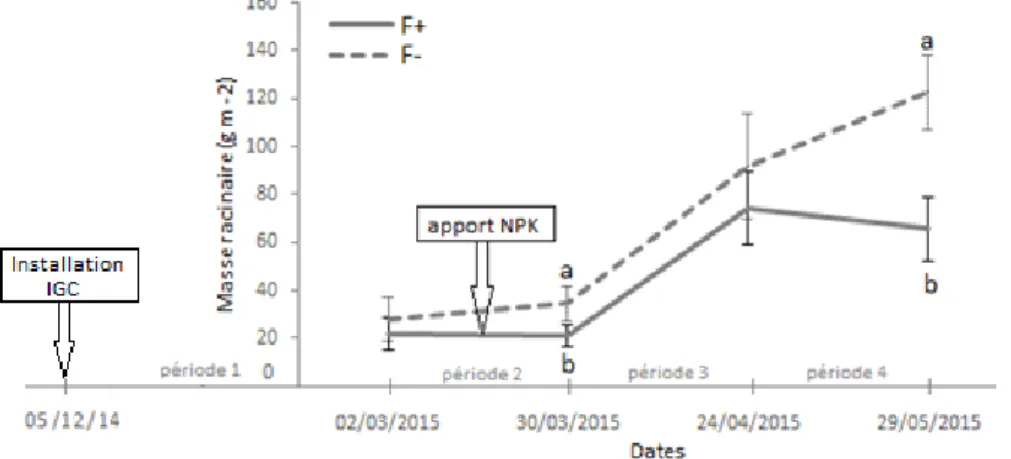
La figure 5 représente la production de masse sèche racinaire (g m-2) mesurée par les ingrowth-core entre deux dates de relevé. Pour les deux traitements, la production racinaire est faible pendant l’hiver (période 1) ainsi qu’en mars, puis devient importante au printemps (avril), suivi d'une décroissance de production observée pour le traitement F+ en mai. La dynamique de croissance racinaire évolue de la même manière entre les deux traitements, jusqu'au 24/04/15. A cette date, un pic de production maximum est atteint pour le traitement F+ (74,3 g m-2) tandis que la production pour le traitement F- augmente encore sur la date suivante. La production de biomasse est significativement plus élevée à la date du 24/04/15 que celle du 30/03/15, (*** : p<0.0001). Globalement, la production de masse racinaire est plus élevée pour le traitement F-, avec des différences significatives observées au mois de mars (** : p< 0,001) et au mois de mai (*** : p<0,0001). Cependant, pour chaque traitement, un effet bloc est constaté, le bloc 1 étant
moins productif que le bloc 2. Les différences significatives de production racinaire entre traitement sont observées uniquement dans le bloc 2 avec une différence significative de production racinaire en gestion F- pour la période 1 (*** : p<0.0001), antérieure à la mesure du 02/03/15 et pour la période 2 (* : p<0.05), antérieure à la mesure du 30/03/15.
Fig. 5- Biomasse racinaire (g MS m-2) produite dans les ingrowth-core à chaque date de prélèvement, pour les traitements de fauche
fertilisée F+ (trait continu) et de fauche non fertilisée F- (trait discontinu). La période 1 est située entre le 5/12/14 au 2/03/15; la période 2 : 2/03/15 au 30/03/15 ; période 3 : 30/03/15 au 24/04/15 ; période 4 : 24/04/15 au 29/05/15. Les barres d’erreurs représentent l’erreur standard. Pour chaque date, les lettres (a, b) représentent les différences de traitements mises en évidence par les contrastes du modèle mixte. Pour chaque date, les valeurs sont calculées par la moyenne des 4 répétitions par parcelle, puis par la moyenne ± erreur standard des 4 parcelles de chaque traitement.
3.2. Effet de la température du sol sur la production racinaire
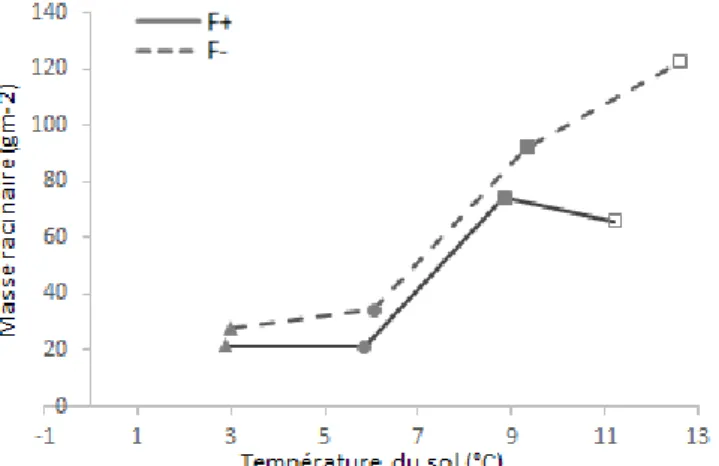
Les capteurs de température installés sur le dispositif SOERE ACBB site de Theix ont permis de mesurer pour les deux traitements (F+ et F-) la température du sol par parcelle à 20 cm de profondeur. On observe une différence significative de température moyenne pour la période 4 (** : p<0.001) avec une température moyenne significativement plus élevée pour le traitement F- (12,6 °C) que pour le traitement F+ (11,2 °C), (** : p<0,001) (Fig 6). Il n’y a pas d’effet bloc sur la température du sol au cours des 4 périodes. Si on considère la biomasse racinaire produite pour une température moyenne donnée, on observe qu’elle est toujours supérieure en F- sur la période (écart de + 6,1 g m², + 19,6 g m², + 37,3 g m², + 94 g m², respectivement pour les périodes 1 à 4).
Fig. 6- Masse racinaire (g m-2) mesurée au cours des quatre périodes de mesures en fonction de la température moyenne journalière du
sol. Les températures sont mesurées à une profondeur de 20 cm sur les parcelles en fauche fertilisée (F+, trait continu) et en fauche non fertilisée (F-, trait discontinu). ( période 1, période 2, période 3, période 4).
3.3. Effet de la fertilisation NPK sur les productions racinaires et aériennes
Sur la période de décembre à mai, le cumul de la masse racinaire produite (g m-2) montre une différence significative entre les traitements F+ et F- (p<0,01), avec une production plus forte pour le traitement F- (277 g m-2) que le traitement F+ (181 g m-2) (Fig 7a). Pour la production de biomasse aérienne, il existe aussi une différence significative entre les deux traitements (p<0,01), le traitement F- étant moins productif que le traitement F+ (Fig. 7a). Le rapport masse racinaire sur masse aérienne est significativement différent entre les traitements F+ et F- (p<0,001). Pour le traitement F+, le ratio indique que 35% de la biomasse produite est accumulé au niveau des racines, contre 79% pour le traitement F- (Fig. 7b).
Fig. 7- (a) Productions racinaires et aériennes (g m-2) cumulées sur la période du 05/12/2014 au 29/05/2015 en fonction des traitements
de fauche fertilisée (F+, noire) et de fauche non fertilisée (F-, gris). (b) Rapport du cumul de la masse racinaire sur la masse aérienne, sur cette période. La masse sèche aérienne a été déterminée lors de la première fauche de l’année, le 27/05/2015, et est mesurée sur le couvert autour de l’ingrowth-core. Les barres d’erreurs représentent l’erreur standard. Les différences de traitements sont signalées par les étoiles (*: p<0.01 ; ** : p< 0.001).
3.4. Analyses des traits agrégés
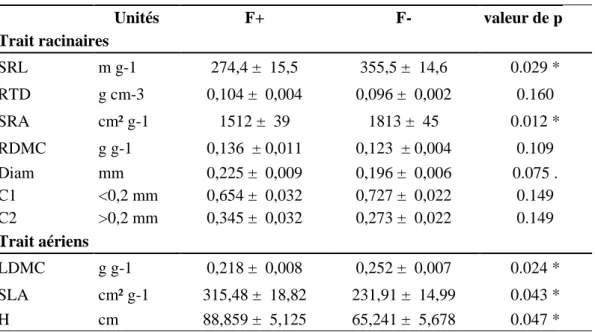
Le tableau 1 présente les valeurs des traits agrégés des communautés végétales, mesurées à partir des espèces dominantes de la couronne des ingrowth-core (fauche de la couronne de l’IGC du 27/05/15) et les valeurs des traits racinaires calculées sur la période 4 (prélèvement du 29/05/15), pour les traitements F- et F+. Les analyses montrent que le facteur traitement à un effet significatif sur les traits des parties ariennes et sur quelques traits des parties racinaires. Pour les traits racinaires, la longueur spécifique des racines (SRL) et la surface spécifique racinaire (SRA) sont significativement plus importantes pour le traitement de fauche F- (* : p<0,05) qu'en fauche F+. La proportion de racines fines (C1) à tendance à être supérieure en F-, alors que les autres valeurs de traits ont tendance à être inférieures en F-, telles que la densité racinaire (RTD), le taux de matière sèche des racines (RDMC), le diamètre moyen (Diam) et la proportion de grosses racines (C2). L'effet traitement est significatif sur les trois traits aériens mesurés. En F-, la teneur en matière sèche des feuilles (LDMC) est plus forte (* : p<0,05), la surface spécifique foliaire (SLA) et la hauteur reproductive (H), sont plus faibles (* : p<0,05) qu’en F+.
Tableau 1. Effet des traitements en fauche fertilisée (F+) et en fauche non fertilisée (F-) sur les traits agrégés racinaires et les traits agrégés aériens mesurés sur la période 4 au 29/05/15. Les traits racinaires sont représentés par les variables suivantes : longueur spécifique des racines (SRL, m g-1), densité des tissus racinaires (RTD, g cm-3), la surface spécifique racinaire (SRA, cm² g-1), le taux
de matière sèche des racines (RDMC, g g-1), le diamètre moyen des racines (diam, mm), et la proportion de racines en pourcentage
dans les classes de diamètre <0.2 mm (C1) et >0.2mm (C2). Les traits aériens sont représentés par les variables suivantes : le taux de matière sèche des feuilles (LDMC, g g-1) la hauteur reproductive (H, cm), la surface spécifique foliaire (SLA, cm² g-1). Les traits
aériens sont représentés par les variables suivantes : le taux de matière sèche des feuilles (LDMC, g g-1) la hauteur reproductive (H,
cm), la surface spécifique foliaire (SLA, cm² g-1). Les différences entre les traitements ont été testées par un modèle linéaire à effets
fixes. Pour chaque trait, les valeurs sont calculées par la moyenne des 4 répétitions par parcelles, puis par la moyenne des quatre parcelles à chaque traitement, (moyenne ± erreur standard). Les différences de traitements sont signalées par un astérisque (* : p< 0.05). Effet du traitement Unités F+ F- valeur de p Trait racinaires SRL m g-1 274,4 ± 15,5 355,5 ± 14,6 0.029 * RTD g cm-3 0,104 ± 0,004 0,096 ± 0,002 0.160 SRA cm² g-1 1512 ± 39 1813 ± 45 0.012 * RDMC g g-1 0,136 ± 0,011 0,123 ± 0,004 0.109 Diam mm 0,225 ± 0,009 0,196 ± 0,006 0.075 . C1 <0,2 mm 0,654 ± 0,032 0,727 ± 0,022 0.149 C2 >0,2 mm 0,345 ± 0,032 0,273 ± 0,022 0.149 Trait aériens LDMC g g-1 0,218 ± 0,008 0,252 ± 0,007 0.024 * SLA cm² g-1 315,48 ± 18,82 231,91 ± 14,99 0.043 * H cm 88,859 ± 5,125 65,241 ± 5,678 0.047 *
3.5/ Relations entre les traits et masses sèches racinaire et aérienne
Une analyse en composantes principale (ACP) a été réalisée en considérant les 32 IGC comme individus (4 IGC par parcelle pour les 4 parcelles F- et 4 parcelles F+) avec leurs variables descriptives des traits aérien, racinaire, production de biomasse aérienne et racinaire. Les axes 1 et 2 expliquent 51,6 % de la variance totale. D'après la matrice des corrélations, la variable RDMC, corrélée positivement avec la RTD et la variable SRA, corrélée négativement avec la SLA ont été retirées de l'analyse. La valeur des contributions absolues et relatives indiquent que les variables C1, C2, LDMC, SLA, H et Maer (Masse aérienne) sont représentatives de l’axe 1, qui explique 31% de l'inertie. Les variables SRL et Diam sont représentatives de l’axe 2, qui explique 20,6% de l'inertie (Fig. 8a). La figure 8b montre la représentation des valeurs de traits et masses par emplacement en fonction du traitement sur le plan des deux premiers axes de l’ACP. On distingue les IGC des parcelles non fertilisées (F-) à gauche de l'axe principal et les IGC des parcelles fertilisées (F+) à droite de l'axe principal.
Fig. 8- (a) Cercle des corrélations des variables traits aériens, traits racinaires, masses sèches aériennes et racinaires, où l’axe 1 explique 31% et l’axe 2 explique 20,6% de l’inertie totale. (b) Plan factoriel des IGC, selon le traitement de fertilisation F+ et F- où l’axe 1 explique 31% et l’axe 2 explique 20,6% de l’inertie totale. Les abréviations des variables traits aériens et racinaires sont expliquées dans la légende du tableau 1. Les variables masse racinaire (Mrac) et masse aérienne (Maer) viennent compléter les variables des traits aériens et racinaires. Les données de la masse racinaire et les traits agrégés racinaires proviennent de la repousse de mai 2015 (période 4).
IV/ Discussion
4.1 Dynamique saisonnière de la production racinaire.
Le suivi racinaire par la méthode des IGC a permis de mettre en évidence une dynamique saisonnière de la biomasse racinaire sur la période de décembre 2014 à mai 2015. Bien que notre suivi de production racinaire s'arrête fin mai, une étude similaire conduite en pâturage a montré que le printemps est la saison la plus productive, suivi d'une forte décroissance de biomasse racinaire en été (Bulon, 2015). La production racinaire est déterminée par l’activité
des plantes sous l’influence indirecte de plusieurs facteurs pédoclimatiques et de gestion. Plusieurs travaux ont mis en évidence un lien entre la production de racines et la température (DAVIDSON, 1968 ; Garcia-Pausas et al., 2011 ; Pilon et al., 2013). Ceci est confirmé par la relation mise en évidence entre la masse racinaire et la température du sol (Fig. 6). Au début du printemps, le sol et l’air se réchauffent progressivement ce qui stimule la croissance des plantes et les activités de la rhizosphère (exsudats) des plantes augmentant ainsi les activités des microbes du sol (Rasse et al., 2005 cité par Herfurth, 2015). La température peut aussi stimuler la minéralisation de la matière organique par les microbes et augmenter l’accessibilité des nutriments (Fitter et al., 1999, cité par Pilon, 2011). Dans notre cas, nous avons mis en évidence une différence de dynamique entre les traitements F- et F+.
L’augmentation de la température moyenne du sol en F- observé sur la dernière période pourrait être induite par un couvert végétal moins dense qu’en F+, favorisant une meilleure absorption de la lumière par le sol. Cette température plus importante pourrait contribuer à expliquer le maintien d’une plus forte production racinaire en F- qu’en F+ sur la période 4. Ce maintien de la production racinaire en F- pourrait s’expliquer aussi par une allocation plus importante du carbone vers les racines (Farrar & Williams, 1991 ; Wan et al., 2005, cité par Pilon, 2011) pour permettre un maintien ou une augmentation de la capacité d’exploration et d’acquisition des nutriments, devenus limitants pour la plante. Ces résultats vont bien dans le sens des analyses du sol qui montrent un appauvrissement des nutriments dans les parcelles F- par rapport aux parcelles F+ après 6 ans de traitement (annexe 2).
Concernant le traitement F+, dans lequel est constatée en période 4 une baisse de production racinaire, la température ne peut pas expliquer la baisse de production racinaire. Dans ce traitement, on pourrait penser que la croissance aérienne serait favorisée par l’apport de nutriment, apporté en mars. La phénologie des espèces pourrait aussi expliquer une moindre production de racines en F+ observée fin mai si le traitement F+ est composé en majorité d’espèces ayant une floraison plus précoce. En effet, lors de la phase de floraison, il a déjà été montré que les plantes modifient leur allocation de carbone de préférence dans les organes reproducteurs plutôt que dans les racines (Poorter et al., 2011). En analysant la composition botanique et compte tenu des données sur la floraison des espèces de la communauté (annexe 4, Pontes, 2006), on observe une tendance au développement d’un couvert plus précoce en F+ qu’en F- mais une analyse de la phénologie de chaque espèce sur les parcelles est nécessaire pour confirmer ces tendances. En effet, certaines espèces tardives sont retrouvées en F+ et des précoces en F-.
4.2 Production et allocation de biomasse entre les parties aériennes et racinaires
La plupart des espèces investissent une plus grande proportion de matière sèche vers le système racinaire lorsque la teneur en azote et en phosphore du sol sont plus faibles pouvant induire un stress nutritionnel (Boot et Mensink, 1990). Cette stratégie permet d’accroître la surface d’échanges des racines avec le sol et donc d’augmenter la capacité à assimiler ces nutriments. Cette allocation préférentielle de biomasse pour le système racinaire est observée par une production racinaire cumulée supérieure en F- qu’en F+ et par l’augmentation du ratio « biomasse racinaire : biomasse aérienne » en fauche non fertilisée. La compétition pour les ressources est reconnue en grande partie comme un “filtre biotique” affectant les communautés de plantes (Grace et Tilman 1990, Keddy et Nielsen 2002, Kraft et Ackerly 2010, Wang et al., 2010, cité par Fort, 2013). Nous pouvons supposer qu’en F+, la compétition pour l’absorption d’éléments nutritifs est plus faible, se traduisant par un développement racinaire plus faible, tandis que la compétition pour la lumière est prépondérante dans les interactions entre plantes. En effet, la hauteur reproductive, qui est un bon prédicteur des capacités de compétition aérienne pour la lumière (Bullock et al., 2001, cité par Louault et al., 2005), a été plus élevée en F+. Ces résultats sont en accord avec nos hypothèses sous réserve de suivre la production sur un cycle annuel, nos résultats étant exprimés sur une période de 5 mois.
4.3 Effet des traitements sur les traits moyens des communautés
Les traits agrégés rendent compte de l’état des communautés, qui ont évolué dans leurs structures sous l’effet des traitements. Les relevés de végétation permettent de mettre en évidence ces différences de structure (annexe 3). Concernant les traits de réponse aériens en F-, les communautés ont montré une diminution de la SLA et une augmentation de la LDMC, et comme rapporté par Lavorel et al., 2007, sont caractéristiques d’une stratégie de conservation des ressources, avec un faible taux de croissance et une durée de vie des tissus plus longue. Pour faire le lien entre les traits aériens et racinaires, il a été montré qu’une faible LDMC est fortement corrélée avec une forte SLA et une forte SRL chez les graminées (Garnier & Laurent, 1994 ; Ryser et Lambers, 1995, cité par Louault et al., 2005), ce qui va à l’encontre de nos résultats concernant la SRL. Picon-Cochard et al., 2009 expliquent qu’en faible fertilisation azotée, l’accroissement de la biomasse racinaire, et l’augmentation de la longueur des racines permettraient d’améliorer l’efficacité d’acquisition de l’azote. Ces changements de morphologie en réponse à la fertilisation peuvent s’expliquer par la plasticité des racines. Les communautés de prairie sont souvent composées d’espèces qui diffèrent pour leurs besoins en nutriments, et donc dans leurs capacités à modifier leurs systèmes racinaires en réponse au
niveau de fertilisation (Hill et al., 2006). Cette plasticité est considérée comme une modification de la capacité du système racinaire à capturer des ressources, soit en explorant de nouveaux espaces grâce à des changements morphologiques soit physiologiquement en modifiant la capacité d’absorption par unité de surface, ou les deux à la fois. Les résultats d’une étude en praire pâturée (Hill et al., 2006), montrent des résultats contrastés dans la réponse des traits à la disponibilité en phosphore et azote, avec une augmentation de la SRL et diminution de la masse racinaire lorsque la quantité de phosphore diminue, ou bien que la quantité d’azote augmente.
4.4 Lien entre trait fonctionnel et production de biomasse.
Les résultats de l’ACP de l’axe 1 mettent en évidence des stratégies d’acquisition et ou de conservation des ressources opposées entre les parties aériennes et souterraines. Pour les traits racinaires, une partie de l’axe 1 reflète une stratégie d’acquisition rapide des ressources, caractéristique du traitement F-, mis en évidence par une proportion de racines fines plus importante et une masse racinaire élevée, reflétant un taux d’absorption élevé (Roumet et al., 2006). Associé aux traits agrégés aériens, la LDMC (forte en F-) et la SLA (faible en F-) reflètent une stratégie de conservation (Lavorel et al., 2007) cohérente selon la littérature, avec la faible production aérienne observée. L’autre partie de l’axe 1, caractéristique du traitement F+, reflète des stratégies inverses avec une stratégie de conservation des ressources au niveau racinaire : la RTD et la proportion de grosses racines (C2) sont plus importantes qu’en F-. La RTD est corrélée négativement au taux de croissance (Wahl & Ryser, 2000 ; Hummel et al., 2007, cité par Roumet et al., 2006) et positivement à la longévité (Eissenstat & Yanai, 1997 ; Ryser, 1998, cité par Roumet et al., 2006) favorisant la conservation des ressources. Au niveau aérien, les traits reflètent une stratégie d’acquisition rapide des ressources, mais cette fois-ci pour la lumière (hauteur reproductive et SLA plus élevée qu’en F- et LDMC plus faible), donc avec une plus grande production aérienne. Le deuxième axe est également représenté par des facteurs pouvant être reliés avec les stratégies d’acquisition rapide des ressources. Il existe une corrélation négative entre la SRL et le diamètre des racines. Les plantes ayant une forte SRL et un faible diamètre (caractéristique du traitement F-) auraient des taux de respiration élevés et des durées de vie de racines courtes et la capacité d’acquérir rapidement d’importantes quantités de ressources (McCormack et al., 2012, Mommer et al., 2011, Tjoelker et al., 2005, cité par Fort, 2013, Picon-Cochard et al., 2012).
Au final, on constate des stratégies asymétriques de réponse à la fertilisation entre les parties aériennes et racinaires sur nos communautés. Ceci pourrait s’expliquer par le fait que les plantes
doivent trouver un compromis physiologique pour acquérir les ressources les plus limitantes que ce soit pour la lumière en F+ et pour les nutriments (NPK) en F-. Il existe bien un lien entre les traits aériens et la production, en accord avec la littérature et ces stratégies d’acquisition des ressources en F- sont conformes à notre hypothèse. La réponse des traits racinaires reste plus à approfondir, notamment pour les stratégies qui peuvent être en désaccord avec la littérature.
4.5 Regard critique
Le suivi de la production racinaire a pu être étudié par la méthode des IGC. Cependant, cette méthode présente quelques limites. Certaines racines ne peuvent pas être interceptées, telles que les pivots des dicotylédones (par ex. Taraxacum officinale). Seule la croissance semi-horizontale des racines est interceptée. De plus, ces prélèvements induisent un stress pour les plantes.
V/ Conclusion
Classiquement, les expérimentations testant différents niveaux de fertilisation démontrent la plus forte production de biomasse aérienne, mais mettent rarement en parallèle un suivi des biomasses racinaires. Malgré les limites que la méthode des IGC implique, il ont permis de mettre en évidence l’effet des pratiques de gestion, en fauche fertilisée et non fertilisée sur la période de décembre 2014 à mai 2015, que cela soit en termes de dynamique temporelle ou bien sur le cumul de la production souterraine et aérienne. L’analyse des traits agrégés des parties aériennes et racinaires ont montré une divergence de stratégies entre les parties aérienne et racinaire pour l’acquisition des ressources. L’intérêt de l’analyse à l’échelle des communautés est de prendre en considération des processus tels que la compétition, difficilement simulée en conditions de monoculture. Les pratiques de gestions sans apport de fertilisation ont montré une augmentation de la diversité végétale, fournissant de nombreux services écologique telle que l’augmentation des auxiliaires de cultures. Quand a la l’augmentation de la biomasse racinaire, nous pouvons penser qu’elle permet une augmentation du stock de carbone dans les sols prairiaux. De même, un système sans apport de fertilisant réduit les émissions de gaz à effet de serre tel que le protoxyde d’azote. Cependant, dans ce mode de gestion, la production des partie aérienne diminue, d’où la nécessité de trouver un compromis entre le maintien d’une production fourragère et la baisse d’intrants pour une gestion durable des agroécosystèmes.
VI/ Bibliographie
Bartout A., 2014. Dynamique de la croissance racinaire en prairie permanente pâturée par des bovins.Master 1
Biologie et Environnement mention Génomique Ecophysiologie et Productions Végétales Université Blaise Pascal – Clermont-Ferrand
Bulon L., 2015. Dynamique de la croissance racinaire en prairie permanente pâturée par des bovins. IUT Génie
Biologique Option Génie de l’environnement. UNIVERSITE AIX-MARSEILLE.
Boot R.G.A., Mensink M., 1990. Size and morphology of root systems of perennial grasses from contrasting
habitats as affected by nitrogen supply. Plant and Soil 129:2991-299
Chanteloup P., 2013. Patrons d'organisation des traits aériens et racinaires en prairies humides : liens avec la
production primaire (quantité et qualité). Thèse de doctorat, Université de Rennes 1, 211p.
Davidson R.L, 1969. Effect of Root/Leaf Temperature Differentials on Root/Shoot Ratios in Some Pasture
Grasses and Clover. Annals of Botany 33: 561-569
Dray S., Dufour A.B. & J. Thioulouse, 2016. ade4: Analysis of Ecological Data: Exploratory and Euclidean
Methods in Environmental Sciences. R package version 1.7-4
Fort F, 2013. Stratégies d'acquisition des ressources des plantes prairiales sous contraintes hydrique et minérale
- Rôle du système racinaire dans la réponse aux facteurs structurants les communautés. Thèse de doctorat, Université de Toulouse, 215p.
Garcia-Pausas J., Casals P., Romanyà J., Vallecillo S., Sebastià M.T., 2011. Seasonal patterns of
belowground biomass and productivity in mountain grasslands in the Pyrenees. Plant and Soil 340: 315-326. DOI 10.1007/s11104-010-0601-1
Gobat J.M., Aragno M. & W Matthey, 2010. Le sol vivant: Bases de pédologie-Biologie des sols.3 èm Edition.
Chapitre 3. ISBN 978-2-88074-718-3.
Hill J.O, Simpson R.J, Moore A.D & D.F Chapman, 2006. Morphology and response of roots of pasture
species to phosphorus and nitrogen nutrition. Plant and Soil 286:7-19. DOI 10.1007/s11104-006-0014-3
Herfurth D., 2015. Impact des pratiques de gestion sur le stockage du carbone dans le sol des écosystèmes
prairiaux. Thèse de doctorat, Université de Clermont-Ferrand 2.
Klumpp K. et M. Benoit, 2013. Optimiser la gestion des prairies pour favoriser le stockage de carbone, In Étude
Réduction des GES en agriculture. INRA, pp 295-322
Lavorel S., Díaz S. & J Cornelissen, 2007 Plant functional types: are we getting
any closer to the Holy Grail? Terrestrial ecosystems in a changing world, Springer-V (eds J.G. Canadell, D. Pataki & Pitelka), pp 149–164. Berlin Heidelberg.
Loiseau P., Soussana J.F., Casella E. & R. Chaussod, 1995. Evolution des stocks de matières organiques sous
prairies : quantification, évolution, modélisation. Dossiers de l'Environnement de l'INRA 10:57-77
Louault F., Pillar V.D., Aufrère J., Garnier E & J.F Soussana, 2005. Plant traits and functional types in
response to reduced disturbance in a semi-natural grassland. Journal of Vegetation Science 16:151-160
Muller S., 2003. Pratiques agricoles : gérer pour la biodiversité, Espaces naturels. Disponible sur :
Pérez-Harguindeguy N., Díaz S., Garnier E., Lavorel S., Poorter H., Jaureguiberry P.,Bret-Harte M.S., Cornwell W.K., Craine J.M., Gurvich D.E., Urcelay C.,Veneklaas E.J., Reich P.B., Poorter L., Wright I. J.,Ray P., Enrico L., Pausas J.G., de Vos A.C., Buchmann N., Funes G., Quétier F., Hodgson J.G., Thompson K., Morgan H.D., ter Steege H., van der HeijdenS M.G.A., Sack L., Blonder B., Poschlod P., Vaieretti M.V., Conti G., Staver A.C., AquinoX S. & J.H.C. Cornelissen, 2013. New handbook for
standardised measurement of plant functional traits worldwide. Australian Journal of Botany 61:167–234. DOI 10.1071/BT12225
Picon-Cochard, C., Pilon, R. & S. Revaillot, 2009. Plasticity of grass root functional traits and root mass in
response to cutting frequency and N fertilisation. In: 7th ISRR Symposium Root Research and Applications
September 2-4, 2009, Vienna / Austria 31:179-246.
Picon-Cochard C., Pilon R., Tarroux E., Pagès L., Robertson J. & L. Dawson, 2012. Effect of species, root
branching order and season on the root traits of 13 perennial grass species. Plant Soil 353:47-57. DOI 10.1007/s11104-011-1007-4
Pilon R., Picon-Cochard C., Bloor JMG., Revaillot S., Kuhn E., Falcimagne R., Balandier P. & J.F
Soussana, 2013. Grassland root demography responses to multiple climate change drivers depend on root
morphology. Plant and Soil, 364, 395-408.
Pilon R, 2011. Dynamique du système racinaire de l’écosystème prairial et contribution au bilan de carbone du
sol sous changement climatique. Thèse de doctorat, Université Blaise Pascal – Clermont-Ferrand, 268p.
Pontes L. D.A S, Soussana J.F, Louault F., Andueza D. & P. Carrère, 2007. Leaf trait affect the above-ground
productivity of pasture grasses. Functional Ecology 5: 844-853. DOI 10.1111/j.1365-2435.2007.01316.x
Pontes L. D.A S, 2006. Diversité fonctionnelle des graminées prairiales: conséquences pour la productivité et
pour la valeur nutritive. Thèse de doctorat, Université Blaise Pascal, Clermont-Ferrand 2, 337p.
Poorter H., Niklas K.J., Reich P.B, Oleksyn J., Poot P. & L Mommer, 2011. Biomass allocation to leaves,
stems and roots: meta-analyses of interspecific variation and environmental control. New Phytologist 193:30-50. DOI 10.1111/j.1469-8137.2011.03952.x
Roumet C., Urcelay C. & S. Díaz, 2006. Suites of root traits differ between annual and perennial species
growing in the field. New Phytologist 170:357-68. DOI 10.1111/j.1469-8137.2006.01667.x
Russell L, 2016. lsmeans: Least-Squares Means. R package version 2.23
Revaillot S., et C Picon-Cochard, 2005. Méthodes de numérisation et d’analyse d’images de racines de
graminées prairiales. Cahier Technique I.N.R.A. 55:33-44
Vermeer J.G. & F Berendse, 1983. The relationship between nutrient availability, shoot biomass and species
richness in grassland and wetland communities. Plant Ecology 53: 121-126
Violle C., Navas M.L., Vile D., Kazakou E., Fortunel C., Hummel I. & E Garnier E, 2007. Let the concept
of trait be functional ! Oikos 116 : 882-892
http://www.developpement-durable.gouv.fr/Inventaire-des-emissions-de-gaz-a.html. Inventaire des émissions de gaz à effet de serre de la France : les résultats 1990-2012. (Consulté le 15/05/16)
VII/ Annexes
Annexe 1 : Schéma du dispositif expérimental de Theix (Saint-Genès-Champanelle, 63), site prairies
permanentes du SOERE-ACBB. Les parcelles étudiée sont pour le traitement F-(nul) la 4, 11, 18, 25 et pour le traitement F+ (NPK), sont la 6, 11, 20, 27.
Annexe 2 : Analyse chimiques du sol pour la strate 0 à 20 cm de profondeur sur les parcelles gérées en fauche
fertilisée (F+) et en fauche non fertilisée (F-). Les données présentées sont les moyennes ± erreur standard de l’azote (N), le phosphore (P) et le potassium (K), mesurée en 2005 et en 20612. Les unités sont exprimées en gramme par kilogramme de sol. L'effet des facteurs traitement (F+, F-) et bloc ont été testés en interaction par une ANOVA selon un modèle linéaire à effets fixes sur les variables N, P et K. Les différences de traitements sont signalées par (* : p< 0.01) et par (« : P < 0.10)
Année 2005 2012
Traitement F+ F- valeur de p F+ F- valeur de p
Azote (g/kg) 3,47 ± 0,28 3,77 ± 0,26 0.421 3,46 ± 0,221 3,66 ± 0,188 0.428 Phosphore (g/kg) 0,057± 0,006 0,054 ± 0,004 0.635 0,068 ± 0,005 0,047 ± 0,004 0.041 * Potassium (g/kg) 0,230 ± 0,057 0,207 ± 0,031 0.725 0,219 ± 0,021 0,159 + 0,015 0.051 " 5 10 5 15 10 25 20 15 25 30 0 50m 50 25 30 5 10 5 15 10 25 20 15 25 30 0 50m 50 25 0 50m 50 25 30 P1 P2 P3 P45 P4Ov P5Ov P1 P2 P3 P4 P5r P5c Bo+ Bo-Bo+ Bo+ Bo- Bo- Ov-1 1 3 3 Ov-2 2 4 4 5 5 66 7 7 8 8 99 10 10 11 11 13 13 14 14 15 15 16 16 17 17 18 18 19 19 20 20 21 21 22 22 23 23 24 24 12 12 25 25 26 26 27 27 28 28 29 29 Bo+ Bo-30 30 nul pk np k Ab Ab Ab Ab npk npk npk pk pk pk nul nul nul BLOC 1 Blatiere BLOC 2 Moine
ORE Theix Parcellaire : BLOC / TRAIT / n° ID-PAR / REP
Parcelles Pâture : Bovin taux fort (Bo+), Bovin taux faible (Bo-), Ovin taux faible (Ov-) Parcelles Fauche, restitutions nulles (NUL), restitution PK (PK), restitutions NPK (NPK) Parcelles Abandon (AB)
Parcelles Sol Nu (SN) REP1 REP2 REP1 REP2 1 1 ID_PAR de à 3030 TrM TrB
Annexe 3 : Contributions en pourcentage des espèces dominantes (>80%) les communautés végétales par
traitements (F+ et F-), et par bloc (1 et 2).
Traitement
F + F-
Bloc 1 bloc 2 bloc 1 bloc 2
Graminées Agrostis capilaris 0 6,07 8,51 5,07 Alopecurus pratensis 0 8,91 7,07 1,48 Dactylis glomerata 0 0 1,76 5,75 Elytrigia repens 6,66 3,71 0 0 Festuca arundinacea 12,23 8,41 7,83 4,02 Holcus lanatus 10,3 7,07 8,5 2,43 Holcus mollis 3,92 5,74 1,35 0 Lolium perenne 2,92 0 0,83 5,89 Lolium multiflorum 12,01 0 0 0 Poa pratensis 3,52 0 1,3 1,73 Poa trivalis 0 1,54 2,42 3,81 Trisetum flavescens 0 5,74 1,47 1,64 Dicotylédone non légumineuse Achillea millefolium 0 0 2,27 1,02 Cerastium fontanum 0 1,26 2,09 4,66 Hypocheris radicata 0 0 0 3,32 Ranunculus acris 0 0 5,27 0 Taraxacum officinalis 0 0 2,11 2,25 Dicotylédone
légumineuse Trifolium repens 0 0 1,55 2,61
Annexe 4 : Dates des stades de développement phénologique de trois groupes d’espèces de poacée. Les valeurs
représentent la date du stade pour l’espèce la plus précoce et par l’espèce la plus tardive à l’intérieur du groupe. La valeur en gras donne la date moyenne pour le groupe de précocité concerné. Pour chaque groupe, la première ligne exprime des valeurs en jour calendaire, la seconde en degrés jour. Pour le stade végétatif, les valeurs représentent la date de prélèvement de l’échantillon pour la caractérisation des paramètres de valeur nutritive du stade : D’après Pontes, 2006.
Effect of NPK fertilisation on production and trait on plant communities into a permanent graasland under a mowing regime
Abstract- Permanent graasland support plant and animal diversity, forage production or even the ability to store carbon in soils. The dynamics of production of roots play a prominent role in the carbon storage in the ecosystem. However, this effect has not been well documented at the community level. From December 2014 to May 2015, the method of the ingrowth core, allowing for an analysis of roots production in an upland permanent grassland for two levels of fertilization (mowing with fertilizer supply: F+) and mowing without fertilizer supply (F-), has been used. The roots production observed in F- management was higher than in F+. Opposite effect is observed in shoot with a lower production in F- than F+. The root: shoot ratio showed a higher biomass allocation to the roots in F- than F+. Analysis of community-weighted mean traits in roots and leaves of the community has shown that fertilization had a contrasting effect on acquisition resources strategies. In F-, roots traits reflected an acquisition strategy (higher specific root length) and shoots traits reflected a conservation strategy (higher leaf dry matter content). In F+, roots traits reflect a conservation strategy and shoots traits reflect an acquisition strategy. It is necessary to continue this study which tended to show that fertilization has an impact on the plant community both on shoot and root levels. Other factors are involved in carbon storage in the soil of permanent grassland should be considered.
Key world: Permanent grassland, roots and shoot production, community-weighted traits, strategy, mowing
Effet de la fertilisation NPK sur la production et les traits des communautés végétales d'une prairie permanente fauchée
Résumé- Les écosystèmes de prairies permanentes sont reconnus pour leurs nombreux services écosystémiques tels que les supports de diversité végétale et animale, la production fourragère ou bien des capacités de stockage de carbone dans les sols. La dynamique de production des racines joue un rôle déterminant dans le stockage du carbone dans l’écosystème. Cependant, ces thématiques sont peu étudiées à l’échelle des communautés. De décembre 2014 à mai 2015, la méthode des ingrowth-core a permis d’analyser la production racinaire en prairie permanente de moyenne montagne pour deux niveaux de fertilisation (fauche avec apport d’engrais NPK (F+) et fauche sans apport d’engrais (F-)). Nous avons observé que la production racinaire en gestion F- est plus forte qu’en F+. L’inverse est observé pour les parties aériennes avec une production plus faible en F- qu’en F+. Le ratio racine/aérien montre une allocation de biomasse vers les racines plus importante en F- qu’en F+. L’analyse des traits agrégés racinaires et aériens des communautés végétales ont montré que la fertilisation a un effet contrasté sur les stratégies d’acquisition des ressources. En F-, les traits racinaires reflètent une stratégie d’acquisition rapide des ressources (augmentation de la longueur spécifique des racines) et les traits aériens une stratégie de conservation (augmentation de la teneur en matière sèche des limbes). En F+, les traits racinaires reflètent une stratégie de conservation et les traits aériens une stratégie d’acquisition des ressources. Il est nécessaire de continuer cette étude qui tend à montrer que la fertilisation a un impact sur les communautés végétales, tant au niveau aérien qu’au niveau racinaire. D’autres facteurs rentrent en ligne de compte dans le stockage du C dans le sol des prairies permanentes.