The innate immune signaling system as a regulator of disease resistance and induced systemic resistance activity against Verticillium dahliae
11
0
0
Texte intégral
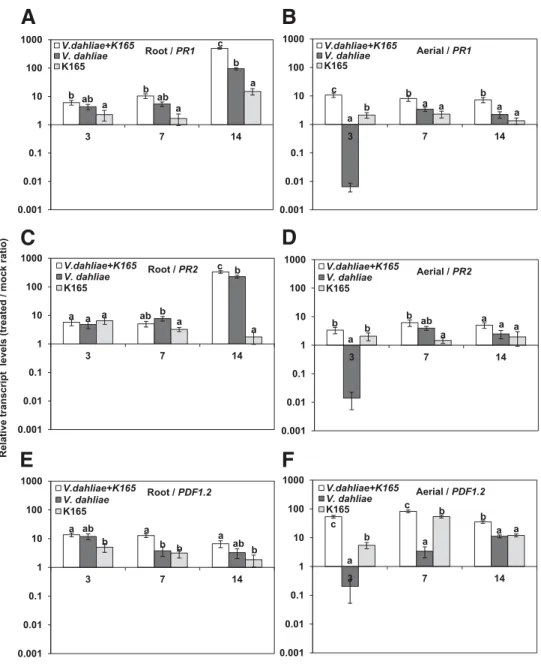
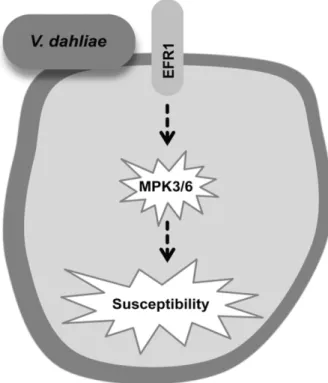
Figure
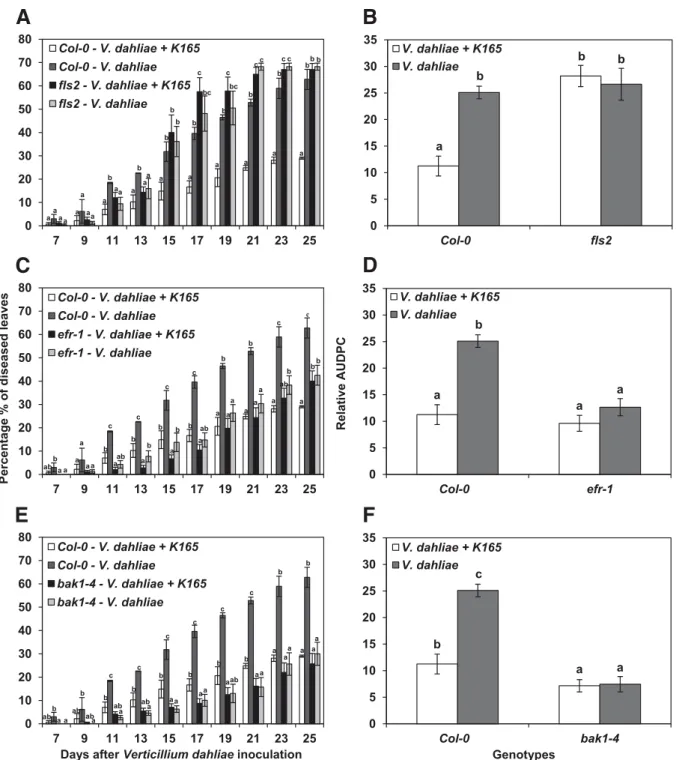
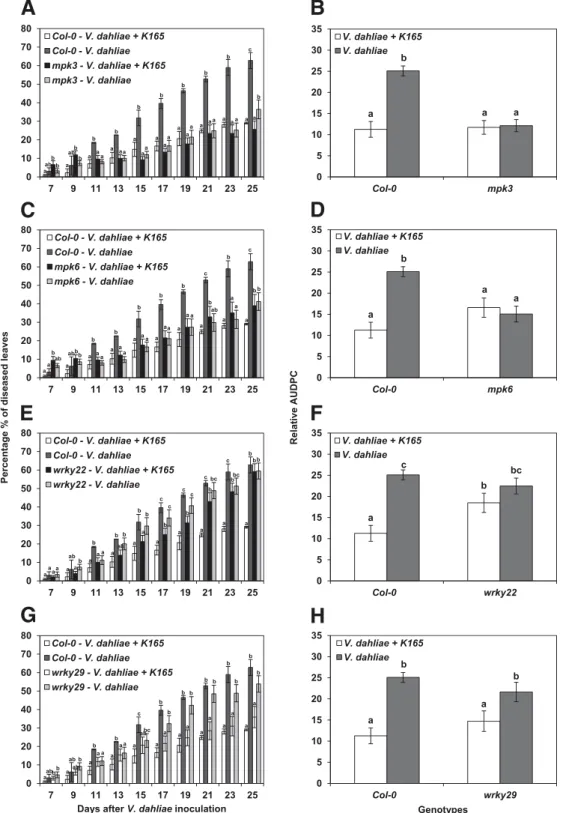
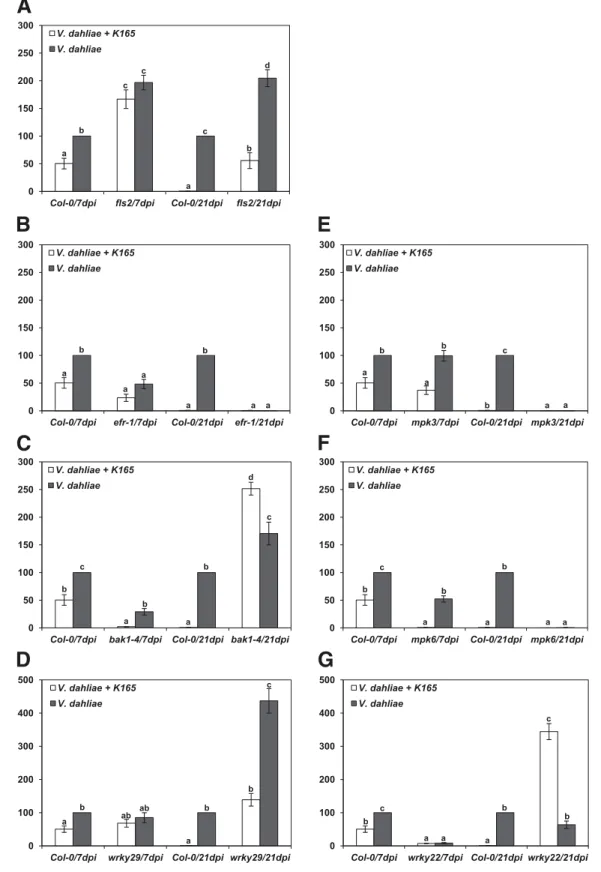
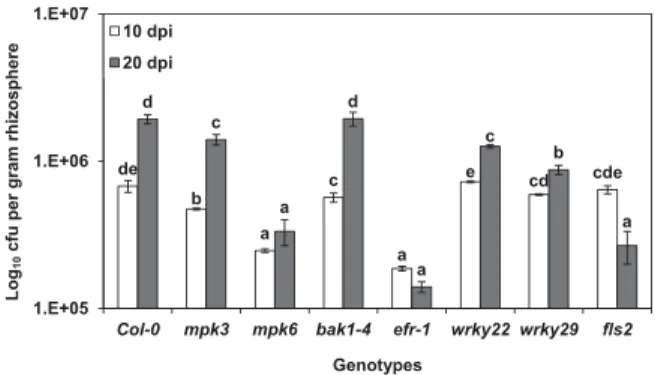
+3
Documents relatifs