MicroRNAs, Major Players in B Cells Homeostasis and Function
Texte intégral
Figure
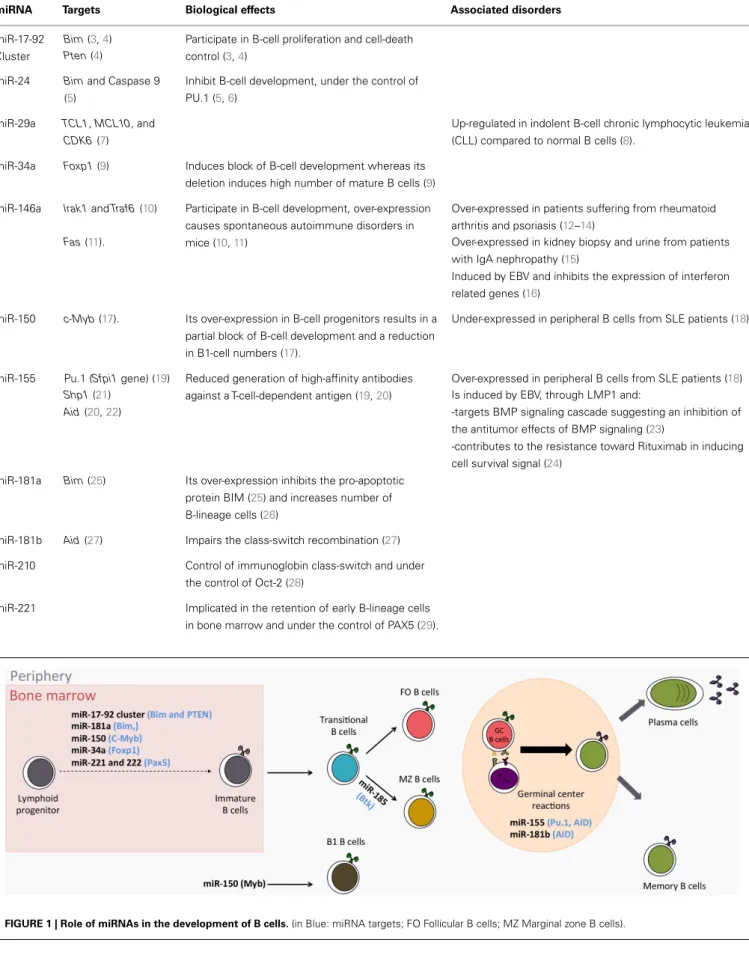
Documents relatifs
The best fT can be achieved by eliminating the body implant while the highest fmax is obtained by also eliminating the drain, but not source, extension and halo implants. Chen,
In 24 hours of test, however, no dampness was seen on the back surface of the inner wall, and no moisture apparently had crossed the cavity.. After 24 hours the wall was removed
L’accès à ce site Web et l’utilisation de son contenu sont assujettis aux conditions présentées dans le site LISEZ CES CONDITIONS ATTENTIVEMENT AVANT D’UTILISER CE SITE WEB.
The volumes of the pose space satisfying the maximum number of correspondences are those that best align the model with the image.. Object motion de nes a trajectory in the
**les recommandations EASD-ADA 2020 privilégient le schéma « basal plus » après échec d’une insuline basale ; le schéma « prémélangé » , dans le diabète de type 2, est
Un enfant demande à son père : “Dis papa c’est maman ou toi qui m’a donné mon intelligence ?” Le père répond : “C’est sûrement ta mère parce que moi je
2) A l’aide d’un algorithme, donner, au jour près, le temps nécessaire pour que le plant de maïs atteigne une hauteur supérieure à 1, 5 m.. 3) On s’intéresse à la vitesse
5 Histograms of link speed estimates versus observations for all time periods (model results are for measured links only).. road), and spatially by categorized values of the
