The non-coding RNA TERRA is a natural ligand and direct inhibitor of human telomerase
10
0
0
Texte intégral
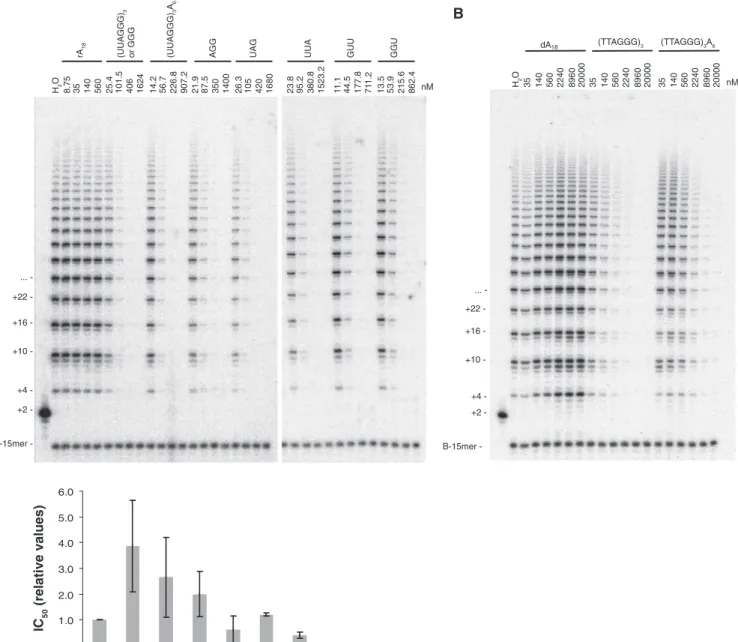
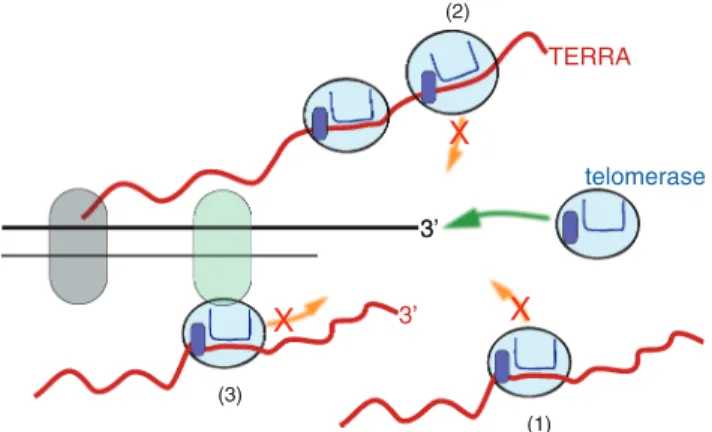
Figure
+3
Documents relatifs
