HAL Id: inserm-00614028
https://www.hal.inserm.fr/inserm-00614028
Submitted on 8 Aug 2011
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of
sci-entific research documents, whether they are
pub-lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destinée au dépôt et à la diffusion de documents
scientifiques de niveau recherche, publiés ou non,
émanant des établissements d’enseignement et de
recherche français ou étrangers, des laboratoires
publics ou privés.
Improved molecular toolkit for cAMP studies in live
cells.
Kwan Hong, Nicholas Spitzer, Xavier Nicol
To cite this version:
Kwan Hong, Nicholas Spitzer, Xavier Nicol. Improved molecular toolkit for cAMP studies in live
cells.. BMC Research Notes, BioMed Central, 2011, 4 (1), pp.241. �10.1186/1756-0500-4-241�.
�inserm-00614028�
T E C H N I C A L N O T E
Open Access
Improved molecular toolkit for cAMP studies in
live cells
Kwan Pyo Hong
1, Nicholas C Spitzer
1and Xavier Nicol
1,2*Abstract
Background: cAMP is a ubiquitous second messenger involved in a wide spectrum of cellular processes including gene transcription, cell proliferation, and axonal pathfinding. Precise spatiotemporal manipulation and monitoring in live cells are crucial for investigation of cAMP-dependent pathways, but existing tools have several limitations. Findings: We have improved the suitability of cAMP manipulating and monitoring tools for live cell imaging. We attached a red fluorescent tag to photoactivated adenylyl cyclase (PACa) that enables reliable visualization of this optogenetic tool for cAMP manipulation in target cells independently of its photoactivation. We show that replacement of CFP/YFP FRET pair with GFP/mCherry in the Epac2-camps FRET probe reduces photobleaching and stabilizes the noise level during imaging experiments.
Conclusions: The modifications of PACa and Epac2-camps enhance these tools for in vitro cAMP studies in cultured living cells and in vivo studies in live animals in a wide range of experiments, and particularly for long term time-lapse imaging.
Background
cAMP is a major cellular second messenger that activates and integrates multiple intracellular signaling pathways and modulates a large range of cellular processes, includ-ing gene transcription [1], cell adhesion and migration [2], and axonal growth and pathfinding [3]. cAMP studies rely on methods to manipulate and monitor cAMP concentra-tions in live cells. Existing tools have been very useful in identifying cAMP-dependent cellular processes, but have some limitations when it comes to understanding cAMP dynamics and localization in living cells. Forskolin and 3-isobutyl-1-methylxanthine (IBMX) are powerful pharma-cological compounds enabling the generation of sustained elevations of cAMP. Forskolin directly stimulates most transmembrane adenylyl cyclases [4] and IBMX inhibits cAMP hydrolysis by phosphodiesterases. Recently, the use of photoactivated adenylyl cyclase alpha (PACa) from the flagellate Euglena gracilis, synthesizing cAMP in response to blue light, has allowed precise spatiotemporal manipula-tion of cAMP [5]. It has been attached to GFP for visuali-zation in live cells [6]. However, the excitation wavelength
of this visible reporter overlaps with the excitation spec-trum of PACa, making it difficult to use this fusion con-struct for independent PACa excitation and reporter imaging.
Monitoring cAMP in live cells has been made possible by the use of FRET probes [7-10]. Epac2-camps is a cAMP indicator that is widely used to monitor cAMP [10] and has been recently improved with a mutation increasing its affinity for cAMP [11]. However, fast photobleaching of the commonly used CFP/YFP FRET pair limits its use in live cell imaging experiments over extended periods of time because the signal-to-noise ratio decreases progressively. The GFP/mCherry FRET pair has been successfully used for cAMP sensors [12], but its photostability and signal-to-noise ratio have not been assessed.
Results
Independent excitation of PACa and mCherry in living cells and live animals
The red fluorescent protein mCherry was expected to be an appropriate tag for PACa since its excitation wavelength in the green range of visible light [13] is dis-tinct from PACa excitation by blue and UV light [14] (Figure 1A). A mCherry-PACa fusion protein was
* Correspondence: [email protected]
1
Neurobiology Section, Division of Biological Sciences, Kavli Institute for Brain and Mind, University of California, San Diego, La Jolla, CA 92093, USA Full list of author information is available at the end of the article
© 2011 Nicol et al; licensee BioMed Central Ltd. This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
generated using a mutant of PACa (R330A) that has a limited adenylyl cyclase activity in the dark (G. Nagel, personal communication). mCherry was attached to the N-terminus of PACa (linker: SGLRSRAQASN-SAVDGTA). The fluorescence of mCherry and light-dependent cAMP synthesis of PACa appear unaffected in the fusion product. mRNA coding for mCherry-PACa was transcribed using the mMessage Ultra kit (Ambion). 1 to 3 ng of mRNA were injected in both blastomeres of 2-cell stage Xenopus laevis embryos, which were incubated in the dark for 24 hr at 23°C. Dissociated cells from stage 21 embryo neural tubes were plated onto plastic dishes and kept in the dark for 2 hr. Cultures from injected but not control animals
were fluorescent when illuminated at 561 nm on a Leica SP5 confocal microscope, revealing the expression of mCherry (Figure 1B). We then developed a bioassay to assess the function of this construct. Circus cells exhibit circular movement of the plasma membrane in cultures from Xenopus neural tubes [15]. Application of 10 μM forskolin blocked the circus movements of these cells (Figure 1B and 1C), demonstrating that a high concentration of cAMP prevents these plasma mem-brane movements. We next tested the light-dependent cyclase activity of mCherry-PACa by illuminating mCherry-positive cells at 488 nm. A 1-minute exposure abolished membrane movements. In contrast, mCherry excitation did not affect circus movements and
A C -+ -14/14 0 20 40 60 80 100 a ct ive ci rcu s ce lls a ft e r st imu la ti o n (% ) PACc light exposure forskolin *** 2/23 3/25 13/13 -+ + + -+ -*** ns B mC h e rry-PAC c light mC h e rry-PAC c W T W T light
1 min 2 min 3 min 4 min 5 min 6 min 7 min 8 min 9 min 10 min forskolin 0 0.2 0.4 0.6 0.8 1 300 400 500 600 700 n o rma lize d e xci ta ti o n mCherry PACc -300 ms 0 ms b lu e ill u mi n a ti o n g re e n ill u mi n a ti o n g re e n ill u mi n a ti o n +300 ms D wavelength (nm) E mCherry-PACc control 0 10 20 30 40 50 60 70 Pro p o rt io n o f re sp o n si ve e mb ryo s (% ) *** *** blue illumination green illumination
Figure 1Characteristics of mCherry-PACa. (A) PACa and mCherry have distinct excitation spectra, predicting independent excitation. (B, C) Sequences of merged panels of transmitted light images, mCherry fluorescence images (red), and 488 nm PACa excitation (blue). The transmission image is omitted during the 488 nm excitation because of saturation of the image (6 min, 2ndand 4throws). Both bath application of 10 μM
forskolin (1strow) and one-minute photoactivation of PACa (2ndrow) induce arrest of circumferential movements of circus cells’ cytoplasm. In
contrast, excitation of mCherry alone (3rdrow) or blue light illumination of cells lacking PACa (4throw) does not affect circus cells’ movements. Arrows indicate the direction of the circus movement in the first image of each row and each time the direction switches. Cells were split in 4 quadrants and were scored as moving when a blob of cytoplasm was seen in all the quadrants during the 4 minutes following stimulation. ≥ 13 cells were scored for each condition. (D, E) mCherry-PACa-injected embryos twitch when illuminated with blue light to excite PACa (2ndrow), but not when exposed to green light to excite mCherry (1stand 3rdrow). Dashed white line marks the initial position of the head of the embryo (-300 ms). Dashed magenta line indicates the head position during (0 ms) and following (+300 ms) the twitch. Embryos that do not express mCherry-PACa are insensitive to both blue and green light. Images were extracted from the movie shown in Additional File 2. Images were chosen 300 ms before (-300 ms), during (0 ms) and 300 ms after (+300 ms) the embryo twitches under blue illumination (BP 470/40 filter). Embryos were immobile under green illumination (BP 560/40 filter), and the 600 ms portions of the recording was chosen randomly (1stand 3rdrow). Embryos
were scored as responsive when they were not completely immobile during the illumination period. ≥ 35 embryos were scored for each condition. Error bars, sem. *** p < 0.001. C, Chi Square test. E, ANOVA.
Hong et al. BMC Research Notes 2011, 4:241 http://www.biomedcentral.com/1756-0500/4/241
mCherry-negative cells did not exhibit plasma mem-brane movement arrest following stimulation by either wavelength (Figure 1B and 1C and see Additional file 1). We conclude that mCherry-PACa combines the features of mCherry for cell identification and PACa for light-sensitive cyclase activity, and that mCherry and PACa excitation wavelengths are exclusive from each other.
To verify that this construct can be used in vivo, Xeno-pusembryos injected with mCherry-PACa mRNA were illuminated with green and blue light in alternation using a fluorescence dissecting microscope with GFP and Texas-red filter cubes. Blue light illumination (excitation filter: BP 470/40) induced embryos to twitch, whereas they remain completely immobile under green light illu-mination (excitation filter: BP 560/40) (Figure 1D and 1E and see Additional file 2). Uninjected embryos did not exhibit light-induced twitching. Excitation of mCherry did not affect embryos’ behavior, confirming the spectral compatibility of mCherry and PACa.
Reduced photobleaching of Epac2-camps using the GFP/ mCherry FRET pair
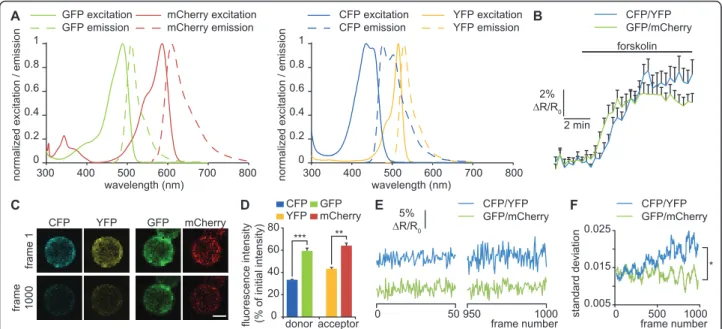
In addition to the improvement of PACa to manipulate cAMP, we modified the FRET pair of the existing Epac2-camps cAMP sensor to improve the stability of its signal-to-noise ratio over longer periods of time. We replaced the CFP/YFP pair by GFP/mCherry, with the same linkers as in the original probe [10]. mCherry and GFP have a low photobleaching rate [16] and constitute an efficient FRET pair [12] with smaller overlap between the emission spectra of the acceptor and the donor (Fig-ure 2A). In addition, we included a mutation in the cAMP binding domain (K405E) that has been shown to reduce the Kd of the probe from 900 nM to 300 nM [11]. To demonstrate the improvement of the sensor, mRNA coding for membrane-targeted Epac2-camps including either CFP/YFP (pm-Epac2-camps-CFP/YFP) or GFP/mCherry (pm-Epac2-camps-GFP/mCherry) was transcribed and expressed in Xenopus laevis neural tube cultures as described for mCherry-PACa. Cultures were continuously superfused and three images per minute were acquired with 442 nm laser excitation and filters adapted for each FRET pair (CFP: 460 nm-500 nm; YFP: 525 nm-580 nm; GFP: 480 nm-550 nm; mCherry: 590 nm-700 nm). 10 μM forskolin induced a similar FRET ratio increase for Epac2-camps-CFP/YFP and pm-Epac2-camps-GFP/mCherry, revealing an appropriate FRET response to cAMP elevation for both probes (Fig-ure 2B). To assess the resistance to photobleaching, cells were continuously excited at 442 nm. CFP and YFP intensity decreased rapidly whereas GFP and mCherry showed reduced photobleaching (Figure 2C and 2D). Although 442 nm is not at the peak of excitation of
GFP, a forskolin-induced increase of the GFP:mCherry ratio can be detected using this excitation wavelength (Figure 2B). Exciting GFP with a 442 nm laser line has the advantage of minimizing the direct excitation of the acceptor (mCherry) that perturbs FRET measurements when longer excitation wavelengths are used. To assess the noise level, we monitored the standard deviation of CFP:YFP and GFP:mCherry FRET ratio changes over 40 frames during the 1000 frames imaging period. The standard deviation of pm-Epac2-camps-GFP/mCherry was stable throughout the measurement period, while the standard deviation of pm-Epac2-camps-CFP/YFP increased over time (Figure 2E and 2F). The progressive increase of the noise level over time may be due to the increased relative contribution of the noise to the fluor-escence measurement of each channel: the fluorfluor-escence intensity decreases because of photobleaching, but the noise level is not affected. Consequently, the noise level of the donor:acceptor ratio is higher after photobleaching.
Conclusion
In summary, we have generated an improved toolkit for cAMP studies. mCherry-PACa allows spatiotemporal con-trol of cAMP in living cells after identification of cells expressing it. Localized illumination of a cell is likely to increase cAMP concentration locally. Further develop-ment of PACa may include its targeting to subcellular compartments to go beyond the limit of precision of opti-cal stimulation and achieve cAMP manipulation bearing closer resemblance to physiological signals. It would be useful to ensure for each experimental condition that the cyclase activity of PACa in the dark does not affect intra-cellular signaling [5,17]. To limit cAMP synthesis without light exposure, we used a mutated PACa (R330A) that has a limited cyclase activity in the dark (G. Nagel, personal communication). This was enough to avoid perturbation of circus cells movement by the cyclase activity of PACa in the dark. In case an extremely low dark activity is needed, bPAC, a bacterial light-sensitive adenylyl cyclase, could be used at the cost of less stringent temporal control of cAMP signaling [17].
pm-Epac2-camps-GFP/mCherry has greater photo-stability than pm-Epac2-camps-CFP/YFP and a lower noise level after extended periods of imaging. However GFP and mCherry make a less effective FRET pair than CFP and YFP, and its use may be beneficial only for FRET experiments requiring an extended period of ima-ging. Versions of CFP and YFP (mTurquoise and Venus respectively) with increased photostability are now avail-able and make an efficient FRET pair for cAMP sensors [18]. Testing its noise level stability would allow compari-son of the behaviour of GFP/mCherry and mTurquoise/ Venus as FRET pairs for prolonged experiments. The
sensor described here has the advantage over the mTur-quoise/Venus probe [18] of sensitivity to lower concen-trations of cAMP, because it is an Epac2-based sensor including a mutation that reduces its Kd [11]. It would be useful to compare it to the Epac1-based sensor using mTurquoise/Venus, with the higher Kd for cAMP [9,10,18].
The use of both tools in the same cell is not yet possi-ble due to the overlap of excitation wavelengths, but further improvements may include the shift of PACa excitation towards the UV to avoid wavelength conflict with the FRET probe excitation, in combination with the switch of the mCherry tag to a longer wavelength fluores-cent protein such as mKate to avoid the overlap of emis-sion between the PACa tag and the FRET acceptor.
Additional material
Additional file 1: Photoactivation of PACa mimics forskolin-induced arrest of circus cells in culture. In circus cells in cultures from the X. laevisneural tube, blebs of plasma membrane propagate around the cell circumference with a period of several minutes (Olson, 1996). Forskolin
stimulation of endogenous adenylyl cyclases in control cells arrests this blebbing movement (top left panel). Blue light (488 nm) irradiation of mCherry-PACa-expressing cells mimics the effect of forskolin stimulation (top right panel). In contrast, blebbing movements are not affected in non-blue light-irradiated mCherry-PACa-expressing cells (bottom left panel) or in control cells illuminated at 488 nm (bottom right panel). Additional file 2: Photoactivation of PACa induces X. laevis embryo twitching. mCherry-PACa-injected embryos twitch when illuminated with blue light to excite PACa, but not when exposed to green light to excite mCherry.
Acknowledgements
We thank Dr M. Roe for the gift of pm-Epac2-camps, and Dr G. Nagel for the gift of PACa. We are grateful to members of our lab for thoughtful discussion. This work was supported by a Fondation pour la Recherche Médicale fellowship and a Marie Curie International Outgoing Fellowship within the 7th European Community Framework Programme to X.N., and by NIH NS15918 to N.C.S.
Author details
1Neurobiology Section, Division of Biological Sciences, Kavli Institute for Brain
and Mind, University of California, San Diego, La Jolla, CA 92093, USA.
2INSERM, U839, Université Paris 6, Institut du Fer à Moulin, 17 rue du Fer à
Moulin, 75005 Paris, France.
A C 1 0.8 0.6 0.4 0.2 0 300 400 500 600 700 800 wavelength (nm) n o rma lize d e xci ta ti o n / e mi ssi o n 1 0.8 0.6 0.4 0.2 0 300 400 500 600 700 800 wavelength (nm) n o rma lize d e xci ta ti o n / e mi ssi o n GFP excitation GFP emission mCherry excitation
mCherry emission CFP emission
CFP excitation YFP emission YFP excitation E 5% ÄR/R0 CFP/YFP GFP/mCherry 0 50 950 1000 frame number fra me 1 fra me 1 0 0 0 mCherry GFP YFP CFP 0 20 40 60 CFP YFP GFP mCherry donor acceptor fl u o re sce n ce i n te n si ty (% o f in it ia l in te n si ty)80 *** ** D F 0.005 0.015 0.025 0 500 1000 CFP/YFP GFP/mCherry frame number st a n d a rd d e vi a ti o n * B CFP/YFP GFP/mCherry 2% ÄR/R0 2 min forskolin
Figure 2Reduced noise level of pm-Epac2-camps-GFP/mCherry compared to pm-Epac2-camps-CFP/YFP. (A) Normalized excitation (solid lines) and emission (dashed lines) of GFP, mCherry, CFP and YFP. Emission spectra of CFP and YFP largely overlap, making FRET measurement difficult. The large separation of GFP and mCherry emission spectra significantly reduces crosstalk between acceptor and donor. (B) ∆R/R0was
computed as (R-R0)/R0where R is donor:acceptor ratio and R0the mean value of this ratio before forskolin stimulation. ∆R/R0of both
pm-Epac2-camps-CFP/YFP and pm-Epac2-camps-GFP/mCherry increase after 10 μM forskolin application. The lag between the pm-Epac2-camps-GFP/mCherry and pm-Epac2-camps-CFP/YFP signals falls under the precision of the perfusion system and is unlikely to reflect a difference between the sensors. No correction factor was applied. ≥ 15 cells were scored for each condition. (C) Acceptor and donor emission of pm-Epac2-camps-CFP/YFP and pm-Epac2-camps-GFP/mCherry before and after 1000 frame acquisitions (excitation with a 442 nm laser line). Photobleaching is less pronounced for pm-Epac2-camps-GFP/mCherry. (D) Fluorescence intensity of acceptors and donors of pm-Epac2-camps-CFP/YFP and pm-Epac2-camps-GFP/ mCherry after 1000 frames, expressed as a fraction of the fluorescence intensity of the first frame. ** p < 0.01, *** p < 0.001. (E) Noise levels for Epac2-camps-CFP/YFP and Epac2-camps-GFP/mCherry are equivalent for the first 50 frames. After 950 frames, only the noise level for pm-Epac2-camps-CFP/YFP has increased. Data are normalized to the value at the beginning of the recording as in B (∆R/R0). (F) The standard deviation
of ∆R/R0over 40 frames, indicating the noise level, shows a progressive increase for pm-Epac2-camps-CFP/YFP but not for pm-Epac2-camps-GFP/
mCherry. * p < 0.05 from frame 819 to 1000. C-F, 5 cells per condition. Error bars, sem. D, Kruskal-Wallis test. F, Mann-Whitney U test. Hong et al. BMC Research Notes 2011, 4:241
http://www.biomedcentral.com/1756-0500/4/241
Authors’ contributions
KPH and XN carried out the experiments. NCS and XN conceived the experimental design. KPH, NCS and XN wrote the manuscript. All authors have read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Received: 6 April 2011 Accepted: 20 July 2011 Published: 20 July 2011 References
1. Sands WA, Palmer TM: Regulating gene transcription in response to cyclic AMP elevation. Cell Signal 2008, 20:460-466.
2. Howe AK: Regulation of actin-based cell migration by cAMP/PKA. Biochim Biophys Acta2004, 1692:159-174.
3. Piper M, van Horck F, Holt C: The role of cyclic nucleotides in axon guidance. Adv Exp Med Biol 2007, 621:134-143.
4. Laurenza A, Sutkowski EM, Seamon KB: Forskolin: a specific stimulator of adenylyl cyclase or a diterpene with multiple sites of action? Trends Pharmacol Sci1989, 10:442-447.
5. Schröder-Lang S, Schwärzel M, Seifert R, Strünker T, Kateriya S, Looser J, Watanabe M, Kaupp UB, Hegemann P, Nagel G: Fast manipulation of cellular cAMP level by light in vivo. Nat Methods 2007, 4:39-42. 6. Weissenberger S, Schultheis C, Liewald JF, Erbguth K, Nagel G, Gottschalk A:
PACα–an optogenetic tool for in vivo manipulation of cellular cAMP levels, neurotransmitter release, and behavior in Caenorhabditis elegans. J Neurochem2011, 116:616-625.
7. Zhang J, Ma Y, Taylor SS, Tsien RY: Genetically encoded reporters of protein kinase A activity reveal impact of substrate tethering. Proc Natl Acad Sci USA2001, 98:14997-15002.
8. Zaccolo M, De Giorgi F, Cho CY, Feng L, Knapp T, Negulescu PA, Taylor SS, Tsien RY, Pozzan T: A genetically encoded, fluorescent indicator for cyclic AMP in living cells. Nat Cell Biol 2000, 2:25-29.
9. Ponsioen B, Zhao J, Riedl J, Zwartkruis F, van der Krogt G, Zaccolo M, Moolenaar WH, Bos JL, Jalink K: Detecting cAMP-induced Epac activation by fluorescence resonance energy transfer: Epac as a novel cAMP indicator. EMBO Rep 2004, 5:1176-1180.
10. Nikolaev VO, Bünemann M, Hein L, Hannawacker A, Lohse MJ: Novel single chain cAMP sensors for receptor-induced signal propagation. J Biol Chem 2004, 279:37215-37218.
11. Norris RP, Ratzan WJ, Freudzon M, Mehlmann LM, Krall J, Movsesian MA, Wang H, Ke H, Nikolaev VO, Jaffe LA: Cyclic GMP from the surrounding somatic cells regulates cyclic AMP and meiosis in the mouse oocyte. Development2009, 136:1869-1878.
12. van der Krogt GNM, Ogink J, Ponsioen B, Jalink K: A comparison of donor-acceptor pairs for genetically encoded FRET sensors: application to the Epac cAMP sensor as an example. PLoS ONE 2008, 3:e1916.
13. Shaner NC, Campbell RE, Steinbach PA, Giepmans BNG, Palmer AE, Tsien RY: Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat Biotechnol 2004, 22:1567-1572.
14. Yoshikawa S, Suzuki T, Watanabe M, Iseki M: Kinetic analysis of the activation of photoactivated adenylyl cyclase (PAC), a blue-light receptor for photomovements of Euglena. Photochem Photobiol Sci 2005, 4:727-731. 15. Olson EC: Onset of electrical excitability during a period of circus plasma
membrane movements in differentiating Xenopus neurons. J Neurosci 1996, 16:5117-5129.
16. Tramier M, Zahid M, Mevel J-C, Masse M-J, Coppey-Moisan M: Sensitivity of CFP/YFP and GFP/mCherry pairs to donor photobleaching on FRET determination by fluorescence lifetime imaging microscopy in living cells. Microsc Res Tech 2006, 69:933-939.
17. Stierl M, Stumpf P, Udwari D, Gueta R, Hagedorn R, Losi A, Gärtner W, Petereit L, Efetova M, Schwarzel M, Oertner TG, Nagel G, Hegemann P: Light modulation of cellular cAMP by a small bacterial photoactivated adenylyl cyclase, bPAC, of the soil bacterium Beggiatoa. J Biol Chem 2011, 286:1181-1188.
18. Klarenbeek JB, Goedhart J, Hink MA, Gadella TWJ, Jalink K: A mTurquoise-Based cAMP Sensor for Both FLIM and Ratiometric Read-Out Has Improved Dynamic Range. PLoS ONE 2011, 6:e19170.
doi:10.1186/1756-0500-4-241
Cite this article as: Hong et al.: Improved molecular toolkit for cAMP studies in live cells. BMC Research Notes 2011 4:241.
Submit your next manuscript to BioMed Central and take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit