HAL Id: tel-01617201
https://tel.archives-ouvertes.fr/tel-01617201
Submitted on 16 Oct 2017HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
l’enrichissement de l’environnement : rôle des
lymphocytes T
Hadi Zarif Peyvandi
To cite this version:
Hadi Zarif Peyvandi. Modulation de la plasticité hippocampique par l’enrichissement de l’environnement : rôle des lymphocytes T. Sciences agricoles. Université Côte d’Azur, 2017. Français. �NNT : 2017AZUR4058�. �tel-01617201�
Université de Nice Sophia Antipolis
Faculté des Sciences
Ecole Doctorale des Sciences de la Vie et de la Santé (ED85-SVS)
Disciplines : Interactions Moléculaires et CellulairesTHESE DE DOCTORAT
Modulation de la plasticité hippocampique par
l’enrichissement de l’environnement :
rôle des lymphocytes T
Thèse rédigée par
Hadi ZARIF
et co-encadrée par le Dr. Alice GUYON et le Dr. Agnès PETIT-PAITEL
Soutenue le 13 juillet 2017, devant le jury composé de :
Pr. Nicolas GLAICHENHAUS, PU, Université Côte d’Azur (UCA) Président Dr. José BOUCRAUT, MCU-PH, Université d’Aix-Marseille Rapporteur
Pr. Thomas FRERET, PU, Université de Caen-Normandie Rapporteur
Dr. Piotr BREGESTOVSKI, DR, Université d’Aix-Marseille Examinateur
Dr. Alice GUYON, DR, UCA Co-directrice de Thèse
Dr. Agnès PETIT-PAITEL, CR, UCA Co-directrice de Thèse
Thèse de Doctorat – Hadi ZARIF 2017
Thèse de Doctorat – Hadi ZARIF 2017 2
« I have no special talents.
I am only passionately curious. »
Thèse de Doctorat – Hadi ZARIF 2017
3
Ces travaux de recherche ont été menés au sein de :
Institut de Pharmacologie Moléculaire et Cellulaire (IPMC)
Dans l’équipe :
« Développement de stratégies thérapeutiques innovantes
pour le traitement de la dépression et de l’AVC »
Dirigée par
Dr. Catherine HEURTEAUX
Cette thèse a été financée par :
Thèse de Doctorat – Hadi ZARIF 2017
5
Remerciements
Je remercie chacun des membres du jury, qui m’ont fait l’honneur d’examiner et d’évaluer mes travaux de thèse. Je tiens notamment à remercier le Dr. Joseph Boucraut et le Dr. Thomas Freret, d’avoir accepté d’évaluer mon manuscrit.
Je voudrais remercier le Dr. Catherine Heurteaux, de m’avoir accueilli au sein de son laboratoire et de m’avoir donné tous les moyens nécessaires à mon épanouissement professionnel. Merci également au Labex ICST de m’avoir financé et soutenu.
Je remercie chaleureusement mes deux co-directrices de thèse, le Dr. Alice Guyon et le Dr. Agnès Petit-Paitel d’avoir grandement contribué à ma formation professionnelle et à mon développement personnel. Au même titre, je souhaite remercier le Dr. Joëlle Chabry pour sa contribution et son investissement dans mon projet de thèse.
Je remercie l’équipe du Pr. Nicolas Glaichenhaus, avec laquelle j’ai été en étroite collaboration, en particulier Mélanie Guyot et Anne Lazzari qui ont participé à ma formation sur des techniques d’immunologie.
Je remercie chacun des membres de l’équipe dans laquelle j’ai évolué (Catherine
Widmann, Carine Gandin, Nicolas Blondeau, Sarah Nicolas, Miled Bourourou, Marc Borsotto, Alaeddine Djillani, David-Emmanuel Dehut et Mariel Pietri) pour avoir participé
directement ou indirectement à ces travaux et contribué à mon épanouissement au sein du laboratoire.
Thèse de Doctorat – Hadi ZARIF 2017 6
un laboratoire de recherche, qui m’ont transmis leur passion et leur intérêt pour les sciences fondamentales, et avec qui j’ai eu de longues discussions au cours de ma thèse sur le fonctionnement du système immunologique.
Je remercie le personnel des différentes plateformes techniques avec qui l’ai eu l’opportunité de collaborer :
Julie Cazareth, responsable du tri cellulaire et de la cytométrie en flux ;
Frédéric Brau et Sophie Abelanet, responsables de l’imagerie et de la microscopie ; Marie-Jeanne Arguel, Kevin Lebrigand et Agnès Paquet, de la plateforme génomique ; Anne-Sophie Gay et Delphine Debayle, responsables de la plateforme protéomique.
Je remercie Estelle Ascoletti, Elisa Dourneau, Valentine Golzné et Maud Maillaut, les étudiantes dont j’ai eu l’opportunité d’encadrer durant ma thèse et qui ont apporté chacune leur contribution à ces travaux.
J’aimerai également remercier le responsable de l’animalerie Animex, Thomas Lorivel les animaliers, Lucien Relmy et Alain Barbot pour leur professionnalisme ainsi que Franck
Aguila pour les illustrations de ce manuscrit.
Enfin, je remercie tous mes proches, ma famille et mes amis, qui ont toujours eu, à mon égard, un soutien sans faille et des encouragements permanents, me permettant de puiser l’énergie et la motivation nécessaires dans l’accomplissement de ces travaux.
Thèse de Doctorat – Hadi ZARIF 2017
7
Résumé
INTRO : La plasticité cérébrale est une capacité remarquable des cellules du cerveau à
adapter leur structure et fonction en réponse à l’expérience et l’environnement. Cette plasticité cérébrale est favorisée par des conditions de vie favorables qui peuvent être modélisées chez le rongeur par le modèle de l’Environnement Enrichi (EE). L’EE consiste à mettre un grand nombre de souris dans de grandes cages comprenant de nombreux objets (nids, tunnels, roues…) qui sont changés régulièrement. L’EE induit une activité physique volontaire accrue, des conditions optimales pour la stimulation des interactions sociales, du comportement exploratoire et des fonctions cognitives. L’EE exerce des effets bénéfiques sur les processus physiologiques au niveau de nombreux systèmes (hormonal, cardiovasculaire, immunitaire…). L’EE réduit les comportements anxio-dépressifs, améliore l'apprentissage et la mémorisation. Ces effets sont sous-tendus par des changements au niveau du cerveau et en particulier de l’hippocampe, où l’on observe en EE plus de neurogenèse et synaptogenèse. De manière intéressante, chez les souris immunodéficientes, les performances mnésiques et la neurogenèse sont très altérées, suggérant une interaction bidirectionnelle entre le système immunitaire et le cerveau. Parmi les cellules du système immunitaire, les lymphocytes T (LT) semblent jouer un rôle particulièrement important dans les mécanismes de plasticité neuronale. Notre objectif a été de caractériser le rôle des LT dans les effets de l’EE sur la plasticité cérébrale et de chercher si ces effets impliquent une modification des LT par l’EE.
MATERIELS ET METHODES : Des souris âgées de 4 semaines ont été plusieurs
fois traitées soit avec des anticorps déplétants dirigés sélectivement contre les différentes sous-populations de LT afin de les éliminer soit avec des anticorps « isotypiques contrôles ». Les
Thèse de Doctorat – Hadi ZARIF 2017 8
protocole, plusieurs paramètres ont été analysés.
RESULTATS : Une première étude a été effectuée sur le rôle de l’hormone thyréotrope
(TRH) sur la plasticité montrant que la TRH est capable d’affecter la plasticité synaptique hippocampique par un blocage des récepteurs NMDA et de modifier l’établissement de la PLT.
Une deuxième étude vise à établir un modèle de l’EE et à mesurer différents effets de l’EE comme la neurogenèse hippocampique, la synaptogenèse et la plasticité synaptique au niveau des neurones glutamatergiques de l’hippocampe. Nous montrons que l’EE augmente la neurogenèse et la synaptogenèse, augmente la fréquence des courants post-synaptiques excitateurs spontanés et miniatures et diminue la PLT de la synapse CA3-CA1. Cette étude a désigné la durée d’hébergement de 4 semaines en EE comme étant celle où la plasticité de l’hippocampe est la plus notable.
Enfin, nous avons mené deux études en parallèle sur les effets de l’EE ; l’une sur le rôle
joué par les LT-CD8+ et l’autre par les LT-CD4+. Le rôle des LT-CD8+ dans les phénomènes
de plasticité cérébrale étant méconnu comparé aux LT-CD4+, nous avons mené une étude plus
approfondie sur le rôle des LT-CD8+. Globalement, nous montrons que plusieurs effets de l’EE
sont dépendants de la présence des LT. Premièrement, nous montrons que les effets bénéfiques de l’EE dans les tests de comportement visant à évaluer l’anxiété (Light & Dark, Openfield,
NSF) et la mémoire spatiale (Barnes maze) sont dépendants des LT-CD8+. De même,
l’augmentation du volume de l’hippocampe visible chez les souris hébergées en EE est inhibée chez les souris dont les LT-CD8+ ont été déplétés. Certains effets de l’EE que nous avons
étudiés sont communs entre les LT-CD4+ et les LT-CD8+. Ainsi, l’augmentation de la
neurogenèse hippocampique, de la synaptogenèse, de l’expression de gènes impliqués dans la plasticité synaptique (PSD95, Synapsine, Synaptophysine…) et l’activité électrophysiologique
Thèse de Doctorat – Hadi ZARIF 2017
9
l’aire CA1) sont affectées chez les souris dépourvues de LT-CD4+ ou LT-CD8+.
De manière intéressante, l’EE induit des modifications des LT issus de la rate et des
plexus choroïdes. En effet, le profil transcriptomique des LT-CD8+ et LT-CD4+ des souris en
EE est différent de celui des souris élevées en SE. Le pourcentage de LT-CD8+ « effector
memory » est augmenté dans le cerveau des souris élevées en EE alors que le pourcentage des
LT-CD8+ « central memory » est diminué. Après stimulation in vitro, les LT-CD8+ spléniques
des souris en EE prolifèrent plus rapidement et sécrètent plus de TNFa comparées aux souris en SE.
Afin de comprendre par quels mécanismes les LT affectent la plasticité cérébrale, nous avons focalisé nos recherches sur les plexus choroïdes, où sont localisés la majorité des LT dans le cerveau ; sur les cellules microgliales, pouvant constituer des intermédiaires entre LT et neurones, et sur le système thyroïdien, qui est affecté par la déplétion des LT.
CONCLUSION : Le système immunitaire et en particulier les LT sont impliqués dans
les effets bénéfiques d’un hébergement en EE, pouvant ainsi participer à plusieurs fonctions cognitives. Ces effets impliquent des modifications transcriptionnelles et fonctionnelles des LT du cerveau. Ces travaux novateurs mettent en évidence des interactions complexes entre cerveau et système immunitaire dans les conditions physiologiques et pathologiques.
Thèse de Doctorat – Hadi ZARIF 2017
11
Table des matières
Remerciements ... 5 Résumé ... 7 Table des matières ... 11 Liste des figures ... 14 Abréviations ... 15 Préface ... 17 La plasticité cérébrale post-natale ... 21 I. Définition ... 21 II. La plasticité neuronale ... 22 II.1. La neurogenèse adulte ... 23 II.2. La plasticité synaptique ... 25 II.3. La synaptogenèse ... 26 III. L’hippocampe : une structure cérébrale particulièrement « plastique » ... 27 III.1. Anatomie et structure ... 27 III.2. Les fonctions principales de l’hippocampe ... 29 III.3. La neurogenèse hippocampique ... 30 III.4. La plasticité synaptique hippocampique ... 31 Les acteurs participant à la plasticité cérébrale ... 34 I. Les acteurs cellulaires ... 34 I.1. La microglie ... 34 I.2. Les astrocytes ... 35 I.3. Les oligodendrocytes ... 36
Thèse de Doctorat – Hadi ZARIF 2017 12 III. Les conditions de vie et les facteurs environnementaux ... 57 L’enrichissement de l’environnement... 59 I. Le modèle murin et les effets bénéfiques chez le rongeur ... 59 I.1. Description du modèle murin d’EE ... 59 I.2. Les effets sur le système nerveux ... 60 I.2.a. Les effets comportementaux ... 61 I.2.b. Les effets sur la plasticité cérébrale ... 61 I.2.c. Les effets sur les neuropathologies ... 62 I.3. Les effets sur les fonctions physiologiques périphériques ... 63 II. Les mécanismes par lesquels l’EE peut affecter la plasticité cérébrale ... 85 Rôle des lymphocytes T (LT) dans les effets de l’EE ... 134 I. Système immunitaire et plasticité cérébrale ... 134 I.1. Interactions entre le système nerveux et le système immunitaire ... 134 I.1.a. Le système nerveux influence le système immunitaire ... 135 I.1.b. Le système immunitaire agit que le cerveau ... 137 II. Les lymphocytes T (LT) et la plasticité cérébrale induite par l’EE ... 138 II.1. Les LT : définition et fonctions primaires ... 138 II.2. Le rôle des LT dans la plasticité cérébrale ... 141 II.2.a. Le rôle des LT dans la cognition et la neurogenèse ... 141 II.2.b. Localisation et modes d’action des LT dans le cerveau ... 143 II.2.b.i. Dans le liquide cérébro-spinal (LCS) ... 145 II.2.b.ii. Au niveau des plexus choroïdes ... 145 II.2.b.iii. Au niveau des méninges ... 146 II.2.c. Comment les LT peuvent-ils affecter la plasticité cérébrale ? ... 146 Discussion ... 299 I. Introduction ... 299
Thèse de Doctorat – Hadi ZARIF 2017 13 III. Effets de l'EE sur la plasticité du cerveau. ... 300 IV. Mécanismes par lesquels l'EE affecte la plasticité cérébrale ... 305 IV.1. Rôle des LT dans les effets de l’EE ... 307 IV.1.a. La localisation des LT dans le cerveau ... 309 IV.1.b. Rôle des LT dans la neurogenèse ... 310 IV.1.c. Rôle des LT dans la plasticité synaptique ... 312 IV.2. Les effets de l’EE sur les LT ... 314
IV.3. Mécanismes d’action des LT-CD4+ dans les effets de l’EE ... 316
IV.4. Mécanismes d’action des LT-CD8+ dans les effets de l’EE ... 318
V. Application au bien-être animal et à l’Homme et les perspectives sociétales ... 318 Annexes ... 323 I. Le système nerveux ... 323 I.1. Définition ... 323 I.2. Neurone et neurotransmission ... 325 I.2.a. Le potentiel d’action ... 325 I.2.b. La synapse et la transmission synaptique ... 327 I.3. Enregistrement d’un champ électrique ... 330 I.3.a. Potentialisation à long terme (PLT) ... 331 I.4. Patch-clamp ... 331 Annexe II ... 335 Liste des publications ... 351 I. Articles publiés ... 351 II. Articles soumis ou en cours de révision ... 351 III. Articles en cours de rédaction ... 352 Références bibliographiques ... 353
Thèse de Doctorat – Hadi ZARIF 2017 14
Liste des figures
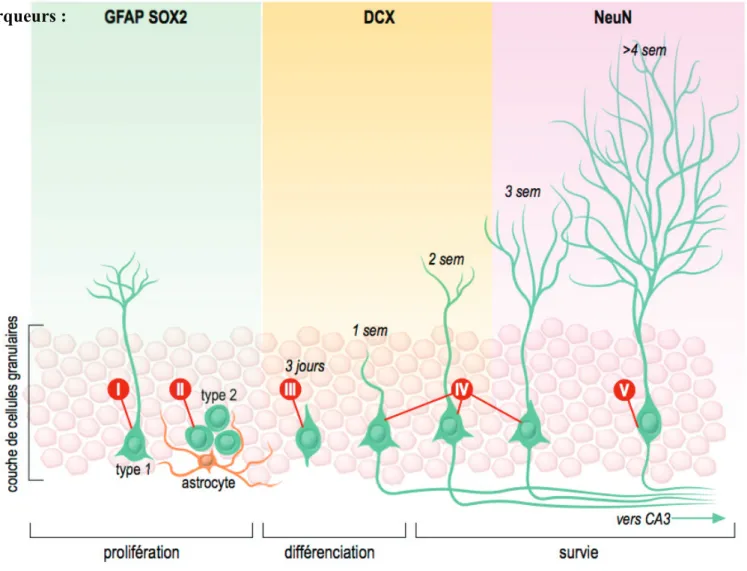
Figure 1 : Schéma représentant la maturation des neurones granulaires ... 24
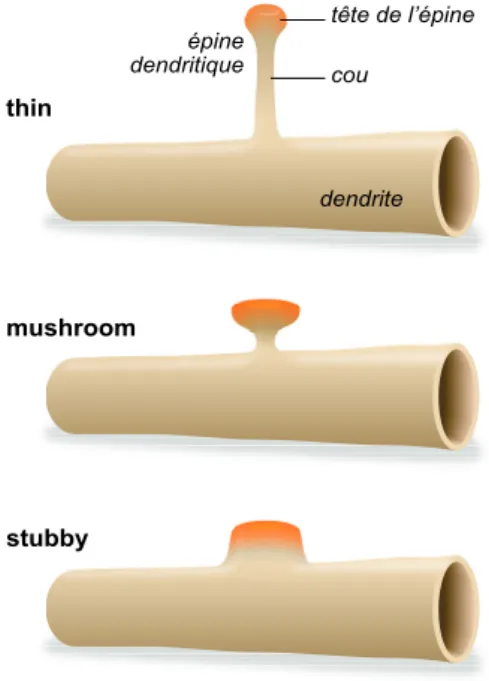
Figure 2 : Les changements morphologiques observés lors de la maturation d’une épine dendritique ... 25
Figure 3 : Organisation de l'hippocampe ... 28
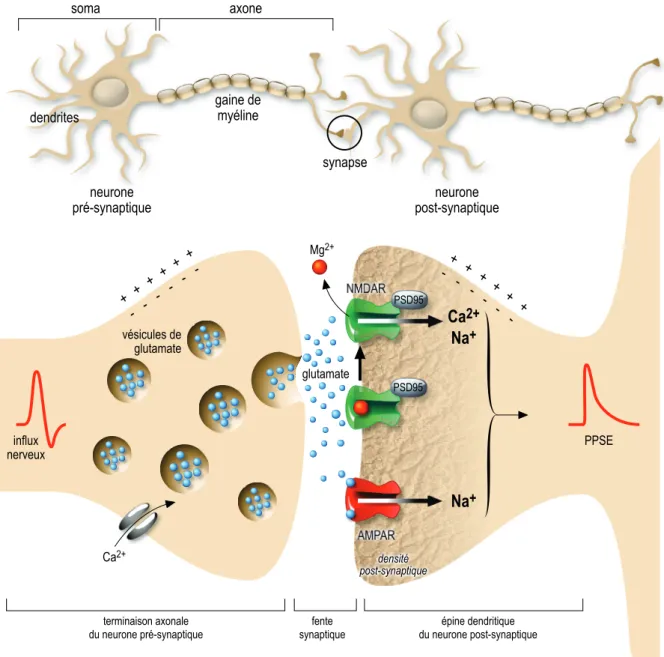
Figure 4 : Schéma d’une synapse chimique glutamatergique entre deux neurones ... 33
Figure 5 : Comparaison d’une cage standard d’élevage (SE – à gauche) ... 60
Figure 6 : Schéma représentant les différents marqueurs de surface des LT ... 141
Figure 7 : Les différents paramètres modulables dans le modèle de l'EE chez le rongeur (d'après Singhal et al., 2014) ... 303
Figure 8 : Schéma des différentes voies d'entrée des LT ... 324
Figure 9 : Les différentes étapes d’un potentiel d’action neuronal ... 326
Figure 10 : Schéma d'une synapse chimique ... 328
Figure 11 : Schéma des différentes configurations de patch-clamp ... 333
Figure 12 : Schéma représentant la méthode d'enregistrement d'un courant évoqué au niveau d'un neurone pyramidale dans la région CA1 de l'hippocampe, suite à une stimulation des collatérales de Schaffer ... 334
Tableau 1 : Effets bénéfiques de l’EE sur les pathologies cérébrales ... 63
Tableau 2 : Les différentes sous-populations de LT-CD4+ ... 139
Tableau 3 : Récapitulatif des effets de l'EE et de la déplétion des LT (CD8+ et CD4+) sur les différents paramètres mesurés ... 298
Thèse de Doctorat – Hadi ZARIF 2017
15
Abréviations
Ac Anticorps
ACTH Hormone corticotrope ou adénocorticotrope
ADN Acide désoxyribonucléique
AMPA Acide α-amino-3-hydroxy-5-méthyl-4-isoxazolepropionique
AMPAR Récepteurs AMPA
ApN Adiponectine
APV Acide 2-amino-5-phosphonovalérique
ARN Acide ribonucléique
AVC Accident vasculaire cérébral
BDNF Facteur neurotrophique dérivé du cerveau (brain derived neurotrophic factor)
BHE Barrière hémato-encéphalique
BrdU Bromodéoxyuridine
CA Corne d'Ammon
CD Cluster de différenciation
CFSE Carboxyfluorescin diacetate succinidyl ester
CNQX 6-cyano-7-nitroquinoxaline-2,3-dione
CNTF Facteur neurotrophique cliliaire (ciliary neurotrophic factor)
CPA Cellule présentatrice d’antigène
CPSE Courants post-synaptiques excitateurs
DAB 3,3'-diaminobenzidine
DLT Dépression à long terme
EE Environnement enrichi
GABA Acide γ-aminobutyrique
GD Gyrus denté
GH Hormone de croissance (growth hormone)
GR Récepteurs des glucocorticoïdes
HT Hormones thyroïdiennes
IFN Interféron
icv Intra-cérébro-ventriculaire
Thèse de Doctorat – Hadi ZARIF 2017 16 IL Interleukine ip Intrapéritonéal LCS Liquide cérébro-spinal LPS Lipopolysaccharide LT Lymphocytes T
CMH Complexe majeur d’histocompatibilité
NGF Facteur de croissance des nerfs (nerve growth factor)
NK Tueuse naturelle (natural killer)
NMDA Acide N-méthyl-D-aspartique
NMDAR Récepteurs NMDA
NTF Neurotrophine
PA Potentiel d’action
PLT Potentialisation à long terme
PPSE Potentiels post-synaptiques excitateurs
PSD Protéine de la densité postsynaptique (postsynaptic density protein)
QX314 N-(2,6-dimethylphenylcarbamoylmethyl)triethylammonium bromide
SCID Immunodéficience sévère combinée (severe combined immune deficiency)
SE Environnement standard (strandard environment)
SEM Erreur type de la moyenne (Standard Error of the Mean)
SHF Stimulation à haute fréquence
TLR Récepteur de type Toll (Toll-like receptor)
TNF Facteur de nécrose tumorale (tumor necrosis factor)
TRH Thyréolibérine ou hormone thyréotrope (thyrotropin-releasing hormone)
TSH Thyréostimuline (thyroid stimulating hormone)
TTR Transthyrétine
TTX Tétrodotoxine
VEGF Facteur de croissance de l’endothélium vasculaire (vascular endothelial growth factor)
WT Sauvage (wild type)
Thèse de Doctorat – Hadi ZARIF 2017
17
Préface
Durant mon cursus en Sciences de la Vie et de la Santé, deux domaines de la biologie m’ont fortement intéressé : d’une part la neurobiologie en raison de la complexité du système nerveux et d’autre part l’immunologie. Au cours de mes travaux de Doctorat, j’ai pu allier ces deux domaines puisque j’ai eu l’opportunité de l’effectuer dans un laboratoire de recherche ayant comme axe thématique la neuro-immunologie.
Historiquement, le laboratoire d’accueil dans lequel j’ai effectué mes travaux de thèse, a étudié la neuro-inflammation dans des conditions physiologiques et pathologiques. Dans ce cadre, une caractérisation exhaustive des différentes sous-populations de cellules immunes présentes dans le cerveau a eu lieu selon un protocole bien établi dans la littérature (Cardona et al., 2006). Brièvement, les souris saines ont été perfusées pour éliminer les cellules sanguines, le cerveau a été disséqué et mis en suspension cellulaire. Grâce à l’utilisation de différentes combinaisons d’anticorps dirigés contre des marqueurs de surface, ils sont parvenus à discriminer plusieurs types de cellules dans le cerveau parmi lesquels les lymphocytes T. En s’intéressant de plus près à la littérature, ils se sont rendus compte que d’autres équipes avaient également décrit dans des conditions physiologiques la présence de lymphocytes T au niveau cérébral et leur attribuaient un rôle potentiel dans la plasticité cérébrale (Kipnis et al., 2004b).
En 2010, les travaux du Pr. Cao démontraient des effets bénéfiques d’un enrichissement des conditions d’hébergement (modèle de l’environnement enrichi) sur l’évolution de différents cancers (côlon et mélanomes). Les mécanismes sous-jacents désignaient un rôle essentiel de l’interaction entre le système nerveux et le système immunitaire (Cao et al., 2010). En
Thèse de Doctorat – Hadi ZARIF 2017 18
intéressé de plus près au modèle de l’environnement enrichi et s’est fixé comme projet d’étude les effets de ce modèle sur la plasticité cérébrale et la neuro-immunité tandis que l’équipe du Pr. Nicolas Glaichenhaus étudiait les effets sur le système immunitaire en périphérie.
De cette collaboration est né mon projet de thèse. Etant donné que les lymphocytes T étaient décrits pour jouer un rôle dans la plasticité cérébrale et que le modèle de l’environnement enrichi agit à la fois sur la plasticité cérébrale et sur le système immunitaire je me suis intéressé à étudier le rôle des lymphocytes T dans les effets de l’environnement enrichi.
J’ai commencé mon projet en participant à une étude sur un modèle murin visant à améliorer la plasticité cérébrale chez l’adulte par l’intermédiaire d’un enrichissement de leur environnement et de leur milieu d’hébergement. Par la suite, je me suis concentré à définir le rôle que pourraient jouer des lymphocytes T dans ce modèle où la plasticité cérébrale est optimale.
Dans ce manuscrit, j’introduirai dans un premier temps la notion de plasticité cérébrale chez l’adulte. Ainsi, les mécanismes et les différentes notions de plasticité seront abordés. Dans ce contexte, je présenterai un premier article scientifique pour lequel je suis premier auteur qui évalue le rôle d’une hormone, la TRH, sur la plasticité d’une région du cerveau qui est l’hippocampe.
Dans un second temps, je détaillerai le modèle de l’environnement enrichi, dans lequel les souris peuvent expérimenter de nombreuses stimulations sensorimotrices favorisant la plasticité du système nerveux central, plasticité qui peut être observée à différentes échelles.
Thèse de Doctorat – Hadi ZARIF 2017
19
l’établissement de ce modèle et l’autre une étude sur la neuro-inflammation.
Puis, j’exposerai le rôle que peuvent jouer les lymphocytes T dans les phénomènes de plasticité cérébrale. Je présenterai deux derniers articles (en cours de soumission) dont je suis le premier auteur, dans lesquels est abordé le rôle des lymphocytes T sur les effets du modèle de l’environnement enrichi et les mécanismes mis en jeu.
Enfin je finirai par une discussion générale sur mes travaux de thèse.
Thèse de Doctorat – Hadi ZARIF 2017
21
La plasticité cérébrale post-natale
Durant le développement intra-utérin, le cerveau se forme après plusieurs étapes allant de la formation de nouveaux neurones à la myélinisation des fibres en passant par la création de nouvelles connexions intercellulaires. Après la naissance, le système nerveux continue à se développer, avec des réarrangements des circuits neuronaux, durant des périodes critiques comme la puberté par exemple. Chez l’adulte, la maturation du système nerveux est terminée d’un point de vue développemental mais le cerveau continue à réarranger ses circuits par l’intermédiaire des différentes étapes mentionnées plus haut, mais cette fois à des fins d’adaptation à l’environnement. C’est la plasticité post-natale qui m’a intéressé durant ma thèse et que je développerai durant ce manuscrit.
I. Définition
Le terme de plasticité est la qualité de ce qui est « malléable », « souple » et « modifiable ». En 1949, Donald Hebb postulait sa théorie sur la manière dont les neurones sont connectés entre eux afin de former des engrammes, traces biologiques de la mémoire. Cependant, cette théorie était peu appuyée par des preuves expérimentales excepté les études d’Eric Kandel sur l’aplysie. Il a fallu attendre les années 1960 et les travaux de Mark Rosenzweig et ses collègues David Krech, Edward Bennett et Marian Diamond sur le rongeur pour avoir la démonstration que le cerveau adulte est anatomiquement et chimiquement soumis à un réarrangement permanent de ses réseaux neuronaux. La plasticité cérébrale est alors définie comme la capacité des neurones et des circuits neuronaux à changer, structurellement et fonctionnellement, en réponse à l’expérience et au vécu. C’est une propriété fondamentale dans l’adaptabilité de notre comportement au cours du développement, des processus d’apprentissage et de mémorisation du cerveau ainsi que de la régénérescence post-lésionnelle. A travers le vécu, l’expérience est traduite en patterns d’activités électriques dans les circuits
Thèse de Doctorat – Hadi ZARIF 2017 22
fonctionnelles et structurelles de plasticité à travers la coordination, dans le temps et dans l’espace, de l’action de cellules et molécules spécifiques. La plasticité peut affecter le niveau d’efficacité des réseaux neuronaux : les renforcer, les défaire ou les réorganiser. Tous ces changements peuvent se faire au niveau cellulaire, avec la formation de nouveaux neurones (neurogenèse), au niveau de l’arborisation dendritique et/ou axonale mais également au niveau des synapses (plasticité synaptique).
La plasticité cérébrale est un phénomène observé durant le développement et chez l’adulte. Les mécanismes impliqués semblent être similaires au cours de la vie. Cependant il y a une grande différence de plasticité entre un cerveau en développement, très plastique, et un cerveau adulte, beaucoup moins plastique (Hensch, 2005). Le cortex visuel a été le premier modèle d’étude dans la plasticité dépendante de l’expérience. En effet, Wiesel et Hubel, qui ont obtenu le prix Nobel, ont démontré que la privation de la vision sur un œil engendrait des changements et des réarrangements dans le cortex visuel en faveur de l’œil non privé (Wiesel & Hubel, 1963a; Wiesel & Hubel, 1963b). Depuis cette étude, de nombreuses équipes de recherche ont démontré l’existence de plasticité dans plusieurs autres structures cérébrales dans de nombreuses conditions expérimentales (Cerebral Plasticity, New perspectives, 2011, Chalupa, Bernardi et al., https://mitpress.mit.edu/books/cerebral-plasticity).
II. La plasticité neuronale
La plasticité neuronale ou la neuroplasticité correspond à toutes les modifications cellulaires et/ou synaptiques des neurones. Ainsi, la formation de nouveaux neurones, autrement appelée neurogenèse, les modifications de la force des synapses et la création de nouvelles synapses, appelée la synaptogenèse font partie de la plasticité neuronale.
Thèse de Doctorat – Hadi ZARIF 2017
23
La neurogenèse adulte
Jusqu’au milieu du XXème siècle on considérait le cerveau adulte comme incapable de
renouveler ses cellules nerveuses. Il a fallu attendre 1958 pour qu’une équipe découvre la mitose et la synthèse d’ADN dans le cerveau de souris adultes (Messier et al., 1958). Plus tard, plusieurs études montrent que de nouvelles cellules sont générées dans le système nerveux adulte chez le canari (Alvarez-Buylla et al., 1998; Paton & Nottebohm, 1984) et d’autres oiseaux adultes (Alvarez-Buylla et al., 1990), le poisson zèbre (Zupanc et al., 2005), les cnidaires (éponges), insectes (grillons) (Cayre et al., 2007), souris, rats (Altman & Das, 1965; Kuhn et al., 1996) et autres mammifères (Lois & Alvarez-Buylla, 1993) dont l’homme (Eriksson et al., 1998; Gross, 2000; Kempermann et al., 2015). Du bromodéoxyuridine (BrdU), un nucléoside synthétique, analogue structurel de la thymine, incorporé dans le cerveau de patients atteints de tumeurs cérébrales, a permis de montrer la multiplication des cellules nerveuses au niveau de l’hippocampe (voir chapitre neurogenèse hippocampique page 30).
De la neurogenèse adulte est également observée au niveau de la zone sous-ventriculaire (ZSV). Contrairement à la neurogenèse hippocampique, les cellules néoformées de la ZSV migrent au niveau du bulbe olfactif (au-dessus de la cavité nasale chez l’homme). Au niveau du bulbe olfactif, les cellules dégénèrent si elles ne sont pas intégrées aux réseaux neuronaux. Ce renouvellement cellulaire pourrait jouer un rôle dans la perception et le codage de nouvelles odeurs. Cependant, chez l’homme, moins d’1% des neurones sont renouvelés tous les 100 ans (Bergmann et al., 2012). De la neurogenèse a été décrite également dans l’hypothalamus chez le rongeur (Yuan & Arias-Carrion, 2011).
La formation de nouveaux neurones chez l'adulte est un processus complexe, qui nécessite une série d'étapes : division asymétrique des cellules souches neuronales, migration, différenciation en neurones granulaires et intégration dans les réseaux neuronaux. Enfin, de la neurogenèse adulte est également décrite suite à une mort neuronale anormale. C’est le cas dans
Thèse de Doctorat – Hadi ZARIF 2017 24
les ischémies cérébrales (Arvidsson et al., 2002). Cependant, dans le cadre de ma thèse je me suis essentiellement focalisé sur la neurogenèse adulte dans des conditions physiologiques et pathologiques en lien avec des perturbations du système immunitaire (déplétion des LT).
Figure 1 : Schéma représentant la maturation des neurones granulaires au niveau du gyrus denté de l’hippocampe
Thèse de Doctorat – Hadi ZARIF 2017
25
La plasticité synaptique
La plasticité au niveau des synapses, autrement appelée plasticité synaptique, correspond à toutes les modifications morphologiques et/ou fonctionnelles survenant au niveau des synapses. Les synapses évoluent avec le temps. Certaines sont supprimées, d’autres se créent (synaptogenèse), mais toutes se modifient avec le temps et tendent, soit à se renforcer, soit à affaiblir les communications entre les neurones selon les besoins. Donald Hebb postulait que « lorsqu’un neurone reçoit de façon répétée des afférences d’un autre neurone l’efficacité des connexions entre les deux neurones est augmentée » désignant la plasticité synaptique à la base des processus d’apprentissage et de mémorisation. Les modifications de l’activité synaptique peuvent être sous-tendues par des changements au niveau de la quantité de neurotransmetteurs libérée, de l’expression des récepteurs aux neurotransmetteurs ou des protéines d’ancrage et peut également s’accompagner de changements morphologiques de l’épine (stubby : pas de cou, thin : long cou fin avec une petite tête, mushroom : en forme de « champignon », considérée comme appartenant à une synapse mature) (Hering & Sheng, 2001; Rochefort & Konnerth, 2012).
Figure 2 : Les changements morphologiques observés lors de la maturation d’une épine dendritique thin mushroom stubby dendrite tête de l’épine cou épine dendritique
Thèse de Doctorat – Hadi ZARIF 2017 26
La synaptogenèse
La synaptogenèse, qui correspond à la formation de nouvelles synapses, constitue un aspect très important dans le développement cérébral et la régulation des fonctions cognitives comme l’apprentissage et la mémoire (Roberts et al., 2010; Tschida & Mooney, 2012; Xu et al., 2009). Elle est intense durant le développement et diminue avec l’âge mais se produit tout au long de la vie (Coggan et al., 2004; Gan et al., 2003; Rosenzweig & Barnes, 2003). La formation d’une synapse implique des mécanismes très complexes. Le premier contact de l’axone du neurone pré-synaptique avec les épines dendritiques du neurone post-synaptique initie une série d’interactions impliquant un échange complexe de signaux entre les deux neurones qui mènera à la formation de la synapse. La formation et la maturation d’une nouvelle synapse nécessite à la fois une maturation de l’arborisation axonale du neurone pré-synaptique et de la membrane du neurone post-synaptique. La première étape se fait grâce à un gradient moléculaire et à la sécrétion de facteurs, notamment par les cellules gliales proches, pour guider les prolongements axonaux du neurone pré-synaptique afin de trouver les prolongements dendritiques du neurone post-synaptique. La maturation consiste en la formation et l’accumulation de vésicules pré-synaptiques, à la formation de densités sous membranaires
post-synaptiques (PSD), et à l’augmentation du nombre de canaux Ca2+ transmembranaires.
Potentiellement, plus la tête de l’épine dendritique est grosse plus le nombre de récepteurs et la surface de libération de neurotransmetteurs sont grands. Une fois la connexion synaptique établie, les interactions entre les éléments pré et post-synaptiques, des facteurs neurotrophiques comme le BDNF, libérés en faibles quantités par les cellules cibles, permettent de contrôler la différenciation, la croissance et la survie de la synapse (Garner et al., 2002; Vaughn, 1989; Waites et al., 2005).
Plusieurs régions du cerveau présentent une plasticité. Cependant, pendant mes travaux de thèse, je me suis intéressé à une structure cérébrale présentant une plasticité remarquable en
Thèse de Doctorat – Hadi ZARIF 2017
27
arguments. Tout d’abord, l’hippocampe est une structure centrale dans plusieurs processus cognitifs du cerveau. Il est en étroite communication avec plusieurs autres régions du cerveau comme le cortex préfrontal, l’hypothalamus ou encore l’amygdale. La structure de l’hippocampe est très organisée et très étudiée, et par conséquent les outils d’études et les connaissances sont abondants dans la littérature. Enfin, l’hippocampe joue un rôle primordial dans les phénomènes d’apprentissage et de mémorisation, mais aussi dans les émotions, et une modification de la plasticité neuronale dans cette région serait directement corrélée à des modifications de comportement des rongeurs dans les tests de tâches mnésiques, ou des tests d’anxiété, par exemple.
III.
L’hippocampe : une structure cérébrale particulièrement
« plastique »
III.1.
Anatomie et structure
Chez l’homme, l’hippocampe fait partie du système limbique et se situe au niveau de la cinquième circonvolution temporale, sous le cortex, dans le lobe temporal. Il se compose principalement de trois sous-régions : le gyrus denté (GD), la corne d’Ammon (divisée en trois régions : CA1, CA2 et CA3) et le subiculum. Les informations entrant dans l’hippocampe passent par la voie perforante correspondant aux projections du cortex entorhinal sur les cellules granulaires du GD (couche granulaire). Les neurones granulaires du GD envoient alors des axones, les fibres moussues, faire synapses sur les neurones pyramidaux de l'aire CA3 qui innervent à leur tour les cellules pyramidales de CA1 par l'intermédiaire des collatérales de Schaffer. Ce réseau de connexions forme le circuit trisynaptique de l’hippocampe (Amaral & Witter, 1989; Andersen et al., 1971). Chacune de ces synapses est excitatrice (libération du
Thèse de Doctorat – Hadi ZARIF 2017 28
sortent alors par le subiculum et sont dirigées vers les différentes aires du néocortex.
L’organisation extrêmement structurée en couches des corps cellulaires des différents neurones de l’hippocampe est un réel atout pour les études électrophysiologiques (voir Annexe).
Figure 3 : Organisation de l'hippocampe
A. Coupe sagittale de cerveau de souris adulte. L’hippocampe est anatomiquement et fonctionnellement considéré comme un vrai carrefour cérébral, recevant des fibres de l’hypothalamus et du thalamus et projetant vers le cortex.
B. Coupe transversale de l’hippocampe observée par microscopie optique. Son organisation caractéristique, avec le gyrus denté (GD) et les différentes régions (CA3 et CA1) est clairement visible.
C. Circuit trisynaptique de l’hippocampe. La 1ère synapse est constituée des fibres de la voie perforante qui se terminent sur les cellules en grain du gyrus denté (GD). La 2ème synapse est établie par les fibres moussues (FM), entre les axones des cellules du GD et les dendrites des cellules pyramidales de CA3. Enfin la 3ème synapse est établie par les collatérales de Schaffer (CdS) entre les cellules de CA3 et CA1.
cortex mésencéphale cervelet bulbe rachidien pont thalamus hypothalamus striatum bulbe olfactif hippocampe CA1 CA3 GD CdS FM GD CA1 CA3 efférences voie perforante
Thèse de Doctorat – Hadi ZARIF 2017
29
Les fonctions principales de l’hippocampe
L'hippocampe joue un rôle central dans l’apprentissage, la mémoire, la navigation spatiale. Il participe également à la régulation de l’humeur et des émotions, à l’acquisition des connaissances et à l’adaptation d’un individu à son environnement (Opitz, 2014).
En 1957, William Scoville et Brenda Milner publient leurs observations sur le patient HM. Ils décrivent que des dommages hippocampiques bilatéraux entraînent une amnésie antérograde sévère (difficultés à former de nouveaux souvenirs) associée à une amnésie rétrograde partielle (Scoville & Milner, 1957), ce qui suggère l’implication de l’hippocampe dans l’acquisition de nouvelles informations et dans la consolidation des souvenirs (Chozick, 1983). La taille de l’hippocampe est positivement corrélée à la capacité de mémorisation chez différentes espèces (Jacobs, 2003) et à l’enrichissement des conditions d’hébergement chez le rongeur (Huttenrauch et al., 2016). Une diminution du volume hippocampique est observée avec le vieillissement (Schuff et al., 1999) et dans le cas de dépressions sévères (Videbech & Ravnkilde, 2004), mais peut être évitée par la pratique d’une activité physique régulière (Erickson et al., 2011). En 1971, John O’Keefe et Lynn Nadel découvrent que des neurones spécifiques s’activent au niveau de l’hippocampe lorsque le rat se déplace dans un environnement bien précis. Ces cellules sont appelées les cellules de lieu et ces travaux mettent en lumière le rôle de l’hippocampe dans le repérage et le codage de l’information spatiale appelé mémoire spatiale (O'Keefe & Dostrovsky, 1971).
Des études ont démontré que la partie dorsale (postérieure) de l’hippocampe est plutôt impliquée dans les fonctions cognitives comme l’apprentissage et la mémoire spatiale alors que la partie ventrale (antérieure) est plutôt impliquée dans les émotions et le stress (Fanselow & Dong, 2010).
Thèse de Doctorat – Hadi ZARIF 2017 30
La neurogenèse hippocampique
De nouvelles cellules nerveuses sont produites chaque jour au niveau du GD (Shors et al., 2012). Cette neurogenèse perdure tout au long de la vie, et diminue avec l'âge (Kempermann et al., 2002).
Le rôle fonctionnel de la neurogenèse n’est pas encore complètement établi. Elle aurait un possible rôle dans le contrôle de l’humeur (Malberg et al., 2000), l’apprentissage et la mémoire spatiale (Deng et al., 2010). Plusieurs facteurs peuvent moduler la neurogenèse. Le
vieillissement (Kuhn et al., 1996), le manque de sommeil (Hairston et al., 2005), le stress
(Gould et al., 1997), l’exposition aux glucocorticoïdes (Nicolas et al., 2015) et certains neurotransmetteurs comme le GABA (Tozuka et al., 2005) tendent à diminuer la neurogenèse. A l’inverse, les facteurs neurotrophiques tels que le BDNF (Lee et al., 2002), le CNTF (Chojnacki et al., 2003; Emsley & Hagg, 2003), l’IGF1 (Aberg et al., 2003), l’activité cognitive et l’activité physique (Brown et al., 2003; Curlik et al., 2013; Curlik & Shors, 2013; Kempermann et al., 1997b) et certains neurotransmetteurs comme le glutamate (Bai et al., 2003) ou la sérotonine (Banasr et al., 2004; Brezun & Daszuta, 1999) peuvent influencer positivement la neurogenèse. Certains cytokines comme l’IL-1b (Chen et al., 2013), l’IL-6 (Bowen et al., 2011; Monje et al., 2003), l’IL-10 (Perez-Asensio et al., 2013), l’IFNg (Johansson et al., 2008b) ainsi que le TNFa (Monje et al., 2003) peuvent affecter selon leur concentration, positivement ou négativement, la neurogenèse au niveau du GD (Borsini et al., 2015).
Outre ces nombreux facteurs, les hormones comme les hormones thyroïdiennes (Desouza et al., 2005) ou la consommation de certains médicaments tels que les antidépresseurs (Duman et al., 2001; Malberg et al., 2000) ou drogues peuvent également moduler la neurogenèse hippocampique (Lledo et al., 2006).
Thèse de Doctorat – Hadi ZARIF 2017
31
La plasticité synaptique hippocampique
L’électrophysiologie sur des tranches de cerveaux est un réel atout dans l’étude de la plasticité de l’hippocampe car elle permet entre autres de mesurer la force des synases. Dans le cas de l’hippocampe, les neurones excitateurs sont principalement glutamatergiques, c’est-à-dire que le neurotransmetteur excitateur est le glutamate. Le glutamate se fixe sur ses récepteurs présents sur le neurone post-synaptique qui sont des récepteurs-canaux ou ionotropiques AMPA (AMPAR, perméables aux ions sodium) et NMDA (NMDAR, perméables aux ions sodium et calcium, normalement bloqués par les ions magnésium au potentiel de repos). Le glutamate peut également se fixer sur des récepteurs métabotropiques mGluR couplés aux protéines G, qui sont souvent pré-synaptiques. Au niveau post-synaptique, l’entrée des cations va engendrer des modifications au niveau du neurone post-synaptique et induire un changement du potentiel de membrane dans le sens d’une dépolarisation. Cette dépolarisation peut être enregistrée sous forme d’un potentiel post-synaptique excitateur (PPSE). Si la sommation spatiale et temporelle des PPSE permet d’atteindre le seuil de déclenchement des potentiels d’action au niveau du segment initial de l’axone, un potentiel d’action est généré et se propage à son tour jusqu’à la terminaison axonale suivante.
La plasticité des réseaux de l’hippocampe se manifeste au niveau de la formation de nouvelles synapses (synaptogenèse). Il est décrit qu’une tâche mnésique ou la pratique régulière d’un exercice physique volontaire modifie la synaptogenèse hippocampique en augmentant le nombre d’épines dendritiques et la taille des synapses sur certaines populations neuronales (Ambrogini et al., 2013; Mahmmoud et al., 2015). La plasticité se manifeste également au niveau fonctionnel (vérifiable par des études électrophysiologiques) : les synapses de l’hippocampe présentent plusieurs formes de plasticité fonctionnelle. Expérimentalement, on a ainsi mis en évidence une plasticité à court terme (« paired pulse facilitation ou depression ») (quelques millisecondes), une plasticité à moyen terme (secondes) et une plasticité à long terme
Thèse de Doctorat – Hadi ZARIF 2017 32
correspond à une modification prolongée de la force synaptique dans le sens d’une diminution (Dépression à Long Terme ou DLT) ou d’une augmentation (Potentialisation à Long Terme, PLT). La PLT peut être provoquée de façon expérimentale par une stimulation à haute fréquence (SHF) des collatérales de Schaffer (synapses CA3-CA1). Classiquement sur des tranches d’hippocampe maintenues en survie, on effectue une stimulation basale des neurones pré-synaptiques (axones des neurones CA3), et on enregistre la réponse post-synaptique engendrée (au niveau des neurones CA1), appelée réponse évoquée, soit en patch-clamp dans la configuration cellule entière (la réponse est alors un Potentiel ou un Courant Post-Synaptique Excitateur évoqué, ou ePPSE ou eCPSE), soit avec des électrodes de champ (la réponse enregistrée est alors une réponse d’ensemble appelée PPSE de champ ou « field potential », fPPSE). En l’absence de toute stimulation expérimentale, on peut observer une libération basale de neurotransmetteur sous la forme de PPSE spontanés (sPPSE). Même en présence de TTX, lorsque les PA sont bloqués, on observe toujours une petite libération de glutamate probablement due à une fluctuation de la concentration de calcium dans les terminaisons. Les PPSE alors observés sont appelés PPSE miniatures (mPPSE).
Suite à une stimulation à haute fréquence des collatérales de Schaffer, les ePPSEs ou les fPPSEs induits par le stimulus de base deviennent plus amples (potentialisation) sur une longue durée (à long terme). La PLT peut reposer sur des modifications à la fois pré (libération accrue de glutamate) et post-synaptiques (modification des AMPAR et NMDAR). Au niveau de la synapse CA3-CA1, la SHF dépolarise fortement le neurone post-synaptique ce qui a pour effet
de lever le blocage des NMDAR par des ions magnésium Mg2+. Ceci conduit à l’entrée du
calcium extracellulaire dans la cellule, second messager qui induit une cascade de modifications intracellulaires. A court terme (secondes), ces modifications se traduisent par la phosphorylation des AMPAR, les rendant plus sensibles au glutamate. A moyen terme
Thèse de Doctorat – Hadi ZARIF 2017
33
neurone
pré-synaptique post-synaptiqueneurone synapse
dendrites gaine demyéline
soma axone vésicules de glutamate influx nerveux Ca2+ NMDAR Mg2+ PSD95 PSD95 PPSE Ca2+ Na+ Na+ terminaison axonale
du neurone pré-synaptique synaptiquefente du neurone post-synaptiqueépine dendritique + -+ -+ -+ -+ -+ - - ++- + - +- + - - + densité post-synaptique glutamate
plus long terme (heures et plus), une augmentation de la biosynthèse de nouveaux AMPAR est observée (Bennett, 2000).
Figure 4 : Schéma d’une synapse chimique glutamatergique entre deux neurones
La plasticité au niveau des neurones est finement orchestrée par plusieurs facteurs endogènes et environnementaux.
Thèse de Doctorat – Hadi ZARIF 2017 34
Les acteurs participant à la plasticité cérébrale
I. Les acteurs cellulaires
Plusieurs types de cellules influent directement et/ou indirectement sur la plasticité neuronale. Je détaille dans ce chapitre, de manière non exhaustive, le rôle que peuvent jouer certaines cellules gliales sur la plasticité neuronale.
I.1.
La microglie
Les cellules microgliales, sont des cellules dynamiques et en interaction permanente avec leur environnement et notamment les neurones (Davalos et al., 2005; Nimmerjahn et al., 2005; Stence et al., 2001). Elles présentent des pseudopodes qui sont des prolongements longs et ramifiés permettant leur déplacement et les interactions avec leur environnement. Au niveau du cerveau, la microglie, en collaboration avec les phagocytes polynucléés (neutrophiles, macrophages périvasculaires, macrophages au niveau des plexus choroïdes, macrophages des méninges, macrophages infiltrants provenant des monocytes circulants) participe à la réponse immunitaire et la neuro-inflammation (Guillemin & Brew, 2004). Par ailleurs, de récents travaux démontrent que, dans des conditions non pathologiques, ces cellules sont capables de réguler la plasticité cérébrale (Delpech et al., 2015; Morris et al., 2013). Les cellules microgliales sont décrites pour jouer un rôle important dans le développement et l’activité neuronale, le remodelage de la structure des neurones et la modulation de la plasticité synaptique (Blais & Rivest, 2003; Hanisch, 2013; Kettenmann et al., 2011; Kettenmann et al., 2013). La microglie interagit avec les neurones à travers l’expression de neurotransmetteurs excitateurs et inhibiteurs ainsi que de leur récepteurs (Kettenmann et al., 2011; Morris et al., 2013; Pocock & Kettenmann, 2007) et la sécrétion de cytokines (Hanisch, 2002; Mallat & Chamak, 1994; Streit & Xue, 2012).
Thèse de Doctorat – Hadi ZARIF 2017
35
phagocytose et élimination du matériel synaptique immature et/ou non fonctionnel. Dans l’hippocampe, ce mécanisme se fait sous le contrôle de la fractalkine (CX3CL1) dont le récepteur, CX3CR1 est exclusivement exprimé par la microglie (Maggi et al., 2011; Paolicelli et al., 2011; Schafer et al., 2012). Une autre voie d’action pourrait être l’interaction entre CD200, exprimé à la surface des neurones, et son récepteur CD200R exprimé sur la surface des cellules microgliales (Barclay et al., 2002; Costello et al., 2011; Hoek et al., 2000; Walker et al., 2009). La microglie participe à la synaptogenèse en sécrétant une cytokine anti-inflammatoire, l’IL-10, dont le récepteur est exprimé sur les neurones hippocampiques (Lim et al., 2013). La microglie sécrète l’IL-1b, l’IL-6 et le TNFa qui peuvent agir sur l’activité synaptique, moduler la plasticité cérébrale et réguler des processus cognitifs comme l’apprentissage et la mémorisation (Blank & Prinz, 2013; Boulanger, 2009; Delpech et al., 2015; Goshen et al., 2007; Yirmiya & Goshen, 2011).
L’action de la microglie sur la plasticité neuronale peut également se faire à travers l’expression et la sécrétion du BDNF (Parkhurst et al., 2013), un facteur neurotrophique, liant sont récepteur neuronal TrkB (Ferrini & De Koninck, 2013; Zirrgiebel et al., 1995). La microglie exprime également TLR4, un récepteur impliqué dans la réponse immune innée et étant capable de lier le LPS (Okun et al., 2012; Pascual et al., 2012).
I.2. Les astrocytes
Les astrocytes sont caractérisés morphologiquement par leur forme étoilée (astro – étoile et cyte – cellule) et très ramifiée. Ils ont plusieurs fonctions : ils participent au maintien de la barrière hématoencéphalique (BHE), régulent l’approvisionnement en nutriments, l’élimination des déchets, et le métabolisme énergétique du cerveau et participent à la réponse immunitaire. Les astrocytes régulent également la genèse, la stabilisation et l’élimination des
Thèse de Doctorat – Hadi ZARIF 2017 36
neurotransmetteurs, tels que le glutamate, appelés gliotransmetteurs mais également de les capter au niveau synaptique et de réguler ainsi leur recyclage et leur clairance (Buard et al., 2010; Ko & Robitaille, 2015; Ullian et al., 2004). Ils participeraient également à la communication nerveuse par la production de vagues calciques intracellulaires (Scemes & Giaume, 2006).
Les astrocytes régulent finement la plasticité cérébrale en régulant la neurogenèse adulte et la plasticité synaptique, par la sécrétion de facteurs comme le BDNF (Gomez-Casati et al., 2010), l’IL1b, l’IL-6 et le TNFa (Beattie et al., 2002; Lau & Yu, 2001; Van Wagoner & Benveniste, 1999) et par l’expression de protéines membranaires comme la GFAP et la vimentine (Wilhelmsson et al., 2012) (Sims et al., 2015).
I.3. Les oligodendrocytes
Ce sont des cellules de soutien du SNC. Comme les cellules de Schwann au niveau du SNP, la fonction principale de ces cellules est la myélinisation axonale dans le SNC. Les prolongements des oligodendrocytes autour des axones et la fusion des membranes forment la gaine de myéline. Etant un isolant électrique, la gaine de myéline permet une conduction saltatoire (d’un nœud de Ranvier à l’autre) du signal électrique, ce qui engendre une grande rapidité de la propagation de l’influx nerveux (jusqu’à 120 m.s-1). C’est la gaine de myéline qui est reponsable de la couleur blanche de la substance blanche dans le cerveau. Une augmentation de la myélinisation des axones est observée au cours du développement engendrant une accélération de la propagation de l’influx nerveux, alors qu’on observe une démyélinésation dans certaines pathologies (sclérose amyotrophique latérale) et lors du vieillissement.
Thèse de Doctorat – Hadi ZARIF 2017
37
II. Les facteurs moléculaires
La facteurs neurotrophiques comme le BDNF, l’IGF-1 et le NGF sont des molécules qui agissent sur les cellules nerveuses en régulant l’expression de gènes codant pour des protéines anti-apoptotiques, des enzymes antioxydantes et des protéines impliquées dans le métabolisme et l’homéostasie ionique (Lee et al., 2002; Mattson et al., 1995; Mattson et al., 1993). Ainsi, des perturbations génétiques peuvent être à l’origine de troubles de la régulation de la plasticité cérébrale. Des travaux ont par exemple montré qu’une mutation du gène codant pour le BDNF conduit une diminution de la neurogenèse hippocampique (Vithlani et al., 2013).
D’autres molécules comme le glutamate, un neurotransmetteur excitateur, sont capables d’influencer la plasticité cérébrale en régulant la neurogenèse (Suzuki et al., 2006), la synaptogenèse (Mattson et al., 1988) et la survie neuronale (Sendtner et al., 2000). Comme précédemment indiqué, les cytokines pro et anti-inflammatoires sont également impliquées dans la plasticité.
Les hormones peuvent également affecter la plasticité, et en particulier celles de l’axe corticotrope (Yang et al., 2004), l’axe gonadotrope (Galea et al., 2013) et l’axe thyroïdien (Remaud et al., 2014; Vara et al., 2002). Dans le cadre de ma thèse, des résultats (présentés dans l’article 5) m’ont conduit à étudier les effets des hormones thyroïdiennes (HT) sur la plasticité de l’hippocampe. Dans ce contexte, nous nous sommes particulièrement intéressés à l’action d’une hormone hypothalamique, la TRH, dans la plasticité synaptique de l’hippocampe. Le rôle des HT dans les phénomènes de plasticité cérébrale est bien étudié. Cependant, l’essentiel des travaux ont été effectués sur l’action des hormones T3 et T4. Peu d’études se sont intéressées aux effets de la TRH sur la plasticité hippocampique chez l’adulte. Ces études sont souvent contradictoires et concernent exclusivement le GD ou la synapse entre les fibres moussues et CA3 (Gilbert, 2011; Ishihara et al., 1991; Morimoto & Goddard, 1985). Par des approches électrophysiologiques sur des tranches de cerveau, nous avons étudié les
Thèse de Doctorat – Hadi ZARIF 2017 38
effets de la TRH sur les neurones glutamatergiques de la région CA1 de l’hippocampe. Cette étude a été publiée 2016 dans le journal Neuropharmacology (Zarif et al., 2016).
Une brève introduction sur la régulation de l’axe thyroïdien est nécessaire avant de présenter mes travaux personnels.
Les HT font partie des éléments majeurs du neuro-développement et de la neuroplasticité. La biosynthèse des HT par la glande thyroïde est finement régulée par l’axe hypothalamo-hypophysaire. L’hormone thyréotrope TRH (thyrotropin-releasing hormone), un tripéptide de structure Glu-His-Pro produit au niveau de l’hypothalamus, agit sur ses récepteurs au niveau de l’anté-hypophyse provoquant la libération de la thyréostimuline TSH
(thyroid-stimulating hormone) qui va à son tour agir au niveau de la glande thyroïde et stimuler la sécrétion de la triiodothyronine (T3) et la tyroxine (T4) qui vont alors agir sur les cellules cibles. La T3 et la T4 régulent l’axe thyroïdien par rétrocontrôle négatif sur les sécrétions de TRH et de TSH. Le transport de la T4 au niveau central se fait essentiellement par l’intermédiaire de la transthyrétine (TTR) (Eneqvist et al., 2003; Ingbar, 1958), une protéine présente en abondance dans le liquide cérébro-spinal (LCS), synthétisée par les plexus choroïdes (Kabat et al., 1942). La T4 peut être convertie en T3 localement, notamment par les astrocytes (Schroeder & Privalsky, 2014), et la T3 est ensuite captée par les cellules cibles grâce à des transporteurs comme le MCT8, codé par le gène Slc16a2 (Jansen et al., 2005).
Thèse de Doctorat – Hadi ZARIF 2017
Thèse de Doctorat – Hadi ZARIF 2017 40
Article 1 : Effet biphasique de la TRH sur la modulation
des afférences glutamatergiques des neurones CA1 de
l’hippocampe chez la souris
Objectifs et résumé
Dans un premier temps, par des études d’immunofluorescence, nous avons montré que le récepteur de la TRH, la TRHR, est exprimé au niveau des neurones de l’hippocampe.
Des tranches de cerveau de 350 µm d’épaisseur, maintenues en survie, issues de souris femelles C57BL6J âgées de 8 semaines ont été utilisées pour les études électrophysiologiques. Nous avons testé l’effet de la TRH sur :
1) Les potentiels de champ enregistrés au niveau de la région CA1 de l’hippocampe en réponse
à une stimulation des collatérales de Schaffer (fibres CA3-CA1).
2) Les courants post-synaptiques excitateurs (CPSE) enregistrés sur les neurones au niveau de
la région CA1 par la technique de patch-clamp en configuration cellule entière.
3) La plasticité synaptique de la synapse CA3-CA1 par un protocole de PLT.
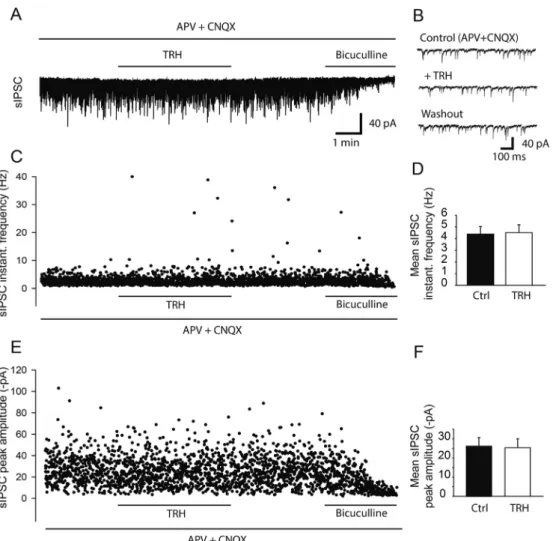
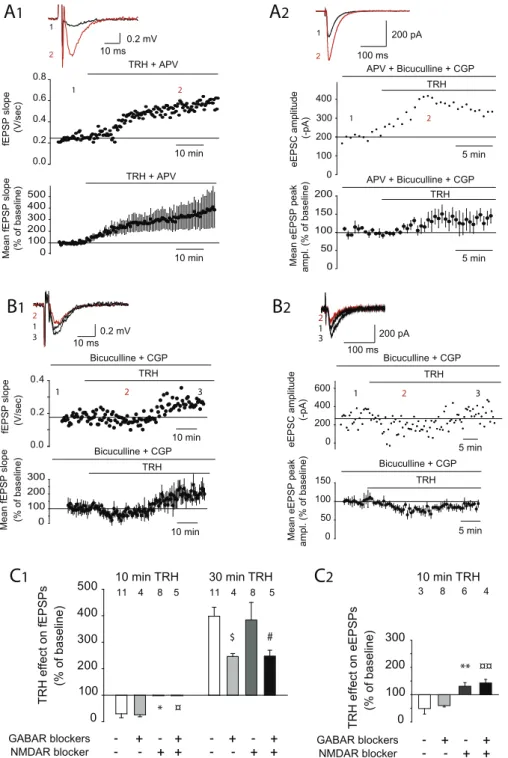
La TRH est capable d’induire un effet biphasique sur la réponse enregistrée par la technique de réponse de champs ou en patch-clamp. Un effet précoce consiste en une diminution de la réponse (» 20 %) 10 min après l’application de la TRH sur les tranches. Un deuxième effet tardif consiste en une grande potentialisation (» 300 %) de la réponse après 30 min d’application.
Thèse de Doctorat – Hadi ZARIF 2017
41
excitateur. Afin d’élucider les mécanismes par lesquels la TRH agit sur la réponse glutamatergique nous avons étudié les rôles putatifs des récepteurs AMPA et NMDA (AMPAR et NMDAR).
De manière surprenante, en isolant la réponse AMPA (blocage des NMDAR en utilisant l’APV, un inhibiteur spécifique) l’effet précoce (la diminution de la réponse) était aboli. La diminution de la réponse engendrée par l’application de la TRH est donc dépendante des NMDAR.
En ce qui concerne l’effet tardif, nous avons émis l’hypothèse que la TRH pourrait induire une dérégulation des réseaux inhibiteurs, d’où l’apparition de cette potentialisation importante. Le GABA étant principal neurotransmetteur inhibiteur, nous avons étudié le rôle des récepteurs GABA sur les effets de la TRH. Les récepteurs (GABAR) GABAA et GABAB ont été bloqués par l’utilisation de bicuculline et de CGP. Dans ces conditions, l’effet précoce de la TRH reste toujours présent tandis que la potentialisation durant l’effet tardif est diminuée de moitié. Ceci démontre que l’effet tardif de la TRH est au moins partiellement dépendant des GABAR.
Les limites techniques du patch clamp en configuration cellule entière ne nous permettant pas d’enregistrer de manière stable la réponse des neurones pendant plus de 30 minutes, nous nous sommes focalisés sur l’effet précoce NMDAR-dépendante de la TRH.
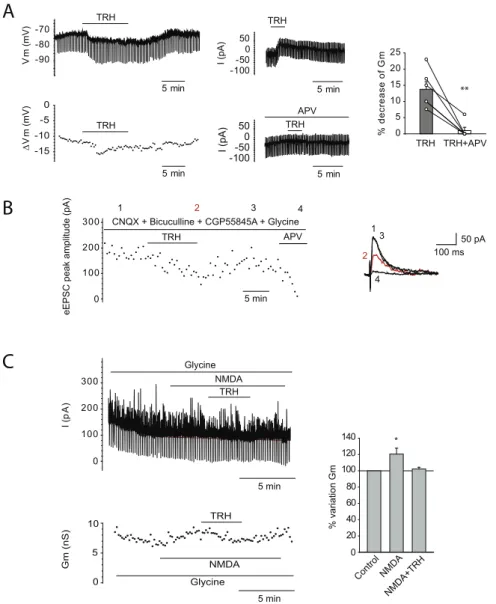
L’application de TRH engendre une hyperpolarisation faible du potentiel membranaire associée à un courant sortant et une diminution de la résistance membranaire sur les neurones pyramidaux de la région CA1 de l’hippocampe. Ceci suggère que la TRH engendre la fermeture de canaux cationiques. Nous avons émis l’hypothèse que ces canaux pourraient correspondre aux NMDAR.
Thèse de Doctorat – Hadi ZARIF 2017 42
réponse NMDA par la technique de patch clamp en configuration cellule entière avec un
potentiel membranaire fixé à +40 mV (afin de lever le bloc de Mg2+ qui bloque les NMDAR).
Nous avons également ajouté de la glycine (un co-activateur des NMDAR) et des bloqueurs des autres récepteurs (AMPAR et GABAR) et nous avons appliqué la TRH. La réponse NMDA ainsi isolée est largement inhibée par l’application de la TRH.
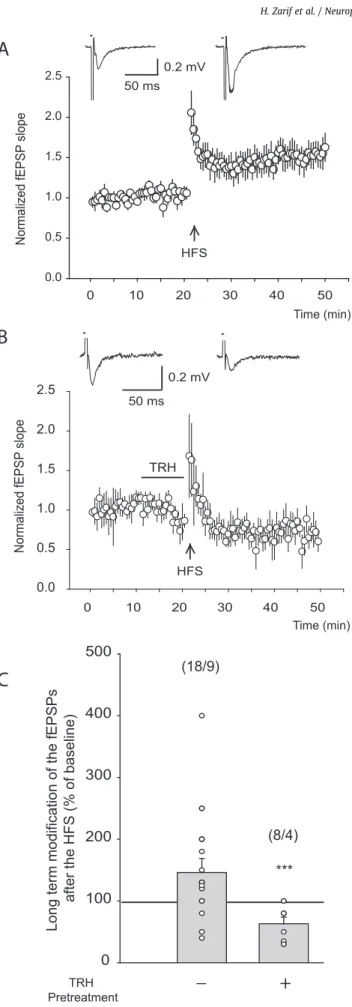
Les NMDAR sont nécessaires à l’établissement de la PLT au niveau de la synapse CA3-CA1. Nous nous sommes donc demandés si le blocage des NMDAR par la TRH était capable d’affecter l’établissement de la PLT. Dans ce but, nous avons appliqué la TRH 10 min avant le protocole de la SHF. Dans ces conditions, le protocole induit une DLT au lieu d’une PLT, indiquant que la TRH est capable, par un blocage des NMDAR, inhiber la PLT.
Conclusion
En conclusion, la TRH est capable d’affecter la plasticité synaptique hippocampique par un blocage des NMDAR et de modifier l’établissement de la PLT.
Cet effet précoce de la TRH sur le blocage des NMDAR peut s’effectuer :
- soit directement : la structure de la TRH (pGlu-His-Pro) et l’acide aminé « Glu »
peuvent suggérer un blocage des NMDAR par un mécanisme compétitif du glutamate en bloquant les sites de fixation.
- soit indirectement : via l’action de son récepteur, TRH-R1, exprimé sur les neurones de l’hippocampe, co-localisé avec la PSD95 au niveau des terminaisons synaptiques.
Thèse de Doctorat – Hadi ZARIF 2017
Thèse de Doctorat – Hadi ZARIF 2017 44
TRH modulates glutamatergic synaptic inputs on CA1 neurons of the
mouse hippocampus in a biphasic manner*
Hadi Zarif, Agn�es Petit-Paitel, Catherine Heurteaux, Jo€elle Chabry, Alice Guyon*
Institut de Pharmacologie Mol�eculaire et Cellulaire, UMR 7275, CNRS, Universit�e de Nice-Sophia Antipolis, F-06560, Valbonne, France
a r t i c l e i n f o Article history:
Received 21 December 2015 Received in revised form 8 March 2016 Accepted 5 April 2016 Available online 7 April 2016
Chemical compounds studied in this article: 2-Amino-5-Phosphonopentanoic acid (DL-APV, PubCem CID 1216)
Bicuculline (PubCem CID 10237) CGP55845A (PubCem CID 123885) 6-Cyano-7-Nitroquinoxaline-2,3-Dione (CNQX, PubCem CID 3721046) Glycine (PubCem CID 750)
N-MethylD-Aspartate (NMDA, PubCem CID
22880) QX314 (PubCem CID 3925) Thyrotropin-releasing hormone (TRH, PubCem CID 32281) Keywords: Thyrotropin-Releasing Hormone (TRH) Hippocampus Mice Glutamatergic transmission NMDA receptors AMPA/kainate receptors Long-Term Potentiation (LTP) a b s t r a c t
Thyrotropin Releasing Hormone (TRH) is a tripeptide that induces the release of Thyroid Stimulating Hormone (TSH) in the blood. Besides its role in the thyroid system, TRH has been shown to regulate several neuronal systems in the brain however its role in hippocampus remains controversial. Using electrophysiological recordings in acute mouse brain slices, we show that TRH depresses glutamate responses at the CA3-CA1 synapse through an action on NMDA receptors, which, as a consequence, decreases the ability of the synapse to establish a long term potentiation (LTP). TRH also induces a late increase in AMPA/kainate responses. Together, these results suggest that TRH plays an important role in the modulation of hippocampal neuronal activities, and they contribute to a better understanding of the mechanisms by which TRH impacts synaptic function underlying emotional states, learning and memory processes.
©2016 Elsevier Ltd. All rights reserved.
1. Introduction
Thyrotropin-Releasing Hormone (TRH) is a tripeptidal hormone (pGlu-His-Pro-NH2) produced by the hypothalamus in medial neurons of the paraventricular nucleus. It is released via the hy-pophyseal portal system, where it stimulates the release of Thyroid-Stimulating Hormone (TSH) in the anterior pituitary (Boler et al., 1969). Besides its role as a release factor controlling thyroid
function, early functional findings suggested a role for TRH in neural modulatory function in different parts of the brain including motor systems (Chizh and Headley, 1994, 1996; Kinoshita et al., 1998; Rekling, 1992), cerebral cortex (Braitman et al., 1980; Kasparov et al., 1994) and several limbic regions (Calza et al., 1992; Hokfelt et al., 1989; Horita, 1998; Sharif and Burt, 1985; Yarbrough, 2003). TRH has been proposed to modulate several neurotransmitter systems for instance GABAergic (Breese et al., 1975; Nemeroff et al., 1975), cholinergic (Braitman et al., 1980), aminergic (Plotnikoff et al., 1975) and glutamatergic systems (Chizh and Headley, 1994, 1996; Kasparov et al., 1994; Kinoshita et al., 1998; Rekling, 1992). Fast excitatory synaptic transmission in the CNS (Central Nervous System) uses glutamate, which acts on
* This paper has been recommended for acceptance by Danny Winder.
*Corresponding author. IPMC, UMR 7275, CNRS, UNSA, 660 Route des Lucioles, F-06560 Valbonne, France.
E-mail address:alice.guyon@ipmc.cnrs.fr(A. Guyon).
Contents lists available atScienceDirect
Neuropharmacology
j o u r n a l h o m e p a g e : w w w . e l s e v i e r . c o m / l o c a t e / n e u r o p h a r m
http://dx.doi.org/10.1016/j.neuropharm.2016.04.004 0028-3908/© 2016 Elsevier Ltd. All rights reserved.