HAL Id: tel-02805974
https://hal.inrae.fr/tel-02805974
Submitted on 6 Jun 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
To cite this version:
Iva Jebava-Ondrackova. Système autolytique de Lactobacillus helveticus. Ingénierie des aliments. Université de Bretagne Occidentale; Ecole Doctorale Vie-Agro-Santé; AGROCAMPUS OUEST, 2011. Français. �tel-02805974�
THESE / AGROCAMPUS OUEST
Sous le sceau de l’Université Européenne de Bretagne pour obtenir le diplôme de :
DOCTEUR DE L'INSTITUT SUPERIEUR DES SCIENCES AGRONOMIQUES, AGRO-ALIMENTAIRES, HORTICOLES ET DU PAYSAGE
Spécialité : « Biochimie – Biologie Moléculaire et Cellulaire»
Ecole Doctorale : VieAgroSanté
présentée par :
Iva JEBAVA née ONDRACKOVA
Système autolytique de Lactobacillus helveticus
Soutenue le 26 mai 2011 devant la commission d’Examen
Composition du jury : Rapporteurs
Monique Zagorec, Directrice de Recherches INRA- FLEC, Jouy
Pascal Le Bourgeois, Directeur de Recherches, Université Paul Sabatier, Toulouse
Peter Šebo, Académie de Science de République Tchèque, Prague Membres
Michel Gautier, Professeur, Agrocampus Ouest, Rennes Kateřina Demnerová, Professeur, ICT Prague
Hana Opatová, Professeur, ICT Prague Jarmila Pazlarová, Professeur, ICT Prague Directeur de thèse
Milada Plocková, Professeur, ICT Prague
Sylvie Lortal, Directrice de Recherches, INRA – STLO, Rennes N° ordre : 2011-11
Résumé
L´autolyse est une dégradation du peptidoglycan par des enzymes bactériennes endogènes appelées hydrolases du peptidoglycane ou autolysines. Grâce à cette réaction enzymatique, la cellule bactérienne peut éclater et le contenu intracellulaire est libéré. Chez Lactobacillus
helveticus, le ferment protéolytique fromager, les enzymes libérées lors de l´autolyse jouent
un rôle important dans le développement de l´arôme et la texture des fromages au cours de l´affinage. Il apparait également que l´autolyse accélère l´affinage de fromage, un processus long et coûteux. Il a été démontré que l´aptitude à l´autolyse était un facteur souche dépendant mais les mécanismes à l´origine de cette diversité n´ont pas été élucidés.
L´objectif de ce travail est de comprendre pourquoi les souches de L. helveticus s´autolysent de manière différente. Nous avons élaboré la stratégie suivante : (i) construire une collection de L. helveticus la plus diverse possible, (ii) rechercher l´origine de la diversité autolytique au niveau génétique et (iii) déterminer le rôle de la paroi cellulaire.
Une collection de référence composée de 26 souches de L. helveticus a été constituée de manière à offrir la plus large diversité possible en termes d´origine géographique et de biotope. La diversité des souches au niveau génomique a été confirmée par électrophorèse en champ pulsé. Le suivi de l´autolyse réalisé en tampon, par microscopie électronique, coloration de Gram et cytometrie en flux, et en milieu fromager modèle ont montré que les souches se distinguent pour leur capacité autolytique. Pour étudier la diversité génétique dans la collection de référence nous nous sommes appuyé sur le génome séquencé de la souche autolytique L. helveticus DPC 4571. Neuf couple d´amorces, correspondant aux neuf gènes codant pour les autolysines prédits chez la souche DPC 4571, ont été dessinés et validés. La présence des gènes codant pour les autolysines ainsi que leur transcription ont été recherchées par PCR et RT-PCR chez les 26 souches. La technique de zymogramme a été utilisée d´une part pour rechercher l´activité enzymatique des autolysines et d´autre part pour rechercher la diversité des parois cellulaires. Puis la diversité des parois cellulaires parmi les souches de L.
helveticus a été étudiée par analyse des acides teïchoiques et par analyse d´activité
enzymatique sur différents substrats par zymogramme.
Les travaux réalisés au cours de cette thèse ont permis de comprendre que la diversité autolytique n´est pas due à une variabilité du nombre des gènes codant pour les autolysines ni à leur transcription. Nous montrons ainsi que ce sont des étapes de traduction et/ou modification post-traductionnelle qui peuvent jouer un rôle dans la diversité autolytique. La composition de l´acide teïchoique varie d´une souche à l´autre ce qui montre une diversité parmi les souches au niveau de la paroi cellulaire. De plus, il a été montré que la cytométrie en flux peut être utilisée pour la mesure de l´autolyse in vitro.
Souhrn
Autolýza je enzymatická reakce spočívající ve štěpení peptidoglykanu pomocí endogenních peptidoglykan-hydrolas neboli autolysinů. Může vést k rozpadu bakteriální buňky a uvolnění intracelulárních enzymů. Uvolněné enzymy, u sýrařské zákysové kultury
L. helveticus, se podílejí na rozvoji senzorických charakteristik sýrů a umožňují urychlit
zdlouhavý a drahý zrací proces. Kmeny L. helveticus se mezi sebou liší autolytickými schopnostmi.
Cílem této práce bylo objasnit příčinu rozdílných autolytických schopností mezi jednotlivými kmeny L. helveticus. Navržená strategie spočívala ve (i) vytvoření co možná nejrozmanitější sbírky L. helveticus, (ii) srovnání distribuce a funkčnosti jejich autolytických genů a (iii) objasnění role buněčné stěny.
Sbírka 26-ti kmenů L. helveticus byla sestavena tak, aby nabídla co nejširší rozdílnost, pokud jde autolytické schopnosti, zeměpisný původ a biotop. U sbírkových kmenů byla prokázána genetická rozmanitost použitím gelové elektroforézy v pulzním poli (PFGE). Dále byl ověřen rozdíl v autolytických aktivitách těchto kmenů pomocí metody měření autolýzy v pufrových systémech, s využitím elektronové mikroskopie, Gramova barvení a půtokové cytometrie, a v modelovém sýrovém médiu. Pro studování genetických odlišností v autolýze mezi sbírkovými kmeny jsme vycházeli z genomu autolytického kmene L. helveticus DPC4571. Na základě analýzy devíti známých genů, kódujících peptidoglykan-hydrolasy pro autolytický kmen L. helveticus DPC 4571, bylo navrženo devět specifických párů primerů. Primery byly uplatněny při detekci a sledování transkripce těchto devíti genů pomocí PCR, resp. RT-PCR u všech sbírkových kmenů. Technika zymogramu byla použita pro detekci aktivity produkovaných autolysinů a pro testování rozdílu ve složení buněčných stěn jednotlivých kmenů. Rozdíl v buněčných stěnách byl dále studován pomocí analýzy složení kyseliny teichoové a pomocí testování různých substrátů technikou zymogramu.
Všech devět genů bylo systematicky přítomných a přepsaných do mRNA u všech studovaných kmenů. Naopak aktivita autolysinů pozorovaná pomocí zymogramu se lišila kmen od kmene a zároveň byla závislá na aplikovaném substrátu. Složení kyseliny teichoové bylo u jednotlivých kmenů odlišné.
Výzkum uskutečněný během této disertační práce prokázal, že rozdíl v autolytických schopnostech mezi kmeny L. helveticus není závislý na počtu přítomných genů ani schopností transkripce v buňkách jednotlivých kmenů. Tyto výsledky naznačují, že rozdíly v autolýze se mohou nacházet na dalších úrovních exprese genů jako je post-translační modifikace nebo regulaci exprese anebo na složení buněčné stěny. Navíc bylo zjištěno, že průtoková cytometrie umožňuje měření autolýzy in vitro.
Summary
Autolysis results from peptidoglycan degradation, by so called endogenous peptidoglycan hydrolases or autolysins, leading to disintegration of the bacterial cell which causes release of intracellular content. In case of L. helveticus, a dairy starter culture uses for cheesemaking, released enzymes by autolysis involves a development of sensory properties and accelerates cheese ripening. It was shown that L. helveticus autolysis is a strain-dependant phenomenon.
The aim of this work was to determine the origin of autolytic diversity of L. helveticus. Following strategy was elaborated: (i) construction of a collection of L. helveticus strains, (ii) evaluation of the diversity for autolytic genes, and (iii) determination of the role of the cell wall.
A collection of 26 stains of L. helveticus, diverse in terms of origin and biotope, was assembled. Pulsed field gel electrophoresis was applied to assess the genomic diversity of all tested strains. Autolytic capacity was evaluated in vitro in buffers (by electron microscopy, Gram staining, flow cytometry) and in situ in a model cheese. Nine genes coding PGHs were previously annotated in the genome of the high autolytic strain L. helveticus DPC 4571. Nine pairs of primers, corresponding to the nine PGHs genes, were designed and validated against DPC 4571. Distribution of the genes coding for autolysins were tested in 26 strains by PCR and RT-PCR. Zymogram was used to detect enzymatic activity of autolysins and also for observed the diversity of the cell wall. The difference in composition of the cell wall among six selected strains was accomplished by analysis of teichoic acid and by zymogram using different substrates.
The nine PGHs genes are ubiquitous and transcribed early during growth. Zymograms were similar in terms of molecular size of the bands, but exhibited strain to strain variations in the number of bands revealing from 2 to 5 lytic bands per strain. Composition of teichoic acid varied from strain to strain.
The work realized during this thesis permitted to understand that the autolytic diversity does not depend on the distribution of autolysins, not even to their transcription. These results indicate that the origin of the autolytic diversity could be due to post-translation steps or to the regulation of peptidoglycan hydrolases, or to the composition of the cell wall. In addition, it was shown that flow cytometry could be used for the determination of autolysis in vitro.
Cette thèse a été préparée à Département de la Technologie du Lait et des Matières Grasses de l´Institut Supérieur de Technologie Chimique de Prague et dans les laboratoires STLO, INRA-UMR 1253 Agrocampus-Ouest de Rennes à partir de septembre 2007 jusqu´à mars 2011.
Je déclare que j´ai rédigé la dissertation individuellement et j´ai dénoté tous les ressources appliquées et paternité commune. Je suis d'accord avec la publication de la thèse de doctorat, conformément à la loi n ° 111/1998 Coll., sur l'enseignement supérieur, tel que modifié. J'étais conscient du fait que mon travail est soumis aux droits et obligations suite de la loi n ° 121/2000 Coll., loi d´auteur, tel que modifié.
Prague le 31 mars 2011
... Signature
Remerciements
Je souhaite tout d´abord exprimer ma reconnaissance aux membres du jury qui m´ont fait l´honneur d’évaluer ce travail de recherche : Monsieur Michel Gautier qui préside ce jury, Madame Monique Zagorec, Monsieur Pascal Le Bourgeois et Monsieur Peter Šebo qui ont accepté d´en être les rapporteurs. Je remercie également à Hana Opatová et Jarmila Pazlarová qui ont assumé la charge d´examinateurs. Plus particulièrement, je remercie mes rapporteurs pour leur lecture attentive du document de thèse malgré un emploi du temps chargé.
Je tiens également à remercier tous les membres du comité de thèse qui ont suivi l´évolution du travail, pour les discussions enrichissants que nous avons menés ainsi que pour les conseils scientifiques qui m´ont accordé.
Mon plus grand MERCI appartient aux trois femmes, Milada Plocková, Sylvie Lortal et Florence Valence-Bertel, qui m´ont influencée et encadrée durant les trois dernières années. Merci pour leurs soutiens et pour toutes ces discussions enrichissantes, tant sur le plan humain que scientifique.
Merci à Milada Plocková, directrice de thèse de coté tchèque, pour ses nombreux conseils et le soutien moral qu’elle m’a apporté durant les moments difficiles de cette thèse.
Merci à Sylvie Lortal, directrice de thèse de coté française, pour m´avoir ouvert les portes de son laboratoire et pour la confiance qu´elle m´a accordée. Ses conseils ont guidé mes réflexions scientifiques tout au long de ces dernières années.
Merci à Florence Valence-Bertel pour sa disponibilité et sa patience pendant l´apprentissage des méthodes utilisées dans ce travail, ainsi que pour la lecture attentive du document de thèse malgré un emploi du temps chargé.
Je tiens à remercier également Hélène Falentin pour son aide précieuse lors de la partie génétique, Valentin Loux pour les nombreux conseils dans le domaine bioinformatique et
Irina Sadovskaya pour le dosage d´acide teïchoiques.
Comme cette thèse a été réalisée en cotutelle entre les laboratoires Science et Technologie de Lait et de L´Œuf, INRA-UMR 1253 Agrocampus-Ouest de Rennes en France et le Département de la Technologie du Lait et des Matières Grasses de l´Institut de Technologie Chimique de Prague en République Tchèque, je voudrais remercier toutes les personnes de ces deux laboratoires pour leur aide et pour les moments agréables que nous avons passés ensemble. Merci à Kateřina Solichová, Céline David, Růžena Bačinová, Victoria Chuat, Sandrine Breton, Marie-Noëlle Madec, Stéphanie-Marie Deutsch, Valérie Gagnaire, Rachel Boutrou, Anne Thiery, Francesco Fancello, Juliana Nobrega, Leila Sadat, Magali Genay et Blandine Hoarau. Ils sont nombreux et cette feuille ne sera jamais assez grande pour les remercier tous.
Enfin je souhaite remercier de tout cœur ma famille : mon mari Martin, mon frère Honza et ma mère, pour leurs soutiens moral au cours de mes longues années d´études.
Ce manuscrit est dédié
à la mémoire
de mon papa
1
TABLES DES MATIERES
Tables des matières ...1
Introduction ...4
Synthèse bibliographique ...5
1 Lactobacillus helveticus ...5
1.1 Le genre Lactobacillus ...5
1.2 L´espèce L.helveticus ...7
1.3 L. helveticus en technologie laitière ...9
1.3.1 Le rôle de L. helveticus en technologie fromagère ... 10
1.3.1.1 L´acidification ... 12
1.3.1.2 La protéolyse ... 12
1.3.1.3 L´autolyse ... 14
2 Le mécanisme d´autolyse ... 19
2.1 Les peptidoglycanes hydrolases (PGHs) ... 19
2.1.1 Classification des PGHs ... 19
2.1.2 Les autolysines ... 20
2.1.2.1 Fonctions des autolysines ... 22
2.1.2.2 Synthèse, sécrétion et régulation des autolysines ... 24
2.1.3 Les endolysines ... 24
2.2 La paroi cellulaire ... 26
2.2.1 Le peptidoglycane... 27
2.2.2 Les autres constituants de la paroi cellulaire ... 29
2.2.2.1 Les polysaccharides anioniques... 29
2.2.2.2 Les polysaccharides neutres ... 30
2.2.3 La couche S ... 30
3 Etat des connaissences sur les PGHs de L. helveticus ... 32
3.1 Les autolysines de L. helveticus ... 32
3.2 Les endolysines de L. helveticus ... 35
3.3 Méthodes de suivi de la lyse et de l´autolyse ... 35
3.3.1 Dosage de la lyse in situ ... 35
3.3.2 Dosage de la lyse in vitro ... 36
Objectif et strategie ... 38
Matériel et méthodes... 40
1 Techniques de microbiologie ... 40
1.1 Les souches utilisées... 40
1.1.1 Constitution de la collection de L. helveticus ... 40
1.1.2 Isolement des souches de L. helveticus ... 40
1.2 Conservation des souches, remise en culture et milieux utilisés ... 43
1.3 Dosage spectrophotométrique ... 43
1.4 Numération bactérienne ... 44
1.5 Mesure du pH ... 44
1.6 Coloration de Gram ... 44
1.7 Dosage des activités enzymatiques ... 45
2
1.9 Observation en microscopie électronique à transmission... 47
1.10 Suivi de la croissance bacterienne dans le milieu MRS ... 47
1.11 Suivi de la lyse induite par le transfert des cellules en solution tamponnée... 47
1.11.1 Transfert des cellules en solution tamponnée ... 47
1.11.2 Détermination de la capacité autolytique en solution tamponnée ... 48
1.11.3 Analyse statistique ... 49
1.12 Suivi de la lyse dans du retantat ultrafiltré ... 49
1.12.1 Préparation du lait ultrafiltré ... 49
1.12.2 Détermination de la capacité autolytique dans du lait ultrafiltré... 50
2 Techniques de biologie moléculaire ... 50
2.1 Isolement des acides nucléiques... 50
2.1.1 Extraction de l´ADN ... 50
2.1.2 Extraction de l´ARN ... 51
2.2 Transcription inverse ... 51
2.3 Réaction en Chaîne par Polymérase (PCR) ... 52
2.4 Purification des fragments de PCR ... 55
2.5 Séquençage ... 55
3 Techniques électrophorétiques ... 56
3.1 Électrophorèse d´ADN sur gel d´agarose ... 56
3.2 Électrophorèse en champs pulsés (PFGE) ... 56
3.2.1 Préparation des blocs d´agarose ... 56
3.2.2 Digestion de l´ADN par les endonucleases ... 57
3.2.3 Préparation des gels et migration électrophorétique ... 58
3.2.4 Analyse des gels ... 58
3.3 Électrophorèse SDS-PAGE ... 59
3.3.1 Préparation des échantillons ... 59
3.3.2 Formation des gels d´acrylamide et conditions de migration ... 59
3.4 Zymogramme ... 60
3.4.1 Préparation des substrats pour le zymogramme ... 60
3.4.1.1 Cellules de M. lysodeikticus ... 60
3.4.1.2 Cellules de L. helveticus non-autoclavées ... 60
3.4.1.3 Cellules de L. helveticus autoclavées ... 61
3.4.1.4 Parois cellulaire de L. helveticus ... 61
3.4.2 Réalisation des zymogrammes et renaturation ... 61
4 Techniques biochimiques ... 62
4.1 Analyse des acides teïchoiques ... 62
4.1.1 Préparation des parois cellulaires ... 62
4.1.2 Extraction des ATs ... 63
4.1.3 Analyse de la composition des ATs... 63
5 Techniques bio-informatiques ... 63
5.1 Recherche des gènes codant pour les PGHs ... 64
5.2 Construction des amorces ... 64
5.3 Analyses des séquences d´ARNr 16S... 64
5.4 Analyses des séquences de PHGs ... 65
Résultats et discussion ... 67
1 Mode de présentation des résultats ... 67
2 Constitution et caractérisation de la collection bactérienne ... 68
2.1 Isolement et identification des souches ... 68
3
2.2.1 Caractérisation de la diversité génomique ... 69
2.2.2 Etude de la variabilité des profils protéiques ... 69
2.2.3 Autolyse « in vitro » en milieu tamponné ... 70
2.2.4 Autolyse en matrice laitière ... 71
3 Neuf gènes de peptidoglycane hydrolases sont ubiquitaires et transcrits de manière précoce chez Lactobacillus helevticus (Article 1) ... 76
4 Analyse des génomes de L. helveticus ... 84
4.1 Vérification des relations d´orthologies ... 84
4.2 Vérification de la taille des gènes ... 86
4.3 Analyse des séquences des PGHs ... 88
5 Proposition de PCR espèce-spécifique pour l´identification de L. helveticus (Article 2) ... 91
6 Recherche des différences entre les parois cellulaires de L. helveticus ... 102
6.1 Les différences de la paroi cellulaire mise en evidence par les zymogramme……….102
6.2 Les différences de la paroi cellulaire mise en place par l´analyse d´acides teïchoiques ... 104
Conclusions, Synthèse et Perspectives ... 106
Références bibliographiques ... 114
Liste des abréviations utilisées ... 125
Annexes ... 127
Communications et matériels scientifiques ... 143
1 Participation à des conférences ... 143
1.1 Présentation ... 143
1.2 Posters ... 143
2 Articles ... 144
2.1 Résumés parus dans les recueils de conférences... 144
2.2 Articles parus dans les journaux internationaux sélectionné par le comité de lecture……….………..145
4
INTRODUCTION
Le fromage est un aliment hautement nutritif de notre régime alimentaire. C´est une source majeure de protéines et de calcium pour de nombreux Européens. Un morceau de fromage à pâte dure de la taille d´une petite boîte d´allumettes (40 à 50 g) satisfait environ un cinquième des besoins en protéines d´un adulte et fournit à peu près un tiers du calcium dont les adolescents ont besoin chaque jour. Le fromage contient également des vitamines A, B2,
B3, B12 et D, des minéraux comme le zinc et le phosphore (Walther et al., 2008).
La fabrication des fromages à pâte dure et extra dure, dont Lactobacillus helveticus est un ferment essentiel, est un processus long et coûteux. L´étape d´affinage, qui est une période très importante pour le développement de la texture et de l´arôme, consomme le plus de temps (en fonction du fromage de 6 à 24 mois) et génère des coûts de stockage importants (Fox et al., 2004). Beaucoup de méthodes visant à raccourcir le temps d´affinage et à améliorer les caractéristiques sensorielles en même temps ont été essayées comme par exemple : l´application de souches lysogènes (Deutsch et al., 2002a), l´application de souches produisant des bactériocines (Garde et al., 2007), l´application de levains atténuées (Koch et al., 2008) ou la production d´OGM (Pillidge et al., 2002). Il semble qu´une autre voie possible pour accélérer l´affinage du fromage soit l´utilisation de souches autolytiques (Lortal et al., 2005).
L’autolyse est un phénomène d’ouverture de la cellule bactérienne. Elle résulte de la dégradation du peptidoglycane, constituant de la paroi, par des hydrolases de peptidoglycane endogènes appelées autolysines. Ces enzymes assurent des fonctions biologiques essentielles au sein de la cellule bactérienne comme la croissance bactérienne et la séparation des cellules filles (Chapot-Chartier, 1996). L. helveticus est considéré comme un des ferments les plus protéolytiques (Sadat-Mekmane et al., 2011). Suite à l’autolyse, les enzymes bactériennes intracellulaires, notamment des peptidases, sont libérées au sein de la matrice fromagère où elles peuvent alors participer à l’élaboration de la texture et de la flaveur du fromage (Hannon
et al., 2007, Hickey et al., 2007). L´autolyse participe donc de manière indirecte à la
protéolyse au cours de l´affinage d´ou l´importance de ce phénomène en technologie fromagère et la nécessité de prendre le critère d´aptitude à la lyse en compte pour la sélection des ferments.
L’aptitude à l’autolyse est un caractère souche dépendant mais, jusqu’à ce jour les mécanismes à l’origine de cette diversité ne sont pas vraiment élucidés.
5
SYNTHESE BIBLIOGRAPHIQUE
1
LACTOBACILLUS HELVETICUS
1.1 LE GENRE LACTOBACILLUS
Le genre Lactobacillus, appartient au groupe des bactéries lactiques, son métabolite principal est l´acide lactique. Celui-ci peut inhiber la croissance de certaines flores indésirables et est également un agent technologique participant à la texture et à la flaveur typique de certains produits laitiers (Crow et Curry, 2003).
Les bactéries appartenant au groupe de Lactobacillus sp. sont des bactéries à Gram positif, asporulées, catalase négatives formant des bâtonnets réguliers immobiles de taille variable d´une longueur de 1 à 10 µm et d´une épaisseur de 0,2 à 1,2 µm en fonction de l´espèce étudiée. Les lactobacilles sont anaérobies obligatoires ou aérotolerants, leur métabolisme est strictement fermentaire. Ils exigent pour une croissance adéquate, la présence de nombreux nutriments dans leur milieu de culture comme par exemple des acides aminés, des peptides, des saccharides, des acides gras et des minéraux. Une partie de leur demande nutritionnelle complexe est garantie par un grand assortiment d´enzymes protéolytiques (Torriani et al., 1994, Hammes et Hertel, 2006).
La première classification des lactobacilles a été réalisée par Orla-Jensen en 1919. Elle est constituée de trois groupes en fonction de leur caractère homo/hétérofermentatif et de leur température optimale de croissance : Streptobacterium, Thermobacterium et Betabacterium. Dans l´ouvrage « Bergey´s Manual of Systematic Bacteriology » (Kandler et Weiss, 1986), les lactobacilles sont classés dans la section 14 « Bâtonnets réguliers Gram-positifs asporulés » et sont répartis en trois groupes similaire aux groupes d´Orla-Jensen : A - homofermentaires obligatoires, B - hétérofermentaires facultatifs et C - hétérofermentaires.
Des études phylogénétiques basées sur l´analyse des séquences de l´ADN codant pour l´ARNr 16S ont permis de réévaluer les espèces (Stackebrandt et al., 2002). Elles montrent que les lactobacilles peuvent être divisés en trois groupes principaux: a L. delbrueckii, b
6 Prenant en compte les classifications phénotypiques et phylogénétiques, les lactobacilles peuvent être divisés (Annexe 3) en six groupes Aa, Ab, Ba, Bb, Cb et Cc. Les capitales A, B et C font référence au type fermentaire et les minuscules a, b et c font référence aux divisions phylogénétiques en prenant en considération le type de peptidoglycane (Holzapfel et Wood, 1995, Hammes et Vogel, 1995, Stiles et Holzapfel, 1997).
Dans le groupe A on trouve des lactobacilles à métabolisme homofermentatif obligatoire. Par la voie métabolique d´Embden-Meyerhof-Parnas (EMP), ils transforment des hexoses en acide lactique qui est le produit unique (à plus de 85 %) de leur métabolisme saccharidique. Bien que possédant l´enzyme fructose-1,6-biphosphate aldolase, ils ne sont pas capables d´utiliser les pentoses et le gluconate car ils ne possèdent pas de phosphocétolase. Deux sous-groupes ont été établis : le sous-groupe Aa caractérisé par le type de peptidoglycane « Lys-D-Asp » et aussi appelé le groupe de L. delbrueckii, et le sous-groupe Ab caractérisé par un autre type de peptidoglycane contenant souvent l´acide diaminopimelique (DAP).
Le groupe B réunit les lactobacilles à un métabolisme hétérofermentatif facultatif qui transforment des pentoses et des hexoses, selon les conditions environnementales en acide lactique, éthanol ou acide acétique et CO2. En présence de glucose, la fermentation des
pentoses est réprimée. Deux sous-groupes ont été établis : le sous-groupe Ba caractérisé par le type de peptidoglycane « Lys-D-Asp », et le sous-groupe Bb caractérisé par le type de peptidoglycane « DAP ».
Le dernier groupe, Groupe C, est constitué des lactobacilles à métabolisme hétérofermentatif obligatoire. Grâce à la chaîne phosphoglucuronate, ils transforment des pentoses et des hexoses majoritairement en éthanol ou acide acétique et CO2. La production
de gaz est significative. Deux sous-groupes ont été établis Cb et Cc. Le sous-groupe Cb est caractérisé par le type de peptidoglycane « DAP », similaire au groupe Bb mais n´ayant pas de voie EMP. Le sous-groupe Cc est caractérisé par d´autres types de peptidoglycane.
Les lactobacilles sont très répandus parmi les végétaux et les animaux ainsi que chez l´homme où on les trouve notamment au niveau du tractus vaginal et gastro-intestinal. Ce sont des bactéries d´intérêt alimentaire notoire utilisées entre autre pour la production des aliments fermentés comme les produits laitiers, le pain, le saucisson, la choucroute, les pickles ou encore pour l’ensilage (Crow et Curry, 2003).
Actuellement on connait 192 espèces et sous-espèces dans ce genre (DSMZ, 2010) dont 15 espèces ont été complètement séquencées et annotées (Tab. 1) et 13 espèces sont au cours de séquençage ou sous forme de draft « draft assembly » (NCBI, 2010).
7
Tab. 1 L´espèces du genre Lactobacillus complètement séquencés jusqu’ à 2010 (NCBI, 2010).
Génomes disponibles Groupe
L .acidophilus Aa L. amylovarus Aa L. brevis Cb L. casei Bb L. crispatus Aa L. delbrueckii Aa L. fermentum Cb L. gasseri Aa L. helveticus Aa L. johnsonii Aa L. plantarum Bb L. reuteri Cb L. rhamnosus Bb L. sake Bb L. salivarius Ab 1.2 L´ESPECE HELVETICUS
Lactobacillus helveticus (Tab. 2) est un lactobacille thermophile à métabolisme
homofermentatif obligatoire du groupe Aa - L. delbrueckii. Ce groupe se subdivise en deux sous-groupes génotypiques : le sous-groupe L. delbrueckii - comprenant L. delbrueckii et ses sous-espèces (bulgaricus, delbrueckii, lactis), et le sous-groupe L. acidophilus - regroupant les espèces restant y compris L. helveticus. Certaines souches de L. helveticus ont la capacité de se développer à 50 - 52 °C, ce qui explique son ancien nom de Thermobacterium
helveticum.
Tab. 2 Taxonomie de Lactobacillus helveticus.
Empire Procaryotes Règne Bactérie Phylum Firmicutes Classe Bacilli Ordre Lactobacillales Famille Lactobacillaceae Genre Lactobacillus
Espèce Lactobacillus helveticus
Il est difficile de distinguer les membres du groupe Aa L. delbrueckii. C´est pourquoi leur classification taxonomique nécessite à la fois des critères phénotypiques et génotypiques. Parmi les lactobacilles, L. helveticus se caractérise phénotypiquement par la production des isomères optique D(-) et L(+) du lactate, par sa capacité à se développer à 45 °C et par l´utilisation du lactose, du glucose et du galactose. L´identification de L. helveticus au niveau
8 génomique peut être réalisée par PCR espèce-spécifique en détectant des gènes codant pou les macromolécules ubiquitaires telles que les protéines de la couche S (Ventura, 2000) ou les peptidases (Fortina et al., 2001). L´identification basé sur la séquence nucléotidique d´ARNr 16S donne souvent des résultats mitigés. Des méthodes basées sur les techniques moléculaires tels que repPCR, RAPD et PFGE ainsi que la méthode de SDS-PAGE sont utilisées pour la caractérisation (Drake et al., 1996b, Lortal et al., 1997c, Lombardi et al., 2002).
Le premier génome de L. helveticus qui a été séquencé est celui de la souche DPC 4571, connue comme étant fortement autolytique. Cette souche a été séquencée par Callanan et publiée dans les bases de données génomiques (NCBI, EMBL-EBI, GenomeNet) en janvier 2008. Elle possède un chromosome circulaire dont la taille est de 2,1 Mbp et 1610 protéines ont été identifiées et annotées à partir de sa séquence (RefSeq : NC_010080). Aucun prophage n’a été prédit (Callanan et al., 2008).
En 2010, un deuxième génome de L. helveticus appartenant à la souche type DSM 20075, a été séquencé et publié sous forme de draft « draft assembly ». La taille de ce génome est de 1,8 Mbp et il coderait pour 2078 protéines (RefSeq : NZ_ACLM00000000). Trois autres souches sont aujourd´hui en cours de séquençage (NCBI, 2010): CNRZ 32 (Utah State University and University of Wisconsin-Madison, USA), H9 et H10 (Inner Mongolia Agricultural University, Chine).
Deux plasmides de L. helveticus, pLH1 (NCBI, RefSeq : NC_002102) et pLJ1 (NCBI, RefSeq : NC_001379) ont été séquencés et codent pour des protéines de fonction inconnue.
L. helveticus est phylogénétiquement très proche de l´espèce L. acidophilus (Fig. 1).
L´homologie de séquence de l´ARNr 16S entre L. helveticus DPC 4571 et L. acidophilus NCFM est de de 98,4 %. La proximité de ces espèces est confirmée également par le fait que 75 % d´ORFs de L. helveticus DPC 4571 possèdent des gènes orthologues chez L. acidophilus NCFM (Callanan et al., 2008). Une autre caractéristique du génome de L. helveticus DPC 4571 réside dans la similarité génétique avec des lactobacilles provenant du tractus gastro-intestinal (Slattery et al., 2010). Callanan et al. (2007) et Kaleta et al. (2009) ont émit l´hypothèse que la diversité entre les souches de L. helveticus était liée aux séquences d´insertion « Insertion sequence elements ». En effet, 213 séquences d´insertion ont été distinguées chez DPC4571, ce qui est dix fois plus que chez les autres espèces.
9
Fig. 1 Arbre phylogénétique des lactobacilles et des streptocoques séquencés. Les membres du groupe
L. acidophilus - L. delbrueckii sont entourés par un ovale et les ferments laitiers sont sur fond gris (Callanan et al., 2008).
1.3 L. HELVETICUS EN TECHNOLOGIE LAITIERE
Lactobacillus helveticus est un ferment thermophile principalement utilisé dans la
fabrication des fromages mais aussi dans la fabrication des boissons fermentées et des yaourts.
Dans les produits fermentés liquides, comme Evolus (Valio, Finlande) et Calpis (Calpis, Japon), il est utilisé pour la production de peptides bioactifs et pour son caractère probiotique (Vinderola et al., 2007, Jauhiainen et al., 2007, Slattery et al., 2010). Dans les fromages,
L. helveticus est utilisé principalement en raison de son caractère acidifiant et surtout pour son
aptitude protéolytique.
L. helveticus, seul ou en combinaison avec d´autres microorganismes, est utilisé comme
ferment dans la fabrication de plusieurs types de fromages (Fig. 2). Ce sont notamment les fromages à pâte pressée cuite extra dure de type Grana Padano, Parmigiano Reggiano et Provolone (type italien), les fromages à pâte pressée cuite dure de type Emmental et Gruyère (type suisse) ainsi que les fromages semi-durs comme Saint-Paulin, Edam, Gouda et Cheddar (Fox et al., 2004). L. helveticus sert de ferment principal pour les fromages de type italiens et suisses et de ferment secondaire pour les fromages semi-durs.
10
1.3.1 Le rôle de L. helveticus en technologie fromagère
Le fromage est une méthode millénaire de conservation du lait qui permet, en outre, de concentrer et de conserver les protéines et la matière grasse du lait. En transformant le lait de l´état liquide à l´état solide, cet aliment hautement nutritif devient facilement transportable et stockable (Eck et al., 1987).
Les fromages sont obtenus par transformation du lait suivant cinq étapes clés : l´ensemencement d´un ferment, la coagulation, l´égouttage, le salage et l´affinage. Il existe une très grande diversité des types de fromages mais les étapes de fabrication sont globalement les mêmes quelque soit le fromage considéré.
La coagulation résulte de la production d´acide lactique par les ferments conjuguée à l´action de la présure qui entraine la précipitation des micelles de caséines et donc à la formation du caillé. Cette étape est suivie de l´égouttage qui inclue les opérations de tranchage du caillé, de brassage et finalement de pressage. Le salage, effectué en saumure ou en surface, participe au développement de la flaveur et limite la croissance des microorganismes indésirables (Eck et al., 1987).
Dans le cas des fromages à pâte pressée cuite, le brassage est suivi d´une cuisson du mélange caillé-sérum. Cette cuisson ainsi que l´égouttage permettent d´optimiser la teneur en eau du fromage (Eck et al., 1987).
L´affinage, étape complexe et clé pour la fabrication des fromages à pâte pressée cuite, dure de plusieurs semaines à plusieurs mois. Pendant cette étape se déroulent plusieurs types de réactions biochimiques dont les plus importantes sont : la fermentation des sucres, l’hydrolyse des protéines (protéolyse) et l´hydrolyse de la matière grasse (lipolyse). Au cours de l´étape d´affinage, le fromage acquiert sa typicité : aspect, consistance, saveur, odeur et texture (Fox et al., 2004).
Ce sont les paramètres technologiques (composition du ferment, type de présure utilisé, durée de coagulation, taille des grains de caillé, principe de salage, température et durée d´affinage) qui différent d´un fromage à l´autre (Fig. 2). Selon les paramètres appliqués, on obtient des fromages avec des caractéristiques complètement différentes (Fox et al., 2004).
L. helveticus participe au développement de la flaveur au cours de l´affinage grâce à
l´action de ses enzymes protéolytiques. Comme, à l´exception de la protéase pariétal, la majorité des enzymes protéolytiques se trouvent dans le cytoplasme, la lyse des cellules du ferment est indispensable pour leur libération dans la matrice fromagère.
11
12 La lyse cellulaire peut être causée par l´action des autolysines, on parle alors d´autolyse. Un potentiel autolytique élevé est de ce fait un des paramètres technologiques recherchés dans les ferments fromagers.
D´autre part, L. helveticus participe également à la sécurité alimentaire en produisant des bactériocines, comme par exemple l´Helveticin J (Joerger et Klaenhammer, 1986) ou l´Helveticin V-1829 (Vaughan et al., 1992), qui inhibent la croissance des autres lactobacilles et des flores indésirables.
Le rôle acidifiant ainsi que l´impact de l´activité autolytique et protéolytique de
L. helveticus sont développés dans les paragraphes suivants.
1.3.1.1 L´acidification
La voie métabolique principale produisant de l´énergie sous forme d´ATP chez
L. helveticus génère de l´acide lactique. L. helveticus a une forte capacité acidifiante, ce qui
favorise la coagulation et présente une avantage en termes de sécurité sanitaire (Neviani et al., 1995).
L´acidification du milieu ainsi générée est un facteur fondamental pour la transformation du lait en fromage. Elle provoque sa coagulation responsable de la concentration en matière sèche. La production d´acide limite également la croissance et la survie de microorganismes indésirables susceptibles d´altérer la qualité des fromages ou de provoquer des intoxications alimentaires.
1.3.1.2 La protéolyse
La protéolyse est très importante pour l´affinage du fromage. Elle fait partie du catabolisme protéique. C´est une hydrolyse enzymatique en cascade des protéines du lait en peptides, puis en oligopeptides et finalement en acides aminés libres qui sont des précurseurs d´arômes (Fig. 3). En plus de la chymosine, présente dans la présure, et de la plasmine, présente ordinairement dans le lait, les enzymes protéolytiques des bactéries lactiques participent aussi à l´affinage du fromage et sont responsables du développement de la texture et de la flaveur des fromages (Fox et al., 2004, Shin et al., 2004). L´activité protéolytique de
L. helveticus est un facteur souche dépendant (Jensen et al., 2009, Sadat-Mekmene, 2011).
La Chymosine (EC.3.4.23.4), de la présure, est une protéase qui hydrolyse spécifiquement la liaison Phe (105) - Met (106) des κ-caséines. Elle génère ainsi deux peptides : la para–κ-caséine (1-105) et le caséinomacropeptide (106-169). Cela provoque la déstabilisation des micelles de caséines qui, en présence d´ions calcium, forment le caillé.
13
Fig. 3 Représentation schématique du système protéolytique de L. helveticus (Sadat-Mekmene, 2011).
De plus, pendant l´affinage du fromage, elle participe à l´hydrolyse des autres caséines (αs1-,
αs2- et para-κ-caséine). La Plasmine (EC.3.4.21.7.), naturellement présente dans le lait et très
thermostable, est une protéase qui joue un rôle important dans la protéolyse des αs2- et β –
caséines pendant l’affinage (Fox et al., 2004).
Les protéinases de paroi (CEPs – cell envelope proteinases) sont localisées dans l´enveloppe cellulaire (Fig. 3). Elles initient la dégradation des caséines et libèrent des oligopeptides. Chez L. helveticus, 4 CEPs, nommées PrtH, PrtH2, PrtH3 et PrtH4, ont été citées mais ont fait l´objet d´un nombre inégal de travaux : deux seulement sont caractérisées de manière approfondie. La protéinase PrtH a été caractérisée chez L. helveticus CNRZ 32 (Pederson et al., 1999). Sa masse moléculaire est de 204 kDa et sa séquence protéique est identique à 45 % avec celle de PrtP, une protéinase de lactocoque. Des expériences de délétion du gène prtH ont montré qu’il existe au moins deux CEPs diffèrent chez L. helveticus (Pederson et al., 1999). Le gène prtH2, quand a lui est ubiquitaire chez L. helveticus (Genay
et al., 2009). Il a été démontré que les souches possédant simultanément les deux protéinases
ont un plus fort pouvoir hydrolytique des αS1-caséines que les souches ne possédant que PrtH2
(Sadat et al., 2011). La PrtH3 et la PrtH4 ont été notées seulement chez la souche CNRZ 32 (Broadbent et al., 2008, Jensen et al., 2009).
Les oligopeptides générés sous l´action des CEPs sont transportés dans le cytoplasme où ils sont dégradés en acides aminés par un ensemble de peptidases (Fig. 3). Les peptidases
14 peuvent être divisées, selon leur site de coupure de la séquence protéique, en endopeptidases, exopeptidases (carboxypeptidases et aminopeptidases) et enfin en di- et tri- peptidases. Chez
L. helveticus plusieurs endopeptidases, di- et tri- peptidases ainsi que des aminopeptidases ont
été isolées et caractérisées (Fox et al., 2004). Les endopeptidases PepE, PepO, PepO2 et PepO3 et l´aminopeptidase PepN contribuent à la réduction des peptides amers (Soeryapranata et al., 2007). Chez les bactéries lactiques, toutes les peptidases sont intracellulaires (Monnet et al., 1995).
Les acides aminés résultant de l´hydrolyse des caséines contribuent à la saveur du fromage soit par leurs propriétés organoleptiques intrinsèques, soit en tant que précurseurs d´arôme. Le catabolisme des acides aminés se produit selon deux voies qui conduisent à la formation des composés aromatiques.
La première voie est initiée par une réaction de transamination : c´est la principale voie chez les bactéries lactiques. Les aminotransférases coupent le groupe amine d´un acide aminé et créent un α-cétoacide qui est ensuite dégradé en différents composés aromatiques tels que : aldéhydes, alcools, acides carboxyliques et esters. La seconde voie est initiée par des réactions d´élimination, catalysées par des lyases. Cette voie concerne la méthionine et les acides aminés aromatiques. Elle conduit à la formation d´arômes de type methanthiol.
L. helveticus possède un système protéolytique efficace. L´ensemble de ses protéinases et
peptidases lui permet d´hydrolyser les β- et αs1-caséines pour satisfaire ses demandes
nutritionnelles, ce qui en même temps influe positivement sur l´affinage du fromage (Bockelmann et al., 1995). L. helveticus possède une plus grande activité aminopeptidasique et dipeptidasique que les autres bactéries lactiques (Hickey et al., 1983, Sasaki et al., 1995). C´est pourquoi l´utilisation de L. helveticus durant la fabrication du fromage favorise l’augmentation de ses qualités sensorielles (Drake et al., 1997) et aide à accélérer l´affinage du fromage (Bartels et al., 1987). De plus, L. helveticus est capable de dégrader les peptides hydrophobes générés par la présure grâce à ses peptidases, ce qui permet de réduire le niveau d´amertume dans le fromage (El Abboudi et al., 1991, Fajarrini et al., 1999).
1.3.1.3 L´autolyse
L´autolyse, lyse cellulaire causée par des autolysines, provoque la libération des enzymes intracellulaires dans le fromage qui accèdent ainsi à leur substrat et peuvent catalyser les réactions protéolytiques (Dako et al., 1995).
15 La capacité autolytique est de ce fait un des paramètres technologiques clé pour le choix des ferments utilisés en technologie fromagère. Dans la production de yaourts et de boissons fermentées probiotiques l´autolyse des ferments est en revanche considérée comme un facteur négatif (Yokoi et al., 2004).
Du point de vue économique, l´affinage est un processus lent qui implique des coûts de stockage importants. L´affinage des fromages, dans lesquels L. helveticus est utilisé comme ferment, peut durer de 6 à 24 mois (Fox et al., 2004). De nombreuses études ont été réalisées pour essayer d´accélérer cette étape, tout en préservant la qualité des fromages.
Différentes méthodes pour accélérer affinage en favorisant la lyse du ferment cours de l´affinage du fromage ont été développées :
1) l´addition de sel. Le stress osmotique initié par l´addition de sel au milieu nutritif provoque la lyse du ferment (Piuri et al., 2005) et augmente le taux d´autolyse (Palomino et al., 2009).
2) l´utilisation de souches lysogènes. Steen et al. (2007) ont montré que l´utilisation de
Lc. lactis lysogène pouvait accélérer et augmenter le niveau la lyse et ainsi améliorer
la qualité sensorielle du fromage. Les souches lysogènes sont considérées comme étant autolytiques. Chez les bactéries lactiques, l´autolyse a été décrite le plus en détail pour Lcactococcus lactis. Cette espèce est caractérisée par la présence de plusieurs autolysines (voir 2.2.5) ainsi que par la présence de phages. Il a été montré qu´une activité autolytique élevée de la souche Lc. lactis subsp. cremoris AM2 était corrélée à l´induction d´un prophage (LePeuple et al., 1998a, LePeuple et al., 1998b). En revanche chez L. helveticus l´induction d´un prophage n´était pas à l´origine de la lyse de L. helveticus CNRZ 303 dans le fromage (Deutsch et al., 2002b).
3) l´addition d´enzymes lytiques. Le ferment lactique peut être lysé également par les enzymes lytiques d´autres microorganismes comme par exemple Streptomyces
fulvissimus ajoutés artificiellement dans le fromage (Ohbuchi et al., 2001).
4) l´utilisation de souches produisant des bactériocines. Les souches produisant des bactériocines sont utilisées pour augmenter la lyse du ferment. La capacité des souches à se lyser sous l´action d´une bactériocine est un phénomène souche-dépendante (Martinez-Cuesta et al., 1997). Sous l´influence de la lacticin 481, bactériocine produite par Lc. lactis subsp. lactis INIA 639, la souche commerciale hautement protéolytique L. helveticus LH 92 se lyse plus facilement (Garde et al., 2007). Utilisation de Lc. lactis ssp. lactis biovar. diacetylactis UL719, producteur de nisine,
16 associée à la souche protéolytique L. casei L2A, augmente la protéolyse et améliore la texture du Cheddar (Sallami et al., 2004).
5) l´utilisation des ferments atténués. Les ferments atténués par choc thermique (chauffage, congélation), par choc osmotique (lyophilisation, séchage) ou par traitement au lysozyme, sont prédisposés à la lyse. Une augmentation de la protéolyse responsable d´une accélération de l´affinage a été observées pour des fabrications de fromages employant ces méthodes d´atténuation (Bartels et al., 1987, Klein et Lortal, 1999, Madkor et al., 1999, Koch et al., 2008).
6) l´application de souches génétiquement modifié (OGM). Des OGMs de Lc. lactis, ayant une activité autolytique élevée, ont été construits (Pillidge et al., 2002) par insertion du gène de l´autolysine majeure de Lc. lactis, AcmA, et d´un promoteur effectif. Les promoteurs activés par du sel (NaCl – choc osmotique), par le choc thermique, par le pH et par la nisine ont été testés. Il a ainsi été montré que les OGMs avaient une activité autolytique cinq fois supérieure. Cependant, l´utilisation des OGMs est limitée par les directives de l´Union Européenne (1829/2003) ainsi que par le manque d´intérêt des acheteurs (Lortal et al., 2005).
Une voie effective pour augmenter la qualité organoleptique du fromage est la sélection de souches autolytiques de L. helveticus (Courtin et al., 2002, Hannon et al, 2005, Hannon et al., 2006, Hannon et al., 2007, Piraino et al., 2008). L´autolyse est un moyen d´accélérer l´affinage du fromage, d´obtenir un développement plus rapide de ses qualités organoleptiques, d´intensifier certains arômes typiques ainsi que diminuer de l´amertume dans le fromage (Wilkinson et al., 1994, Drake et al., 1996a, Courtin et al., 2002, Hannon et al., 2003, Hannon et al., 2006, Hannon et al., 2007, Hickey et al., 2007). L. helveticus est utilisé avec succès dans la production de fromages ayant une teneur base en matières grasses pour améliorer leur arôme et leur texture (Hannon et al., 2006).
La connaissance et la maîtrise de l´autolyse chez L. helveticus est donc un facteur économiquement important. La sélection de souches autolytiques offre une possibilité de contrôler l´autolyse dans le fromage. De nombreuses études ont été menées pour évaluer le potentiel autolytique de L. helveticus.
L´influence de la température, du pH, de la concentration en magnésium, et du stade de croissance a été étudiée sur l´autolyse induite par un choc osmotique chez L. helveticus CNRZ 414 in vitro par Lortal et al. (1989). Il a été ainsi montré que la température optimale
17 pour l´autolyse de la souche CNRZ 414 était située entre 40 et 45 °C. Au-delà de 45 °C, le système est inhibé. Les cellules récoltées en début de phase exponentielle de croissance s´autolysent de manière plus marquée que les cellules récoltées à d´autres stades de croissance. L´effet du magnésium sur l´autolyse et en corrélation avec sa concentration, soit inhibiteur soit accélérateur.
D´autres essais pour évaluer la capacité autolytique de 34 souches de L. helveticus ont été réalisés in vitro par Lortal et al. (1989) en tampon phosphate (0,1 M, pH 5,8) à 37 °C pendant 16 heures. Cette étude in vitro a montré que l´autolyse est un phénomène souche-dépendant. L´activité autolytique de certaines souches utilisées au cours de ma thèse a déjà été évaluée (Lortal et al., 1989). D´après le taux de lyse (TL), les souches sont divisées en trois groupes : TL≤ 40 % (CIP 615, CIP 103146T, CNRZ 241), 40≤ TL≤50 % (CNRZ 32, CNRZ 303 et ISLC5), TL≥50 % (CNRZ 414).
Valence et Lortal (1995) ont visualisé des autolysines de la souche L. helveticus ISLC5 à l´aide de la technique du zymogramme. En fonction du poids moléculaire apparent sept bandes translucides ont été détectées sur les zymogrammes contenant les cellules de
L. helveticus ISLC5 autoclavées comme substrat (Fig. 4) : une bande à 42,4 kDa, appelée
l´autolysine A, et six bandes vers 30 kDa (29.1, 29.6, 30, 30.8, 31.7 et 32.8), appelées les autolysines B (B1-B6). Une bande supplémentaire de 37,5 kDa a été détectée lorsque le substrat utilisé était des parois cellulaires de L. helveticus ISLC5 traité au SDS.
Fig. 4 Profil électrophorétique (zymogramme) des autolysines détectées chez L. helveticus ISLC5 en phase exponentielle de croissance (1-8 h) par Valence et Lortal (1995).
La lyse in situ a été démontrée dans les fromages commerciaux à pâte pressée cuite, en utilisant la dipeptidase PepD, un marqueur de lyse spécifique de L. helveticus (Valence et al., 1998).
18 L´autolyse et la protéolyse dans du fromage à pâte pressé cuite type Emmental ont été suivies pour deux souches de L. helveticus présentant différentes capacités autolytiques en tampon (Valence et al., 2000). La souche ITG LH1, moins lytique in vitro que la souche ITG LH77, (taux de la lyse respectifs de 34 % et 62 %), a pourtant montré une activité autolytique et protéolytique plus forte dans le fromage que la souche ITG LH77.
Des recherches sur la lysogénie et les conséquences de l´état lysogène sur l´aptitude à la lyse in situ des souches de L. helveticus ont été réalisées par Deutsch et al. (2002b). Il a été montré que 50 % des souches de L. helveticus sont lysogènes, et malgré cela, sont aptes à être utilisées en technologie fromagère. Les cellules bactériennes se lysent lors de la fabrication du fromage dès la fin du pressage, et une libération spontanée de phage a été observée. Le gène de l´endolysine du phage temperé Φ-303, intégré dans la souche lysogène CNRZ 303, a été séquencé, puis le rôle du prophage dans la lyse et son impact sur la protéolyse ont été étudiés. Il a été montré que l´expression du prophage in situ n´était pas responsable de la lyse de
L. helveticus CNRZ 303. Il en a été conclu que la compréhension du phénomène de lyse chez L. helveticus, nécessitait de caractériser les autolysines au niveau moléculaire.
Kenny et al. (2005) ont comparé le potentiel autolytique des 41 isolats de L. helveticus par un criblage in vitro et in situ dans le fromage. Comme Valence et al. (2000), ils ont observé qu’il n´ existait pas de corrélation entre le taux d´autolyse in vitro et le taux d´autolyse in situ. Ils en ont conclu que seule la fabrication fromagère était la méthode ad hoc pour évaluer le potentiel autolytique des souches de L. helveticus. De plus, ils ont par RAPD-PCR regroupé des isolats de L. helveticus en neuf groupes en fonction de la ressemblance génomique. Ils n´ont pas trouvé de similitude dans l´activité autolytique au sein de ces groupes, ce qui montre qu´il n´existe pas de lien entre le taux d´autolyse et la ressemblance génomique.
D´autres essais pour évaluer la capacité autolytique in situ de L. helveticus ont été réalisés par Hannon et al. (2006, 2007). L´utilisation de la souche fortement autolytique L. helveticus DPC 4571 a permis d´accélérer l´affinage du Cheddar (Hannon et al., 2003). Cette souche utilisée pendant la fabrication a permis d´augmenter le niveau de la protéolyse dans le fromage et d´en améliorer les qualités sensorielles (Hannon et al., 2003, Hickey et al., 2007). En 2008, Callanan et al. ont séquencé cette souche L. helveticus DPC 4571, ayant des impacts positif sur l´affinage de fromage, et neuf gènes codant pour les autolysines ont été ainsi annotés, gènes que nous présenterons plus en détail dans le paragraphe 3.1.
19
2
LE MECANISME D´AUTOLYSE
L´autolyse est un phénomène multienzymatique. Elle est basée sur l´action des hydrolases du peptidoglycan endogènes, aussi appelées autolysines, sur le peptidoglycane, constituant essentiel de la paroi bactérienne.
2.1 LES PEPTIDOGLYCANES HYDROLASES (PGHs)
Les hydrolases du peptidoglycan (peptidoglycane hydrolases, PGHs) sont des enzymes situées au niveau de la paroi cellulaire des bactéries à Gram positif. Elles sont capables de couper les liaisons covalentes du peptidoglycane. Les PGHs endogènes, appelées « autolysines » sont impliquées dans la synthèse et les modifications du peptidoglycane. Cela confère un rôle-clé dans la croissance bactérienne et la division cellulaire (Delcour et al., 1999).
2.1.1 Classification des PGHs
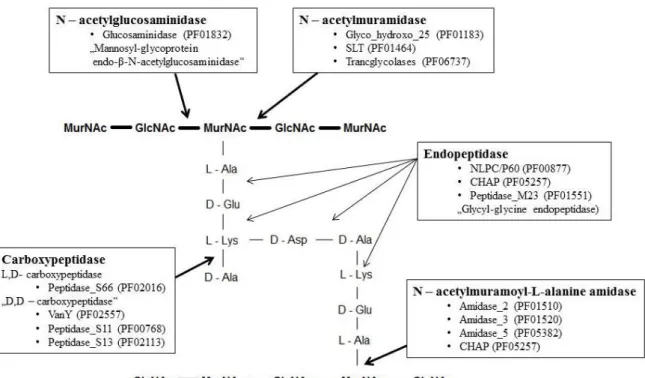
Les PGHs sont classées en trois classes selon la spécificité de coupure au sein du peptidoglycane (Fig. 5) : les glycosidases, les amidases et les peptidases (Delcour et al., 1999, Layec et al., 2008, Vollmer et al., 2008a). Plusieurs PGHs ayant la même spécificité mais différents domaines catalytiques peuvent coexister au sein d´une même bactérie (Layec et al., 2008, Vollmer et al., 2008b).
Les glycosidases hydrolysent les chaines polysaccharidiques composées d´unités alternées d´acide N-acétylmuramique (MurNAc) et de N-acétylglucosamine (GlcNAc) liées par une liaison glycosidique β (1→4). Cette famille inclut les N-acétyl-β-D-muramidases qui
hydrolysent la liaison glycosidique entre le MurNAc et le GlcNAc et les N-acétyl-β-D
-glucosaminidases qui hydrolysent la liaison glycosidique entre le GlcNAc et le MurNAc (Scheurwater et al., 2008). Les N-acétylmuramidases sont divisées en deux sous-groupes : les lysozymes et les transglycosylases. Les lysozymes hydrolysent la liaison glycosidique en produisant un résidu MurNAc sous forme réduite tandis que les transglycosylases forment un résidu 1,6-anhydride de MurNAc.
Seules les N-acétylmuramyl-L-alanine amidases, aussi appelées amidases, hydrolysent la
liaison entre le MurNAc et la L-alanine, premier acide aminé de la chaîne peptidique, et séparent ainsi la partie glycosidique de la partie peptidique du peptidoglycane.
20 Les chaînes peptidiques sont hydrolysées par des peptidases. Selon leur site d´action, les peptidases sont subdivisées en endopeptidases et carboxypeptidases (les D,D-carboxypeptidases et les L,D-D,D-carboxypeptidases) (Delcourt et al., 1999, Courtin et al., 2006).
Les PGHs peuvent encore être divisées selon leur origine génomique en deux groupes : les autolysines et les endolysines.
Les autolysines sont des enzymes bactériennes endogènes tandis que les endolysines sont des enzymes issues des bactériophages. Les autolysines sont responsables de l´induction de l´autolyse. Les endolysines sont des PGHs codées par le double brin d´ADN phagique qui peuvent également causer la lyse. Elles sont particulièrement intéressantes en tant que nouvelles alternatives aux antibiotiques (Parisien et al., 2008, Hermoso et al., 2007, Donovan
et al., 2006, Donovan et al., 2008).
Fig. 5 Représentation schématique des sites de coupure et des domaines catalytiques des PGHs (PF – notation du domaine dans la database Pfam) existantes chez les bactéries à Gram positif (d´après Layec, 2008).
2.1.2 Les autolysines
Comme toute autre protéine, les autolysines sont caractérisées par leur masse moléculaire (MM) et leur point isoélectrique (pI). Les gènes des autolysines code des polypeptides composés de séquences signales (SS), de domaines contenant le site actif (catalytique), et de domaines contenant le site de liaison au peptidoglycane (peptidoglycan-binding site). Il existe
21 des autolysines bi-fonctionnelles ayant deux domaines catalytiques comme par exemple CwlT de Bacillus subtilis (Fukushima, 2008), AltA de Staphylococcus aureus (Oshida, 1995) ainsi que AltL de Staphylococcus lugdunensis (Bourgeois, 2009). Le CwlT a une activité N- acétylmuramidase et DL-endopeptidase tandis que l´AltA et l´AltL ont une activité N- acétylglucosaminidase et N-acétylmyramoyl-L-alanine amidase. Pour s´associer de façon non-covalente au peptidoglycane, les autolysines possèdent des domaines spécifiques, dits de liaison au peptidoglycane, comme par exemple le domaine LysM qui est un domaine ubiquitaire chez les bactéries à Gram positif. Ce motif a une taille caractéristique de 44 à 64 acides aminés. Il peut être présent à l´extrémité C- terminale ou à l´extrémité N- terminale des protéines (Andre et al., 2008, Buist et al., 2008).
Chez les bactéries à Gram négatif, comme E. coli, les autolysines sont localisées dans l´espace périplasmique et dans le cytoplasme. Cinq transglycosylases qui sont des lipoprotéines (MltA, MltB, MltC, MltD et EmtA), une transglycosylase soluble (Slt70), cinq amidases (AmiA, AmiB, AmiC, AmiD et AmpD) et trois peptidases (MepA, PBP4 et PBP7) sont actuellement connues (Vollmer et al., 2008b). Pour une autre bactérie à Gram négatif,
Vibrio anguillarum, une méthode d´identification basée sur la PCR espèce spécifique (Hong et al., 2007) a été proposée employant le gène de N-acetylmuramoyl-L-amidase (amiB).
Hong et al. (2007) a aligné les gènes codant pour la protéine amiB de différentes espèces du genre Vibrio. Il a comparé les séquences redondantes existantes entre les différentes espèces du genre Vibrio sp. ainsi que les séquences différentes et similaires au sein d´une espèce donnée, V. anguillarum. En créant des amorces spécifiques, il a identifié par PCR
V. anguillarum parmi les souches du genre Vibrio sp.. Il a observé que les séquences variables
de genre Vibrio sp. sont remarquablement conservées au sein de l´espèce anguillarum. L´ensemble des souches de cette espèce possèdent donc le même variant du gène. La conservation des autolysines au sein d´une espèce a également été observée chez
Streptococcus pneumoniae (Pagliero et al., 2005).
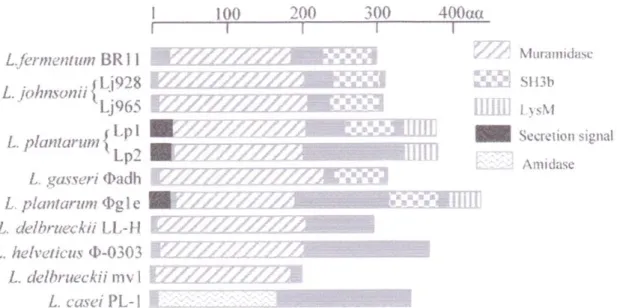
Chez les bactéries à Gram positif, comme S. aureus ou B. subtilis, les autolysines sont localisées au niveau du peptidoglycane et du cytoplasme. Lc. lactis, le modèle le plus exploré des bactéries lactiques, possède cinq gènes de PGHs endogènes. L´autolysine majeure, la N-acétylglucosaminidase, acmA, a été caractérisée par Buist et al. (1995). Elle est constituée de deux domaines : le domaine du site actif au niveau de l´extrémité N-terminale et le domaine de la liaison au peptidoglycane à l´extrémité C-terminale (Steen et al., 2003, Steen et al., 2005). Trois gènes acmB, acmC et acmD ont ensuite été découverts. Il a été montré que ces
22 quatre gènes avait des sites actifs similaires mais qu´ils diffèrent en structure modulaire de protéine (Bolotin et al., 2001 et Huard et al., 2003). Dans le chromosome de Lc. lactis IL 1403 une autre lysine a été mise en évidence, la lysine putative YjgB. Les fonctions des gènes AcmA, AcmB, AcmC, AcmD et YjgB ont été observées in vitro et il a été montré que tous les gènes étaient transcrits chez Lc. lactis MG 1363 (Huard et al., 2004).
Fig. 6 Organisation moléculaire des autolysines de lactocoque (AcmA, AcmB, AcmC, AcmD, YjgB). Les domaines du site actif de la N-acétylmuramidase sont en gris foncé, les domaines de liaison au peptidoglycane « peptidoglycan-binding sites» sont en gris clair, les séquences signales (SS) sont en noir et les autres domaines sont représentés en blanc. (MM- masse moléculaire, pI – point isoélectrique) (Steen et al., 2007).
2.1.2.1 Fonctions des autolysines
Les fonctions des autolysines ont été essentiellement étudiées chez E. coli, bactérie à Gram négatif, chez B. subtilis et S. aureus, bactéries à Gram positif, ainsi que chez Lc. lactis, l´espèce modèle des bactéries lactiques (Vollmer et al., 2008a). Historiquement, les fonctions des autolysines étaient étudiées par délétion des gènes. Des mutants dépourvus d´un ou plusieurs gènes codant pour les PGHs endogènes étaient construits et l´influence de ces délétions sur les caractéristiques phénotypiques et morphologiques était étudiée.
Le fait que les autolysines soient actives en phase exponentielle de croissance suggère qu´elles jouent un rôle important dans la croissance bactérienne, la séparation des cellules et l´assemblage de la paroi cellulaire (Kok et al., 1955, Delcourt et al., 1999, Courtin et al., 2006, Vollmer et al., 2008a). Aucun mutant totalement dépourvu de la totalité des PGHs endogènes n´a jamais pu être obtenu, ce qui démontre le rôle essentiel de ces enzymes dans la croissance bactérienne. Les autolysines sont également impliquées dans la formation du septum de division, une structure pariétale double pré-divisant puis divisant la cellule en deux. Une corrélation entre l´incapacité des cellules à se séparer et une déficience en activité autolytique a été démontrée chez plusieurs espèces bactériennes. Chez B. subtilis, l´amidase
23
LytC, la glucosaminidase LytD et deux endopeptidases LytE et LytF sont impliquées dans la
séparation des cellules filles (Blackman et al., 1998, Margot et al., 1992). L´inactivation du gène Alt, codant pour l´autolysine majeure de S. aureus, conduit également à l´impossibilité des cellules à se séparer. L´addition d´enzyme Alt purifiée dans un milieu contenant ces cellules restaure leur capacité à se séparer (Foster, 1995, Sugai et al., 1995). Dans le cas de
Lc. lactis, l´inactivation du gène AcmA provoque la formation de longues chaînes ce qui
démontre également pour cette bactérie lactique l´implication des autolysines dans la division cellulaire (Steen et al., 2008).
Lors de la croissance cellulaire, les nouveaux précurseurs du peptidoglycane doivent être insérés dans la structure déjà existante. Pour ce faire un renouvellement également appelé « turnover » est nécessaire (Vollmer et al., 2008b). Une rupture provisoire des liaisons covalentes au sein du peptidoglycane existant est nécessaire afin de former de nouveaux sites accepteurs de précurseurs du peptidoglycane. Bien évidemment, une activité incontrôlée des autolysines peut conduire à la destruction de l´intégrité de la paroi cellulaire. Pour éviter l´autolyse, il faut alors que les PGHs endogènes et les peptidoglycane synthétases fonctionnent de concert et que leurs actions soient synchronisées (Antignac et al., 2007,Vollmer et al., 2008b).
Le remodelage de la paroi sous l´action d´un stress environnemental tel qu´un choc osmotique peut conduire à une augmentation de taille incontrôlée de la cellule ou à des modifications de la composition du peptidoglycane. Certaines études ont montrées qu´une paroi stressée était moins sensible aux autolysines qu´une paroi non-stressée. Ainsi chez
S. aureus, une diminution de l´activité autolytique a été observée pour des cellules stressées.
Leurs ponts interpeptidiques au sein du peptidoglycane sont plus courts que ceux des cellules non-stressées (Vollmer et al., 2008b).
Chez des bactéries capable de sporuler telles que B. subtilis, les PGHs endogènes interviennent dans le processus de sporulation. Certaines autolysines sont spécifiquement dédiées à la sporulation elle-même et d´autres interviennent dans l´hydrolyse du PG cortical lors de la germination des spores. Ce phénomène a été mis en évidence par Sekiquchi et al. (1998), qui a montré que l´inactivation de l’amidase (cwlD) chez B. subtilis rendait la souche incapable de germer.
Les PGHs endogènes peuvent également contribuer à la virulence bactérienne. En libérant le contenu cytoplasmique ainsi que des facteurs de virulence, elles génèrent des muropeptides, agents responsables de la réponse inflammatoire et d´effets cytotoxiques tels que le choc