Molecular phylogeny and taxonomy of Testate amoebae (protist) and host-symbiont evolutionary relationships within mixotrophic taxa
159
0
0
Texte intégral
Figure

+7


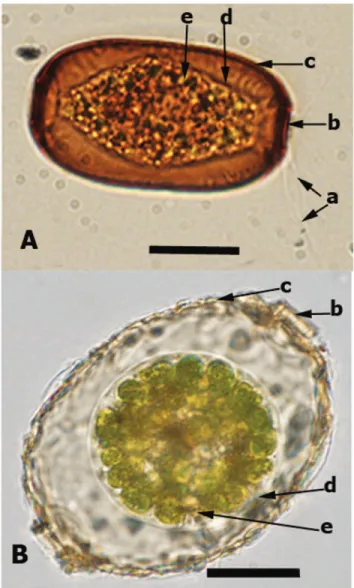
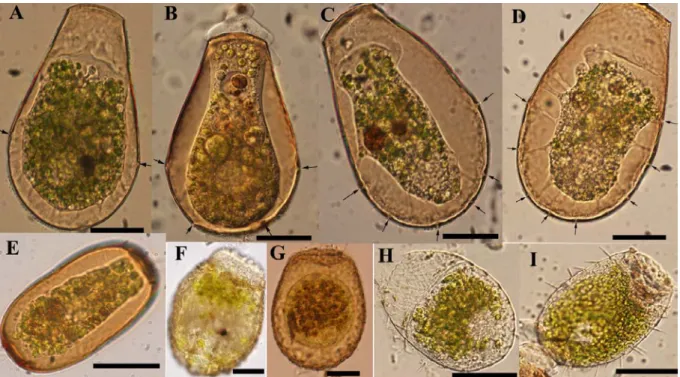
Documents relatifs