Rising Sound Intensity: An Intrinsic Warning Cue Activating the Amygdala
6
0
0
Texte intégral
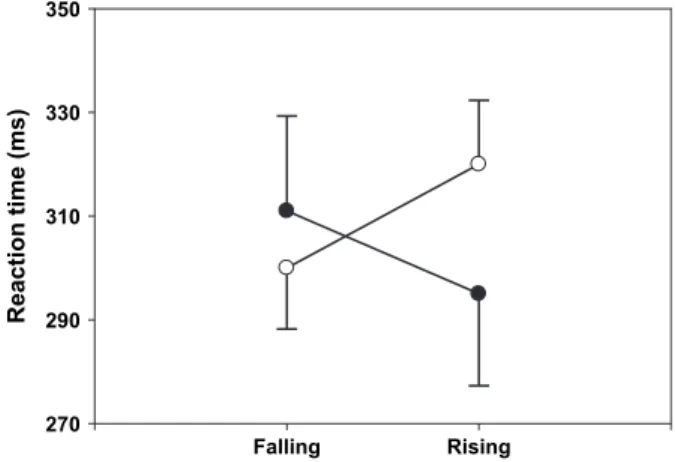
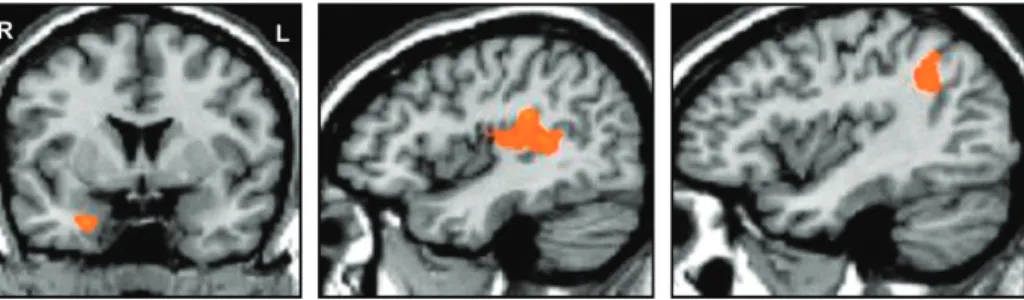
Figure
Documents relatifs