Dynamics of pyrethroid resistance in malaria vectors in southern Benin following a large scale implementation of vector control interventions
Texte intégral
Figure
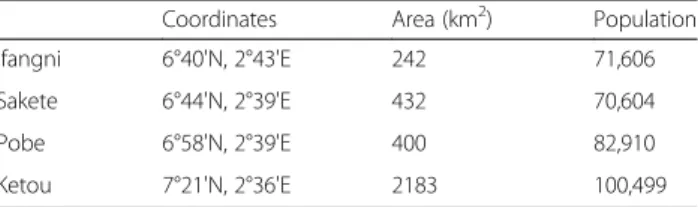
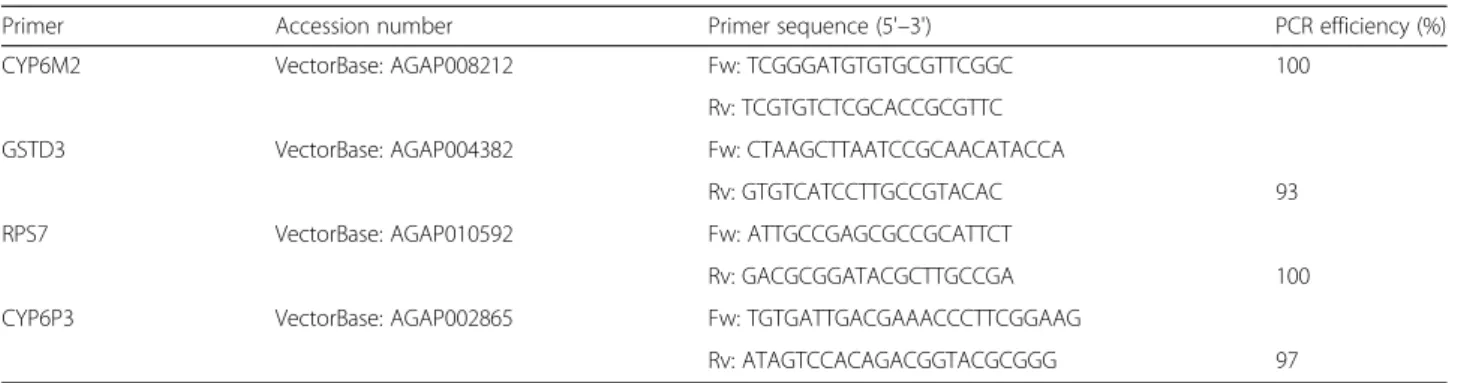

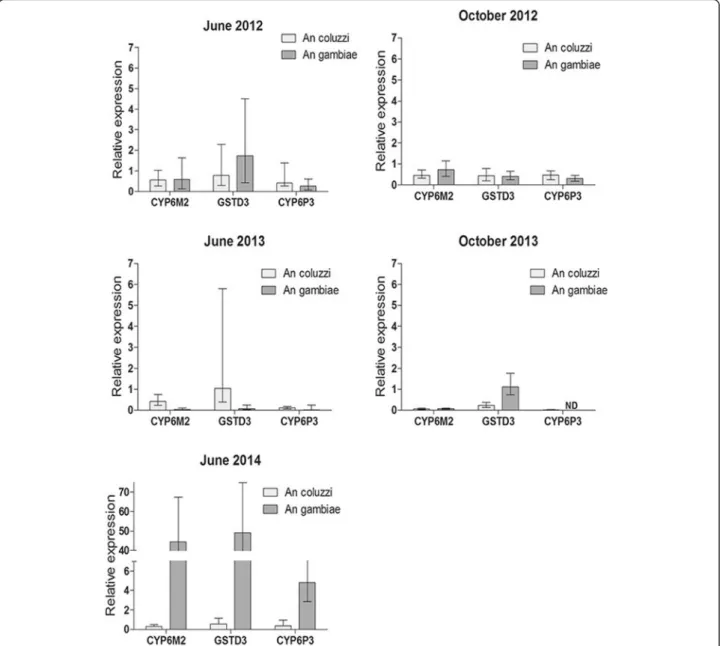

Documents relatifs
As already mentioned, Theorem 2 (Sard for Lipschitz selections) is an extension of [1, The- orem 5] (Morse for Lipschitz selections), albeit not a straightforward one: the proof of
E-V13 is also found at low frequencies on the Anatolian mainland [13] and thus may be useful in teasing apart the relative contributions of Greek colonization (E-V13) from
2 ىلاعت لاق : يدلاو ىلعو يلع تمعنأ يتلا كتمعن ركشا نأ ينعزوأ يبر" لمعا ناو كتمحرب ينلخدأ و هاضرت احلاص "نيحلاصلا كدابع يف ةيلآا لمنلا
Compared to a static landscape, a dynamic landscape clearly produces a large total diver- sity: up to 12.88 species on average (Figure 4, panel A), and up to 24 species in
Djogbenou L, Pasteur N, Bio-Bangana S, Baldet T, Irish SR, Akogbeto M, Weill M, Chandre F: Malaria vectors in the Republic of Benin: Distribution of species and molecular forms of
The low frequency of Kdr gene in BP localities compared with those from CCP and TICP could be due to the fact that in the past these farmers in BP sites used insecticide to
In the present study, first, the efficacy and wash resistance of different materials treated with bendiocarb (a car- bamate approved by the WHO for IRS) at the doses of 100 mg/m 2
These two metals are associated with various chemical fractions of the mine soil sample, as revealed by a five-step, operationally defined sequential chemical extraction procedure
