HAL Id: tel-01775361
https://hal.univ-lorraine.fr/tel-01775361
Submitted on 24 Apr 2018HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Gestion de la construction de la toile chez une araignée
orbitèle, Zygiella x-notata : Perception et utilisation des
informations du milieu
Laurent Thévenard
To cite this version:
Laurent Thévenard. Gestion de la construction de la toile chez une araignée orbitèle, Zygiella x-notata : Perception et utilisation des informations du milieu. Neurosciences [q-bio.NC]. Université Henri Poincaré - Nancy 1, 2004. Français. �NNT : 2004NAN10015�. �tel-01775361�
AVERTISSEMENT
Ce document est le fruit d'un long travail approuvé par le jury de
soutenance et mis à disposition de l'ensemble de la
communauté universitaire élargie.
Il est soumis à la propriété intellectuelle de l'auteur. Ceci
implique une obligation de citation et de référencement lors de
l’utilisation de ce document.
D'autre part, toute contrefaçon, plagiat, reproduction illicite
encourt une poursuite pénale.
Contact : ddoc-theses-contact@univ-lorraine.fr
LIENS
Code de la Propriété Intellectuelle. articles L 122. 4
Code de la Propriété Intellectuelle. articles L 335.2- L 335.10
http://www.cfcopies.com/V2/leg/leg_droi.php
U.F.R. Sciences et Techniques Biologiques
Ecole Doctorale Biologie - Santé - Environnement Département de Formation Doctorale: Neurosciences
Thèse
s.c.o. -
U.H.P.
ANCY 1
iBLIOTHÈOdE DES SCIENCES
P"IP r.'_ !~rrjin Botanique
~..';4· . ~ - :~<ES-NANCY
présentée pour l'obtention du titre de
Docteur de l'Université Henri Poincaré, Nancy-1
en Neurosciences
soutenue publiquement le 27 Mai 2004
par
Laurent THEVENARD
Gestion de la construction de la toile chez une Araignée orbitèle,
Zygiella x-notata :
perception et utilisation des informations du milieu
Président du jury : M. Stéphane FLAMENT Rapporteurs : M. Alain CANARD M. Pierre JOLY Examinateurs : M. Raymond LEBORGNE M. Alain PASQUET
Professeur, Université Henri Poincaré, Nancy I
Professeur, Université Rennes I, Rennes I Professeur, Université Claude Bernard, Lyon I
Maître de conférences, Université Henri Poincaré, Nancy I Chargé de recherche, Université Henri Poincaré, Nancy I
(Directeur de thèse)
Expression et Evolution des Comportements
A Colette et Robert, mes parents...
S.C.o. - U.H.P. NANCY 1
BIBLIOTHÈQUE DES SCIENCES R" '. " " :-"cilrbotanique
Remerciements
Je tiens à remercier ici ceux qui, de près ou de loin, m'ont apporté leur aide et leur soutien lors de la réalisation de ce travail.
Monsieur Alain Pasquet, en tant que Directeur de thèse, et Monsieur Raymond Leborgne, pour m'avoir accueilli dans leur groupe, pour leur patience et les conseils prodigués au cours de ces longues années.
Messieurs Alain Canard et Pierre Joly, ainsi que Monsieur Stéphane Flament, pour aVOIr accepté d'examiner ce travail et de faire partie du jury.
Monsieur Samuel Venner, pour sa collaboration à certaines études.
Madame Marie-Claude Bel-Venner, pour ses remarques constructives sur la partie publiée de ce travail.
Monsieur Christian Vogel, pour son assistance technique et son empressement à rendre servIce.
Les membres de l'ex-Laboratoire de Biologie et Physiologie du Comportement. Tout particulièrement, Mesdames Chantal Roland, Monique Déhaut et Andrée Morel, pour les petites attentions de prime abord insignifiantes mais bien souvent réconfortantes.
Mes amis, pour leurs encouragements et leur sollicitude.
Enfm et surtout, ma famille, pour leur patience, leur soutien, et pour tout ce que l'on ne se dit pas...
Et sans oublier ces chères Zygiella, pour leur compagnie et pour m'avoir ouvert à d'autres
horizons.
Sommaire
Introduction
Il
Chapitre 1
21
Matériel et méthodes
1.1. Modèle biologique 23 1.2. Maintenance au laboratoire 25 1.3. Paramètres mesurés 281.3.1. Caractéristiques des Araignées étudiées 28
1.3.2. Paramètres liés à la construction de la toile 29
1.3.3. Exploitation statistique des données 32
Chapitre II
33
Interactions avec les proies et les congénères dans les conditions naturelles
11.1. Protocoles d'étude 37
II.!.!. Etude de la population naturelle "Gymnase" 37 II.!.2. Etude de la population naturelle "Premier cycle" .43
II.2. Structuration des populations naturelles .45
II.3. Positionnement dans le système retraite-toile et activité des Araignées .49 II.3.!. Evolution de la position des Araignées au cours de la journée .49
II.3.2. Interactions avec les proies 52
II.3A. Construction de la toile géométrique 66
II.3.4.1. Pourcentage de construction de la toile 66
II.3A.2. Distribution du démontage et de la construction de la toile 68
II.3A.3. Caractéristiques de la toile 71
liA. Relations entre les interactions observées et la construction de la toile 75
11.5. Conclusion 87
Chapitre III
95
Construction de la toile en présence de proies et de congénères
IIL1. Influence des proies et des congénères sur la construction de la toile 98
111.1.1. Protocole expérimental 98
III.l.2. Résultats 100
III. 1.2. 1. Caractéristiques des Araignées étudiées 100 III. 1.2.2. Pourcentage de construction de la toile 10 1 III. 1.2.3. Distribution de la construction de la toile au cours du temps 103
III.1.2A. Durée de construction de la toile 105
III. 1.2.5. Caractéristiques de la toile 107
111.1.3. Conclusion 109
111.2. Facteurs de variation pouvant expliquer l'absence d'influence du congénère 112 111.2.1. Influence de la durée de la mise en présence des Araignées 112
III.2.1.1. Protocole expérimental 112
III.2.1.2. Résultats 113
111.2.1.3. Conclusion 115
III.2.2. Influence de l'âge des Araignées sur le comportement de construction 117
111.2.2.1. Protocole expérimental 117
111.2.2.2. Résultats 118
Chapitre IV
123
Informations sensorielles prises en compte par l'Araignée lors de la
construction de la toile en présence de proies et de congénères
IV. 1. Nature des informations sensorielles prises en compte par l'Araignée 126
IV.l.1. Protocoles expérimentaux 126
IV.1.1.1. Généralités 127
IV. 1.1.2. Influence des informations chimiques volatiles 129
IV.1.1.3. Influence des informations tactochimiques+chimiques volatiles 129
IV.1.1A. Influence des informations vibratoires+chimiques volatiles 131
IV.1.2. Résultats 131
IV.1.2.1. Comparaison des groupes "contrôle" des trois protocoles expérimentaux. 133
IV. 1.2.2. Informations sensorielles liées à la présence de proies 137
IV. 1.2.3. Informations sensorielles liées à la présence d'un congénère 143
IV.1.2A. Informations sensorielles liées à la présence simultanée de proies et d'un
congénère 149
IV.1.3. Conclusion 153
IV.2. Réponse de l'Araignée à différentes intensités d'informations liées à la présence de
congénères 159
IV.2.1. Protocole expérimental 159
IV.2.2. Résultats 160
IV.2.3. Conclusion 165
Discussion générale
167
Références bibliographiques
179
Annexes
193
Annexe 1 : Venner, Thévenard, Pasquet& Leborgne (2001) 195
La sélection naturelle favoriserait les animaux les mieux adaptés au milieu dans lequel ils vivent. Le comportement constitue pour les animaux un moyen d'interaction avec leur environnement, et peut ainsi donner prise à la sélection. Des différences comportementales,
pouvant révéler des différences génétiques, peuvent effectivement conduire àdes différences
dans la contribution au pool génétique des générations futures. Les mécanismes de la sélection naturelle favoriseraient les individus se comportant au mieux par rapport aux conditions écologiques, que ce soit dans la recherche d'un abri, dans l'exploitation de la nourriture, dans la défense contre les prédateurs ou les parasites, dans la recherche de partenaires sexuels, la copulation ou l'élevage des jeunes. Les animaux devraient au cours de leur vie adopter les stratégies leur permettant de maximiser leurs chances de survie et la transmission de leurs gènes aux générations futures.
Les anImaux vivent dans un milieu complexe, soumIS à l'influence de facteurs abiotiques - caractéristiques physico-chimiques du milieu - et de facteurs biotiques - proies, partenaires sexuels, compétiteurs conspécifiques et hétérospécifiques pour les ressources du milieu (abris, nourriture, partenaires sexuels...), prédateurs, parasites... Les caractéristiques de ce milieu sont variables dans le temps et dans l'espace, et souvent non-prédictibles pour l'animal: variation des facteurs abiotiques (température, humidité...) et de la disponibilité en nourriture (en qualité et/ou en quantité) entre différentes parcelles ou au cours du temps, variation de la présence des prédateurs et parasites qui s'adaptent eux aussi à la biologie de leurs proies...
Les animaux devraient s'adapter à ce milieu complexe et variable. La plasticité phénotypique
est une solution à ce problème d'adaptation. Elle correspond aux changements du phénotype
présenté par un individu, un génotype, en réponse aux variations de l'environnement. Autrement dit, c'est la propriété d'un génotype à produire différents phénotypes en réponse à différentes conditions environnementales (Pigliucci 2001) ; notons que le comportement est un trait phénotypique parmi d'autres (morphologie, anatomie, physiologie...). Dans ce milieu variable, les individus faisant preuve de plasticité pourraient être avantagés en terme de survie et de reproduction, et être favorisés par la sélection naturelle. Certains auteurs avancent que la plasticité peut être considérée comme un trait ayant une valeur adaptative, cible directe de la sélection naturelle (Via et al. 1995 ; Pigliucci 2001).
A chaque instant les anImaux peuvent s'engager dans une activité, parmi de nombreuses, souvent incompatible avec d'autres: se reposer, se nourrir, parader... Ils peuvent alors avoir le choix entre plusieurs stratégies; par exemple, un prédateur peut chasser à l'affût
ou traquer ses prOIes. Cela signifie dans ce cas qu'ils présentent une certaine plasticité comportementale (une forme de plasticité phénotypique), et qu'ils sont capables de moduler leur comportement en réponse à des situations différentes. De nombreux facteurs peuvent influer sur leur comportement, et agir dans le même sens ou dans des directions opposées générant ainsi un conflit au niveau de l'action à réaliser. La plasticité comportementale des animaux, qui peut leur permettre de répondre à la variabilité environnementale, les place face à une alternative, et les conduit à faire des choix, à prendre des décisions. Afin de maximiser leurs chances de vivre et se reproduire, ils doivent prendre en compte dans leurs processus de prise de décision les variations des différents facteurs environnementaux. Ceci implique qu'ils perçoivent certaines informations les renseignant sur leur environnement. L'adaptation au milieu complexe dans lequel ils vivent nécessite que les animaux soient capables de tenir compte simultanément de plusieurs facteurs, et donc de percevoir et d'intégrer plusieurs types d'informations en même temps.
Les processus de décision peuvent être abordés selon un modèle économique. Le comportement peut être vu comme ayant des coûts et des bénéfices, et les animaux comme étant façonnés par la sélection naturelle pour maximiser le bénéfice net en terme de contribution au pool génétique des générations futures. Dans une situation de choix, les décisions prises par les animaux pourraient résulter de balances entre les coûts et les bénéfices associés à chacune des options, aux différentes activités dans lesquelles ils peuvent s'engager et aux variations des différents facteurs internes et environnementaux. Plusieurs auteurs suggèrent l'existence de tels processus de décision: balances entre la nécessité de remonter respirer à la surface et le comportement de cour chez des Amphibiens (Mc Farland 1977), entre la recherche de nourriture et le kleptoparasitisme chez des Oiseaux (Goss-Custard et al. 1998), entre le risque de prédation et les avantages liés au fourragement, aux relations sociales ou à la reproduction mises en évidence dans divers taxons (Insectes, Arachnides, Crustacés, Mollusques, Poissons, Amphibiens, Reptiles, Oiseaux, Mammifères) (synthèse dans Lima & Dill1990 ; Rayor& Uetz 1990, 1993 ; Arenz& Leger 2000; Rohr& Madison 2001).
Dans les différents groupes biologiques, des études ont montré que les animaux tenaient compte des facteurs du milieu dans leurs décisions: des travaux ont ainsi mis en évidence l'influence du risque de prédation (synthèse dans Lima & DilI 1990), des congénères (e.g. Muller 1998 ; Wiackowski & Staronska 1999; Galef & Giraldeau 2001) ou de la disponibilité en nourriture/proies du milieu (e.g. Jacobs 1992; Martin et aL 2003) sur les décisions relatives au fourragement, au comportement anti-prédateur, aux relations sociales ou à la reproduction. La plupart des études se sont cependant limitées à une approche
monofactorielle, ne testant la réponse des animaux qu'aux variations d'un seul facteur à la fois. Peu de données mettent clairement en évidence les capacités des animaux à prendre en
compte simultanément plusieurs facteurs environnementaux (e.g. Holbrook & Schmitt 1988 ;
Hazlett 2000; Rohr & Madison 2001): Rohr & Madison (2001) ont ainsi montré qu'en
présence simultanée d'informations sensorielles liées aux prédateurs et aux partenaires
sexuels, les mâles Notophthalmus viridescens (Caudata, Salamandridae) présentent une
réponse intermédiaire entre l'attraction pour l'odeur des femelles et la répulsion vis à vis des informations liées aux prédateurs, ce qui traduit une prise en compte des deux types d'informations par ces mâles. Chez les Araignées, l'étude de Riechert (1985) sur la sélection
de l'habitat chez Agelenopsis aperta (Araneae, Agelenidae) en fonction de la température et
de la présence de proies fait figure d'exception; elle montre qu'en réponse aux variations de chacun des facteurs testé isolément les Araignées choisissent les sites du milieu fournissant des proies et une certaine température, mais que, contrairement à ce que l'on pouvait attendre, lorsque les deux facteurs sont testés simultanément les Araignées ne sélectionnent pas les sites fournissant à la fois les proies et la température préférentielle mais ceux fournissant soit l'un soit l'autre. L'une des hypothèses avancées pour expliquer ce résultat est que le système nerveux central des Araignées ne serait peut-être pas capable de résoudre ce type de problème nécessitant d'intégrer - i.e. trier, évaluer, comparer, mémoriser - simultanément les influences de multiples facteurs. Les capacités cognitives des Araignées pourraient limiter leurs capacités de gestion à la prise en compte d'un seul facteur à la fois.
D'une manière générale, peu d'auteurs se sont jusqu'à maintenant intéressés aux capacités
cognitives des Araignées et peu de données existent (Jackson 1995). L'étude du
comportement prédateur de certaines espèces de Salticidae apporte cependant quelques informations sur ces capacités. Ces Araignées errantes repèrent visuellement leurs proies (Forster 1979), et lors de leurs approches elles peuvent réaliser des détours et perdre de vue
les proies, mais, néanmoins réussir à les capturer (Tarsitano & Jackson 1992, 1994, 1997 ;
Jackson & Stimson Wi1cox 1993). Ces détours impliquent que ces Araignées mémorisent la
localisation des proies repérées, et, ceci suggère qu'elles seraient capables de "planifier" leur
parcours pour atteindre ces proies avant de commencer leur approche (Tarsitano & Jackson
1994).
Afin d'étudier les règles sous-jacentes aux processus de décision, il faut donc un modèle animal présentant un comportement variable en fonction des conditions environnementales. Il faut également que ce comportement influe directement sur la fitness, pour donner prise à la
sélection naturelle. De plus, il doit être facilement quantifiable en terme de coûts et de bénéfices.
Chez les Araignées, la prise alimentaire a un effet positif direct sur la croissance et la production d'une descendance (palanichamy 1984; Fritz & Morse 1985 ; Vollrath 1987a), par conséquent, les décisions relatives au fourragement devraient affecter leur fitness. Chez les Araignées à toile, le comportement prédateur peut se décomposer en quatre étapes successives: le positionnement de l'Araignée dans le milieu, la construction de la toile, la capture des proies interceptées et retenues par la toile, et l'ingestion de ces proies. La construction de la toile est une étape importante du fourragement puisque la toile est pratiquement l'unique moyen pour ces Araignées de se procurer l'énergie nécessaire à leur vie et leur reproduction. La coïncidence temporelle entre la mise en place d'un piège efficace et la présence des proies potentielles dans le milieu, les caractéristiques de la toile construite, et l'efficacité du comportement de capture des Araignées déterminent en partie leur succès de fourragement (Buskirk 1975a ; Chac6n & Eberhard 1980; Nentwig 1983 ; Craig & Bernard 1990 ; Higgins & Buskirk 1992 ; Ridwan 1993 ; Ap Rhisiart& Vollarth 1994 ; Barghusen et al. 1997; Tso 1998). Les décisions relatives à la toile et sa construction devraient donc affecter la fitness des Araignées.
La toile est une structure variable. Même si des études comparatives ont pu implicitement laisser penser que la variation dans le design de la toile résidait essentiellement entre les espèces (Risch 1977; Ramousse& Le Guelte 1984), de nombreux travaux ont révélé que les variations entre Araignées d'une même espèce étaient une généralité (synthèse dans Eberhard 1990). Certaines études ont également mis en évidence une variabilité au niveau individuel, entre les constructions successives d'une même Araignée (Higgins 1990; Sandoval 1994 ; Schneider& Vollrath 1998 ; Venner et al. 2000).
A chacune des étapes du fourragement, les Araignées doivent tenir compte dans leurs choix de divers facteurs internes et externes: ainsi le choix du site d'installation peut dépendre de la température du milieu (Riechert 1985 ; Henschel et al. 1992), ou de la présence de proies (Riechert 1985) ou de congénères (Pasquet 1984). La construction de la toile (moment et rythme de construction, caractéristiques de la toile...) peut être influencée par l'état interne des Araignées - e.g. croissance, régénération de segments de pattes, alimentation, réserves de soie, production d'œufs, expérience - (Witt & Baum 1960; Benforado & Kistler 1973 ; Vollrath 1987b; Eberhard 1988; Higgins 1990; Higgins & Buskirk 1992; Pasquet et al. 1994; Sherman 1994; Vollrath & Samu 1997 ; Vollrath et al. 1997; Blackledge 1998 ; Heiling &
Herberstein 1998 ; Venner et al. 2000), et les conditions environnementales - e.g. lumière, température, humidité, vent, gravité, supports, présence de proies ou de congénères - (Szlep
1958 ; Le Guelte & Ramousse 1979 ; Hieber 1984 ; Buskirk 1986 ; Eberhard 1987 ; Gillespie
1987 ; Kremer et al. 1987 ; Leborgne & Pasquet 1987a ; Pasquet et al. 1994 ; Sandoval 1994 ;
Craig et al. 1996 ; Barghusen et al. 1997 ; Vollrath et al. 1997 ; Jakob et al. 1998 ; Leborgne
et al. 1998 ; Schneider & Vollrath 1998). Dans la construction du piège, les Araignées font
donc preuve d'une certaine plasticité comportementale qui leur permet d'adapter leur toile à la situation. Elles tiennent compte également de différents facteurs, tels que les caractéristiques
des proies piégées par la toile (Pasquet & Leborgne 1990 ; Sébrier et al. 1994) ou des proies
précédemment consommées (Sébrier et al. 1994), dans le choix des proies capturées et ingérées.
La construction de la toile présente des bénéfices puisqu'elle permet l'acquisition d'énergie par la capture des proies. A ces bénéfices sont associés des coûts en temps et en énergie: temps nécessaire à la construction de la toile, et énergie nécessaire d'une part à la synthèse des protéines constitutives de la soie et d'autre part à l'activité motrice lors de la mise en place des fils de soie. Les facteurs environnementaux peuvent faire varier les coûts et bénéfices de la construction, par exemple: en fonction de la disponibilité en proies du milieu la prise alimentaire peut être plus ou moins importante, ou en fonction de la température ambiante (dont dépend l'activité enzymatique) les coûts et temps de production et de pose de la soie peuvent être plus ou moins grands. Dans un cadre adaptatif, les Araignées devraient prendre en compte ces coûts et bénéfices, et leur investissement dans la construction devrait refléter cette prise en compte. L'investissement des Araignées à toile dans le fourragement se matérialise directement dans les caractéristiques des toiles construites; si bien que la mesure de ces caractéristiques peut nous permettre d'étudier l'influence de l'environnement sur le comportement des Araignées, ou encore les capacités des Araignées à répondre aux variations de leur environnement.
Les modifications comportementales en réponse aux variations environnementales indiquent une perception par les Araignées d'informations sur leur environnement. Les Araignées peuvent percevoir des informations sensorielles de nature différente: visuelle,
chimique, tactochimique, vibratoire. La nature des informations perçues dépend de
l'organisation sensorielle des Araignées. Ainsi, la vue ne semble jouer un rôle important que
informations de nature chimique et vibratoire peuvent être mises en jeu dans le fourragement, et la communication entre congénères et entre partenaires sexuels (synthèse dans Richman &
Jackson 1992 et Uetz& Roberts 2002).
Chez les Araignées tisseuses, la toile est le principal moyen d'interaction avec l'environnement. Sa structure et ses caractéristiques physiques et biochimiques assurent la transmission d'informations aux récepteurs sensoriels de l'Araignée; la toile constitue une extension de l'espace sensoriel de l'Araignée qui peut émettre et percevoir des informations à partir de cette structure (Witt 1975). La toile peut ainsi être le support d'informations chimiques déposées par l'Araignée, et peut transmettre des signaux vibratoires. Mais, les Araignées tisseuses peuvent aussi interagir avec leur environnement sans utiliser leur toile, par l'émission et/ou la perception d'informations chimiques à distance (volatiles) ou de contact (tactochimiques), et de vibrations se propageant dans l'air. Chez les Araignées à toile, le rôle des informations de nature chimique a essentiellement été abordé dans le cadre de la reproduction et du rapprochement des sexes: les femelles peuvent attirer les mâles et déclencher le comportement de cour de ceux-ci via l'émission de phéromones volatiles ou le dépôt de phéromones au niveau de la soie (fils de cheminement ou toile) (Olive 1982; Roland 1984 ; Miyashita & Hayashi 1996); cependant peu de ces phéromones ont été identifiées (Schulz & Toft 1993; Papke et al. 2001). Des informations de nature chimique, des phéromones en l'occurrence, interviennent aussi dans les relations sociales et notamment dans la tolérance réciproque chez les Araignées sociales (Krafft 1970, 1975). Il ya apparemment peu de données sur l'utilisation d'informations chimiques dans le cadre de la prédation chez les Araignées à toile; le seul exemple trouvé concerne les Araignées du genre Mastophora (Araneae, Araneidae), dont la toile se limiteà un fil de soie vertical où bout duquel pend une gouttelette gluante, et qui attirent leurs proies, des phalènes mâles (Lepidoptera, Noctuidae), en produisant une substance volatile qui mime la phéromone sexuelle attractive des femelles des espèces proies (Eberhard 1977 ; Stowe et al. 1987). Par contre, les Araignées tisseuses peuvent détecter leurs proies, s'orienter dans leur direction et les capturer à l'aide des vibrations émises par ces proies lors de leurs déplacements et transmises dans l'air (Riechert 1985) ou lorsqu'elles se débattent dans la toile qui les a piégées (Witt 1975 ; KHirner& Barth 1982) ; ces dernières vibrations peuvent également permettre aux Araignées de discriminer différents types de proies (Suter 1978). Des signaux vibratoires sont également utilisés par les mâles lors de leur approche des femelles et de leur parade sexuelle, transmis par la toile des femelles (Witt 1975). Des informations de nature vibratoire entrent aussi en jeu dans les interactions agonistiques chez les Araignées coloniales (Buskirk 1975b), et dans la tolérance réciproque chez les Araignées sociales (Krafft 1975).
Les Araignées à toile interagissent donc avec leur environnement par le biais de multiples canaux sensoriels (chimique, tactochimique et vibratoire). Ces informations de nature différente peuvent être utilisées individuellement, mais la détection des proies ou des prédateurs et la communication entre congénères pourraient impliquer simultanément différents modes sensoriels, comme des études récentes l'ont montré chez des Araignées
errantes du genreSchizocosa(Araneae, Lycosidae) (Uetz & Roberts 2002).
Les Araignées à toile sont donc un modèle biologique idéal pour traiter des problèmes
de prise en compte d'informations multiples simultanées dans la gestion du comportement. Ces Araignées vivent dans un milieu complexe avec lequel elles interagissent essentiellement via leur toile. Celle-ci est le reflet de leur comportement prédateur, et elle est indispensable à leur survie et leur reproduction. La construction de la toile est un comportement variable, soumis à l'influence de nombreux facteurs, et, qui de plus est aisément quantifiable. Les Araignées devraient être capables de tenir compte des variations des différents facteurs environnementaux afin d'adapter leurs pièges à la situation.
Nous avons choisit d'aborder ce type de problème chez Zygiella x-notata (Clerck)
(Araneae, Araneidae), une espèce d'Araignée à toile géométrique vivant à proximité de l'homme et de ses habitations, généralement considérée comme solitaire. Différentes études, réalisées sur des populations naturelles de femelles adultes colonisant les fenêtres de bâtiments universitaires, ont cependant mis en évidence l'existence d'une répartition de type
agrégatif des Araignées le long des fenêtres (Leborgne & Pasquet 1987a; Kremer 1989a).
Kremer (1989b) a confirmé ces résultats en montrant que la répartition spatiale de type agrégatif des Araignées dans ce type de biotope se retrouvait à chaque stade de développement, de l'émergence du cocon au stade adulte. Il semble que ni les facteurs physiques (supports d'accueil, conditions climatiques) ni la disponibilité en proies du milieu
n'expliquent cette répartition des Araignées (Pasquet & Leborgne 1986). Ces auteurs
suggèrent que les relations intraspécifiques pourraient être à l'origine de cette répartition; cette hypothèse étant soutenue par la mise en évidence au laboratoire des effets attractifs de la
soie de congénères (Leborgne & Pasquet 1987b). Néanmoins, même si la disponibilité en
proies ne peut à elle seule expliquer la répartition agrégative des Araignées, elle peut influer
sur leur positionnement dans le milieu, comme semble le démontrer l'installation
préférentielle des Araignées sur les sites les plus riches en proies (Ridwan 1993). Proies et
x-Les études antérieures des populations naturelles ont également révélé l'existence de contraintes liées au positionnement agrégatif des Araignées; ce type de structure de la population entraîne pour certains individus des contraintes exercées par leurs congénères, qui se traduisent par une réduction volontaire de la taille du piège pour une partie de la population (Leborgne & Pasquet 1987a; Kremer 1989a). Par ailleurs, au laboratoire, Kremer et al. (1987) et Pasquet et al. (1994) ont respectivement montré que la présence d'un congénère ou la présence de proies conduisait à une diminution de la taille des toiles construites. Proies et congénères ont donc un effet sur la construction de la toile chez Zygiella x-notata.
Nous cherchons à répondre, chez Zygiella x-notata, aux deux questions suivantes: d'une part, est-ce que l'Araignée est capable de prendre en compte simultanément des informations provenant de proies et de congénères, et quelle est sa réponse? - en nous fixant la construction de la toile comme comportement révélateur de la prise en compte; d'autre part, quelle est la nature des informations pertinentes pour l'Araignée?
Ce rapport est divisé en quatre chapitres. Le matériel et les méthodes d'étude utilisés au cours des différents travaux sont présentés dans le premier chapitre; cependant certaines particularités protocolaires et méthodes d'analyse sont précisées dans les chapitres concernés. Le second chapitre expose les observations faites sur des populations naturelles de Zygiella
x-notata qui ont permis de déterminer la nature des interactions ayant lieu entre Araignées, et
entre Araignées et proies, ainsi que leurs conséquences sur le comportement de construction. Dans le chapitre ID, nous avons testé la réponse des Araignées à la présence à la fois de proies et congénères lors de la construction de la toile. Le chapitre IV regroupe une série d'expériences réalisées pour déterminer la nature des informations sensorielles prises en compte par les Araignées dans le cadre de la construction de la toile en présence des deux facteurs proies et congénères.
Chapitre 1
1.1. Modèle biologique
Zygiella x-notataest une Araignée à toile géométrique de la famille des Araneidae, dont
la zone de répartition est quasi mondiale (Levi 1974). Elle se rencontre communément à proximité des habitations humaines : typiquement sur le pourtour des fenêtres, mais aussi sur la végétation des jardins, des haies ...
La durée de vie de Zygiella x-notata est de 8 à
la
mois (Juberthie 1954). Les jeunes Araignées émergent des cocons au printemps, se regroupent sur un édifice soyeux commun, avant de se disperser et de mener une vie solitaire. Elles atteignent le stade adulte au cours de l'été, mâles et femelles sont alors identifiables. Les femelles pondent en Septembre-Octobre, déposant généralement leurs œufs dans plusieurs cocons (2 ou 3 le plus souvent) ; cocons à l'abri desquels les larves passent l'hiver. Les mâles disparaissent peu après s'être reproduit, alors que certaines femelles peuvent survivre jusqu'à l'été suivant.La taille des femelles adultes varie selon les auteurs de 6 à 8 mm (Gertsch 1964; Levi 1974 ; Hubert 1979; Kremer 1989a). Le poids moyen et la longueur totale céphalothorax-abdomen moyenne obtenus à partir des mesures des femelles adultes prélevées sur le terrain dans le cadre de nos travaux sont présentés dans le Tableau 1 ; ces deux caractéristiques physiques des Araignées présentent une forte corrélation entre elles.
Tableau 1 : Moyenne (et Erreur Standard), et corrélation entre le poids et la longueur totale de femelles adultes de
Zygiella x-notataissues de populations naturelles
N X SE TestZ Poids (mg) Longueur (mm) 1399 428 51,0 6,81 0,7 0,06 N R2 Z P 428 0,92 39,44 <0.0001
Les mesures ont été prises au moment de la récolte des Araignées sur le terrain.
Figure 1 : Photographie d'une toile géométrique deZygiella x-notata
Légende: Re=Retraite, Mo=Moyeu, SI=Secteur libre, Fa=Fil avertisseur, Fc=Fil de cadre, Ra=Rayon, Sc=Spirale de capture, Zv=Zone vide.
Zygiella x-notata est une Araignée active pendant la période nocturne: le jour elle se trouve le plus souvent dans sa retraite conique ouverte à ses deux extrémités et construite sur le support, tandis que la nuit elle se tient fréquemment au centre de sa toile ou moyeu. Cette
toile est d'ordinaire reconstruite quotidiennement en fin de période nocturne (Ramousse & Le
Guelte 1984). Au stade adulte, les mâles ne construisent plus de toile mais peuvent utiliser la toile des femelles pour capturer des proies et s'en nourrir (observations personnelles sur le terrain).
La toile de Zygiella x-notata est une toile géométrique de forme elliptique caractérisée, en
général, par la présence d'un secteur libre dépourvu de spire, où passe un fil avertisseur reliant la retraite au moyeu de la toile (Fig. 1). Le moyeu constitué de soie sèche est séparé de la spirale de capture gluante par une zone vide sans spire. Nous pouvons distinguer quatre grandes étapes dans la construction de la toile: elle débute par la mise en place du cadre ancré au support et des rayons qui stabilisent l'ensemble de la structure, puis l'Araignée construit une spirale provisoire de soie sèche de l'intérieur vers l'extérieur, spirale qui est remplacée de suite par la spirale de capture gluante construite de manière centripète, la construction
s'achevant par le remodelage du moyeu qui pourrait correspondre à un réajustement des
tensions dans la toile. Une description précise de la construction de la toile chez Zygiella
x-notataa été faite par Le Guelte (1967).
La présence caractéristique d'un secteur libre et d'un fil avertisseur aide à la détermination de
cette espèce dans la nature. Cependant certaines femelles adultes peuvent construire des toiles sans secteur libre, notamment lorsque la retraite n'est pas dans le plan de la toile (observations personnelles).
1.2. Maintenance au laboratoire
Les Araignées étudiées au laboratoire sont des femelles adultes issues de populations
naturelles de l'Est de la France. Après leur capture ces Araignées sont placées
individuellement dans des boîtes en plastique (1 O,Ox7,Ox2,5 cm) munies sur le couvercle d'une ouverture couverte d'une fine gaze assurant une aération constante (Fig. 2). Les Araignées ne peuvent pas construire de toile géométrique dans ces boîtes, mais déposent un tapis de soie qui leur permet de capturer les proies qui leur sont distribuées. L'alimentation des
Araignées a évolué au cours des années suite à la mise en place d'un élevage de mouches,
Calliphora vomitaria (Diptera, Calliphoridae), au sein du laboratoire. Ainsi, les Araignées
Figure 2 : Schéma d'une boîte d'élevage
+
Ei C) <ri N~ ~ ./f ....c?
~~ ~•... ~ 10,0 cm ~Figure3 :Schéma d'une enceinte d'élevage
~
.../1f
c:~ ~ ",1>-' ~.... ~ 54,Ocm ~utilisées pour les expériences décrites au Chapitre III reçoivent chacune deux larves de
grillon, Gryllus bimaculatus (Orthoptera, Grillidae), dont la taille équivaut à la moitié de la
taille des Araignées, ceci deux fois par semaine; alors que les Araignées utilisées pour les
expériences suivantes sont nourries d'une mouche, Calliphora vomitaria, par semaine. Un
apport en eau est assuré par trois pulvérisations par semaine à travers la gaze du couvercle de la boîte.
La descendance des femelles récoltées sur le terrain est également utilisée, ce qUi permet de poursuivre les études expérimentales après la disparition des femelles adultes dans la nature. Les cocons pondus par les femelles conservées dans les boîtes d'élevage sont collectés et placés dans des enceintes en verre (54,Ox54,Ox87,O cm) pouvant accueillir plusieurs cocons (Fig. 3). Chaque enceinte est constituée de quatre vitres dont deux sont amovibles afm de permettre les différentes interventions inhérentes à l'élevage, leur base repose sur un cadre en bois munis d'un tiroir permettant le nettoyage et l'introduction de la nourriture, tandis que la partie supérieure des enceintes est coiffée d'un petit cadre de bois amovible couvert d'une ftne gaze. L'aération à l'intérieur des enceintes est assurée par la circulation de l'air via les ouvertures recouvertes de gaze de la base et de la coiffe.
Les cocons sont déposés sur les branchages disposés dans chaque enceinte. Après leur émergence, les jeunes Araignées construisent des toiles utilisant montants et branchages
comme supports, et peuvent ainsi capturer les drosophiles (Diptera, Drosophilidae)
régulièrement introduites dans l'enceinte. De l'eau est régulièrement vaporisée pour maintenir l'humidité ambiante. Une fois le stade adulte atteint, les Araignées sont retirées des enceintes
et placées individuellement dans des boîtes d'élevage dans les conditions décrites
précédemment.
L'ensemble des boîtes et des enceintes d'élevage est placé dans une salle où règnent des conditions abiotiques constantes: température de 22-24 oC, cycle d'éclairage de 12 heures par jour inversé par rapport à la lumière naturelle avec des périodes diurne et nocturne
1.3. Paramètres mesurés
1.3.1. Caractéristiques des Araignées étudiées
Au laboratoire, afm d'éviter que des facteurs autres que ceux que nous testons n'influent sur le comportement de construction des Araignées, nous sélectionnons dans l'élevage les Araignées selon certains critères.
Ainsi les Araignées qui ont pondu dans les deux semaines précédant l'expérience ne sont pas sélectionnées; la production des œufs entraînant des variations importantes des caractéristiques des toiles construites durant les périodes précédant et suivant la ponte (Higgins 1990; Sherman 1994).
Nous instaurons un critère d'homogénéisation des Araignées au nIveau alimentaire; des différences de l'état alimentaire entre Araignées pouvant entraîner des différences du comportement de construction (Benforado & Kistler 1973 ; Higgins & Buskirk 1992 ; Pasquet et al. 1994; Sherman 1994; Vollrath & Samu 1997; Blackledge 1998). Rappelons que l'alimentation des Araignées est modifiée au cours des années suite à la mise en place d'un élevage de mouches (cf. 1.2.). Une semaine avant l'expérience, les Araignées préparées pour les expériences du Chapitre III reçoivent trois larves de grillon (Gryllus bimaculatus) en une seule fois, et les Araignées préparées pour les expériences suivantes reçoivent une mouche
(Calliphora vomitaria). Pour les expériences ne sont retenues que les Araignées ayant
consommé soit au moins deux des larves de grillon, soit la mouche.
Pour les mêmes raisons d'influence éventuelle d'événements étrangers à notre protocole entrâmant des modifications de l'état interne des Araignées, toute Araignée qui pond au cours de l'expérience ou qui capture et consomme les proies et/ou congénères pouvant être mis en sa présence lors de l'expérience est exclue de l'analyse des données.
Lors des différentes expériences, le poids des Araignées est déterminé par pesée à 0,1 mg près (Balance Sartorius Basic BAUOS). La longueur totale céphalothorax-abdomen des Araignées est mesurée à l'aide d'un pied à coulisse électronique à 0,05 mm près.
Les caractéristiques physiques des Araignées, notamment le poids et la taille, étant connues pour influencer la taille et la structure des toiles géométriques (Witt & Baum 1960 ; Eberhard 1988 ; Higgins & Buskirk 1992), les Araignées sont réparties selon leur poids et leur longueur de façon homogène dans les différents groupes expérimentaux.
Dans le cadre de certaines expériences menées au laboratoire, deux Araignées sont introduites et maintenues dans un même cadre clos pendant quelques jours. Un marquage sur la zone postéro-dorsale de l'abdomen des Araignées, à l'aide de peinture à maquette de couleur différente (Revell, Allemagne), permet la reconnaissance individuelle des Araignées. Toujours dans un souci d'homogénéité, pour ces expériences toutes les Araignées testées sont marquées qu'elles appartiennent à un groupe expérimental où deux congénères sont introduits dans un même cadre ou à un groupe (tel que le groupe contrôle) où une seule Araignée est introduite dans un cadre.
Pour l'étude de populations naturelles de Zygiella x-notata, le marquage des Araignées est
plus délicat à réaliser. Cette technique nécessite effectivement de pousser délicatement l'Araignée hors de sa retraite, du moins suffisamment pour pouvoir atteindre et marquer l'abdomen d'une fine touche de peinture. Ce dérangement entraîne le départ de quelques Araignées qui quittent leur retraite et se réinstallent sur un autre site du milieu, peu de temps après le marquage ou au cours de la nuit suivante. Ceci nous impose donc d'attendre quelques jours après le marquage avant de commencer les observations, le temps que les Araignées dérangées se stabilisent dans le milieu.
1.3.2. Paramètres liés à la construction de la toile
Au laboratoire, les conditions de température et de luminosité régnant lors de
l'expérimentation sont identiques aux conditions d'élevage. L'inversion du cycle
lumière/obscurité par rapport aux conditions naturelles facilite l'observation des Araignées actives pendant la période nocturne. Le dispositif expérimental de base est constitué d'un cadre en bois (SO,OxIO,OxSO,O cm) fermé par deux vitres (Fig. 4). Les Araignées sont
introduites dans les cadres lors de la transition nuit-jour (1:0), individuellement ou en présence
de proies et/ou de congénères selon les groupes expérimentaux. Les cadres sont répartis au hasard dans la pièce d'expérimentation afin d'éviter tout effet lié à l'hétérogénéité de la pièce. Les expériences se déroulent généralement sur 72 heures (sauf précisions ultérieures), et
débutent à 1:0. Les observations commencent dès cet instant et sont effectuées toutes les
demi-heures pendant les premières 24 demi-heures, et en continu quand une Araignée est en cours de construction. Ensuite deux observations sont faites à 48h et 72h.
Figure 4: Schéma d'un cadre expérimental
/v
C ).. (7:). . C7)/
®
0
/
V
El u o o' li")..
/f . :.. 10,0 cm ~ 50,Ocm ~Tableau 2 : Corrélations entre différents paramètres des toiles de Zygiella x-notata Maille Distance entre
spire externe et spire interne (R.Jo-Ri.Jo) Nombre de tours de spires (NJo) Nombre de rayons Diamètre 0,07
*
0,64**
0,37**
0,48**
Nombre de rayons 0,00 0,30**
0,32**
Nombre de tours 0,09*
0,51**
de spires (NJo) Distance entre spireexterne et spire interne 0,15
**
(Ra.lo-Ri.Jo)
Les mesures ont été prises sur des toiles construites au laboratoire par des Araignées placées individuellement dans des cadres expérimentaux. Carré du coefficient de corrélation(R~et probabilité statistique (Test Z) : N=66,
*
P<0.05,**
P<0.005.Au cours de ces observations, nous nous intéressons uniquement à la première toile construite par chaque Araignée; certaines Araignées construisant plusieurs toiles successives au cours de l'expérience. Le nombre d'Araignées construisant une toile est relevé, ainsi que deux paramètres temporels de la construction de la toile: le moment de la construction définit
comme le temps écoulé entre 1:0 et la mise en place des premiers rayons, ainsi que la durée de
la construction correspondant à l'intervalle de temps entre la mise en place des premiers rayons et le remodelage du moyeu marquant la fin de la construction de la toile géométrique. Différents paramètres sont mesurés sur les toiles construites: le diamètre entre les deux spires captrices les plus éloignées de la toile géométrique, et le nombre de rayons. Nous relevons
également l'ensemble des mesures nécessaires à l'estimation de la longueur de fil de soie
investit par l'Araignée dans la spirale de capture à l'aide de la fonnule mathématique (CTLvh)
mise au point par Venner et al. (2001) (Annexe 1), à savoir: le diamètre entre les spires les
plus internes (D;), le diamètre entre les spires les plus externes (Do), et le nombre de tours de
spire (N) sur les axes horizontal et vertical (h,v), ainsi que la base (B) et la hauteur (H) du secteur libre (Fig. 1 et Tab. 1 de l'Annexe 1).
A partir de certaines des mesures nécessaires à la fonnule précédemment citée, nous estimons la taille moyenne de la maille de la toile. La maille d'une toile géométrique est délimitée par deux rayons et deux spires captrices. Nous estimons sa taille par le calcul de la distance
moyenne entre deux tours successifs de la spirale de capture. Chez Zygiella x-notata, la partie
de la toile géométrique située sous le moyeu est généralement plus grande que la partie supérieure (Le Guelte 1966), et doit vraisemblablement être la portion de la toile la plus impliquée dans la capture, puisqu'elle devrait intercepter plus de proies (Eberhard 1990). Nous choisissons donc d'estimer la taille de la maille sur le plus grand secteur (portion de la toile comprise entre deux rayons) de la partie inférieure de la toile. La fonnule utilisée est la
suivante (cf. Fig. 1 et Tab. 1 de l'Annexe 1 pour les légendes) : (R,.lo-Ri.lo)/Nlo
Tout comme Leborgne & Pasquet (1987a) l'ont mis en évidence sur des populations
naturelles, il existe des corrélations entre différents paramètres des toiles construites au
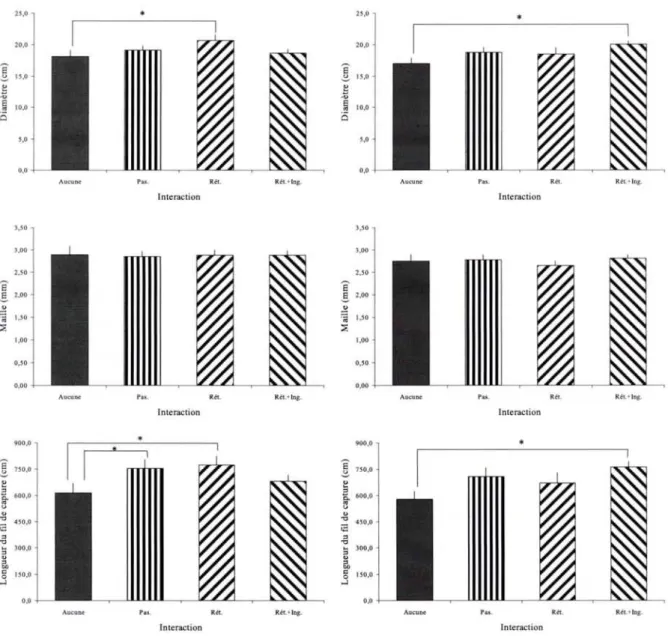
laboratoire par les femelles adultes de Zygiella x-notata (Tab. 2). Par conséquent, les
paramètres retenus pour les comparaisons des différents groupes expérimentaux sont le diamètre et la taille de la maille respectivement considérés comme de bons indicateurs de la taille et de la structure de la toile, ainsi que la longueur du fil de capture qui reflète l'investissement de l'Araignée dans le fourragement.
à une latence par rapport à un moment de référence (en l'occurrence la mise en cadre des Araignées testées) mais au moment réel de la journée (heure solaire) où l'Araignée débute la construction de la toile.
1.3.3. Exploitation statistique des données
Pour les différentes expériences réalisées, différents groupes expérimentaux ont été constitués. Les comparaisons statistiques entre ces groupes des divers paramètres analysés sont réalisées à l'aide de tests paramétriques lorsque nos données remplissent leurs conditions d'application ou à l'aide de tests non paramétriques le cas échéant (Sokal & Rohlf 1995) :
- comparaison de distribution des groupes: Test du X2, Test exact de Fisher ou Test de Mc
Nemar
- comparaison de groupes indépendants: ANOVA ou MANDVA (Analyse de la variance à un ou multiples facteurs), Test t de Student, Test PLSD de Fisher, Test de Kruskal-Wallis ou Test U de Mann-Whitney
- comparaison de groupes appariés: Test de rang de Wilcoxon - corrélation et régression: Test Z et Régression simple
Les protocoles et méthodes d'analyse statistique plus particuliers sont développés ultérieurement dans les chapitres correspondants.
Chapitre
II
Interactions avec les proies et les congénères dans
les conditions naturelles
Chez les Araignées à toile, celle-ci est le moyen de capture des proies; elle fournit ainsi l'énergie pour la survie et la reproduction. C'est aussi le lieu de rencontre des partenaires sexuels; les mâles adultes partent à la recherche des femelles, par la suite, parade et copulation se déroulent généralement au niveau de la toile. La vie et la reproduction des Araignées dépendent directement de leur toile. Elle doit intercepter suffisamment de proies pour que l'Araignée puisse subvenir à ses besoins. Dans cette optique, l'Araignée devrait adapter son comportement de construction aux conditions environnementales, ce qui implique qu'elle devrait prendre en compte les informations qu'elle perçoit de son environnement.
Dans le milieu naturel, les Araignées vivent en présence de prOIes potentielles, de partenaires sexuels, de compétiteurs conspécifiques et hétérospécifiques pour les ressources (nourriture, site d'accueiL), de prédateurs et de parasites. Les relations entre l'Araignée et son environnement se font très souvent par l'intermédiaire de sa toile. Elle constitue effectivement une extension de l'espace sensoriel de l'Araignée: sa structure, et ses caractéristiques physiques et biochimiques assurent la transmission d'informations aux récepteurs sensoriels de l'Araignée (Witt 1975). Les Araignées devraient donc tenir compte des événements se produisant au niveau de leur toile, des informations qu'elles perçoivent via la toile de leur environnement, et moduler leur comportement afin de l'ajuster aux conditions environnementales.
Nous recherchons dans un premier temps s'il existe des relations entre les événements vécus par l'Araignée avec une toile et la construction de la toile suivante. De telles relations pourraient indiquer que l'Araignée a perçu des informations sur son environnement via sa toile, et en a tenu compte pour mettre en place le piège suivant. Nous abordons ce problème
par l'étude de populations naturelles de Zygiella x-notata. Puisque nous avons mis en
évidence que les Araignées de cette espèce peuvent vivre en agrégation et entrent en
compétition pour les ressources du milieu (Leborgne& Pasquet 1987a), nous nous intéressons
aux interactions existant au cours de la journée entre les congénères, et entre les Araignées et leurs proies potentielles, ainsi qu'aux conséquences éventuelles de ces interactions sur le comportement de construction des Araignées.
Figure 5 : Photographie du site d'étude du "Gymnase"
Tl,Tl, T3 : Travées de vitres.
R: Point de référence (xo' Yo) pour la position spatiale des retraites des Araignées. Flèches grisées: Montants des vitres, supports d'installation pour les Araignées.
II.l. Protocoles d'étude
Deux populations naturelles de Zygiella x-notata ont été étudiées. Les Araignées de ces
populations sont alignées le long des vitres d'un gymnase pour la première population, et d'un bâtiment universitaire pour la seconde. La première étude vise à récolter des informations générales sur la construction de la toile, et sur les interactions de l'Araignée avec ses proies et ses congénères dans les conditions naturelles. Le but de la seconde étude est d'essayer de quantifier plus précisément ces interactions et leurs influences mutuelles sur le comportement de construction de l'Araignée.
Une cartographie des deux sites d'étude a été effectuée afin de pouvoir évaluer les densités respectives d'Araignées et de proies; des études antérieures ayant montré que la construction de la toile pouvait dépendre de ces densités (Leborgne& Pasquet 1987a ; Ridwan 1993).
II.l.l. Etude de la population naturelle "Gymnase"
L'étude a été réalisée sur une population colonisant les baies vitrées d'un gymnase. Trois travées de vitres se succèdent le long du bâtiment, d'une longueur d'environ 65 m, séparées les unes des autres par des pans de mur (Fig. 5). Les montants verticaux et horizontaux, en bois, des vitres constituent les supports potentiels où les Araignées peuvent construire leur retraite, et sont, de même que les vitres, les sites d'attachement des toiles. Les deux travées latérales (Tl et T3) présentent les mêmes structures d'accueil; il en est de même pour les deux vitres latérales de chacune de ces travées. La longueur totale des structures d'accueil disponibles le long du bâtiment est de 89,9 m.
Cette étude a eu lieu en Septembre-Octobre 1999, et coïncide en partie (Septembre) avec la période de reproduction de l'espèce. La population est donc essentiellement constituée de femelles adultes et de mâles.
Avant le début des observations, une cartographie de l'ensemble de l'habitat a été réalisée afin de repérer les sites occupés par les Araignées le long des vitres. L'occupation d'un site par une Araignée est matérialisée par la présence d'une retraite renfermant une Araignée. La position spatiale de chaque retraite, dans le plan de la vitre, est relevée (±l cm) par des coordonnées x et y correspondant respectivement à la longueur et la hauteur par rapport à un
Tableau 3 : Position de l'Araignée par rapport au système retraite-toile
Position de l'Araignée
Re Retraite
Re L'Araignée est entièrement à l'intérieur de la retraite
Re1/2p L'Araignée est à l'intérieur de la retraite, une patte, dont seuls les articles terminaux sont visibles, posée sur la fil avertisseur
Rep L'Araignée est à l'intérieur de la retraite, une patte étendue entièrement visible posée sur le fil avertisseur
Rec L'Araignée toujours en contact avec le fil avertisseur est positionnée plus en avant dans la retraite, son céphalothorax étant visible
Rea L'Araignée se situe à l'entrée de la retraite et son abdomen est visible
Fa Fil avertisseur: l'Araignée est totalement sortie de la retraite et se tient sur le fil la reliant au moyeu
Mo Moyeu: l'Araignée est positionnée sur la zone de soie sèche centrale
Sc Spirale de capture: l'Araignée se situe sur l'aire occupée par la spirale gluante Cd Cadre : l'Araignée est sur les fils de cadre de la toile
Su Support: l'Araignée est sur le support où elle a ancré sa toile, reliée à celle-ci par le fil de cheminement
La position plus précise de l'Araignée au sein de sa retraite n'a été relevée que pour les observations diurnes des cinquième et sixième journées; pour les premières journées nous nous sommes contentés de noter qu'elle était dans sa retraite.
point de référence (xo, Yo) situé en haut à gauche de la première vitre de la première travée (Fig. 5). Les Araignées repérées sont marquées comme décrit en 1.3.1., avec un code couleur différent de leurs proches voisines. La cartographie et le marquage sont réactualisés hebdomadairement pour prendre en compte les changements de sites et les arrivées de nouvelles Araignées dans la zone d'étude.
Six journées d'observation ont été effectuées: 8, 10, 15 et 21 Septembre, 8 et 27 Octobre. Au début de chaque journée, les sites occupés par les femelles adultes sont relevés, de même que la présence ou non sur ces sites d'une toile, ainsi que les caractéristiques de cette toile (cf. 1.3.2. pour un rappel des mesures prises sur les toiles). Parmi les toiles présentes, nous distinguons les nouvelles toiles construites au cours de la nuit écoulée des anciennes toiles datant de plus d'une journée; toute toile ne présentant aucun dommage (ni spires captrices accolées ou détruites, ni rayons rompus) est considérée comme étant une nouvelle toile. Les observations des femelles adultes possédant une toile commencent alors. Des observations diurnes sont effectuées de 8hOO à 19h00-20h00 (tombée de la nuit) : deux observateurs font des passages réguliers devant chaque site, l'un effectuant des séries d'observation (scans) d'une minute devant chaque site (il lui faut environ une heure pour tous les visiter, environ 60 sites suivis à chaque journée), le second fait des passages plus rapides lui permettant deux passages par heure devant chaque site. Des observations nocturnes sont également effectuées de OhOO à 8hOO-8h30 (fin de la période de construction) pour les quatre premières journées, et de 5hOO à 8hOO pour la cinquième journée (pas de données nocturnes pour la sixième journée) : un observateur réalise des scans d'une minute devant chaque site, soit un passage par heure, à l'aide d'une lampe de poche munie d'un filtre rouge.
Au cours de ces périodes d'observation diurnes et nocturnes, nous relevons, à chaque passage devant un site occupé, la position de l'Araignée par rapport au système retraite-toile (Tab. 3) et son activité générale (Tab. 4). Nous notons également pour toute interaction entre Araignée et proies, le moment et la nature de l'interaction, la taille de la proie impliquée, la section de la toile où se situe l'interaction, ainsi que la réponse immédiate de l'Araignée à cette interaction. De même pour toute interaction entre Araignées, nous relevons le moment et la nature de l'interaction, le statut résident (propriétaire de la toile, de la retraite) ou intrus des Araignées impliquées, et la réponse immédiate de chacune d'elles. La température et le taux d'humidité régnant sur la zone d'étude sont relevés toutes les heures au point de référence (xo, Yo).
A l'issue de la période d'observation nocturne, un état des lieux de la population est réalisé. Nous relevons les sites occupés par les femelles adultes et la présence ou non sur ces sites d'une toile, nouvelle ou ancienne. Nous mesurons aussi les caractéristiques des nouvelles toiles.
Tableau 4 : Activité générale de l'Araignée
Activité de l'Araignée
lm Immobilité: aucune activité observée Dp Déplacement de l'Araignée
Dét Détection d'une proie: sortie de la retraite et généralement déplacement jusqu'au moyeu, tiraillement des fils de soie (fil avertisseur ou rayons), orientation en direction de la proie
Ca Capture d'une proie: maîtrise et morsure de la proie qui est généralement ramenée à la retraite, mais parfois laissée sur la toile
Ig Ingestion d'une proie: consommation de la proie en général au niveau de la retraite DO Destruction de la toile existante
Co Construction d'une nouvelle toile
La cartographie des sites occupés par les Araignées permet de calculer les distances inter-retraites (une distance inter-retraites étant la distance séparant les retraites respectives de deux Araignées), et sur cette base de déterminer pour chaque Araignée son plus proche voisin, ceci pour chacune des journées d'observation. Ces distances au plus proche voisin sont utilisées par différents auteurs pour rendre compte du mode de distribution des individus d'une population dans un biotope donné (Campbell & Clarke 1971 ; Schoener & Toft 1983 ; Pasquet 1984; Leborgne & Pasquet 1987a). Afin de déterminer le type de répartition (régulier, au hasard ou agrégatif) des Araignées de la population naturelle étudiée, nous utilisons la méthode du "goodness of fit test" (Campbell & Clarke 1971). Cette méthode consiste à comparer la distribution observée des distances au plus proche voisin des Araignées de la population étudiée à une distribution théorique obtenue pour une population de même
densité dont les Araignées sont réparties au hasard (Crisp 1961) par un test du X2(Campbell&
Clarke 1971). Pour cela, nous considérons que les Araignées sont distribuées de façon linéaire le long des vitres. Notons que la méthode présentée ci-dessus nécessite de déterminer la densité de la population étudiée. Pour ce calcul, bien que les montants ne soient pas forcément tous occupés le jour des observations, ils l'ont tous été sur l'ensemble de la saison comme le prouve les anciennes retraites toujours présentes; ils constituent donc tous des supports potentiels qui doivent être pris en compte pour calculer la densité d'Araignées de la population.
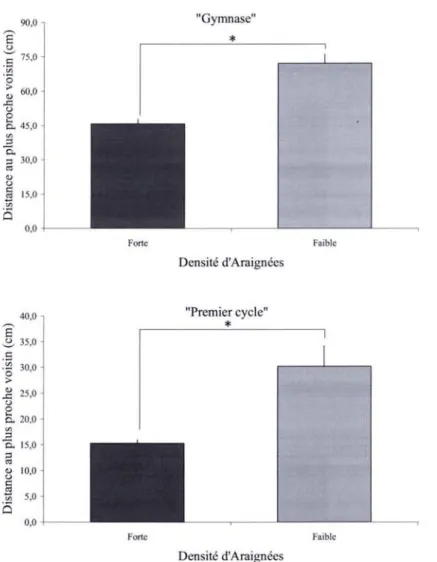
La densité d'Araignées ne semble pas constante tout au long de~ supports. A partir de la
densité moyenne calculée sur l'ensemble des supports du bâtiment, nous pouvons déterminer des zones de forte densité d'Araignées correspondant aux vitres dont la densité est supérieure
à la densité moyenne, et des zones de faible densité correspondant aux vitres dont la densité
est inférieure à la densité moyenne.
Sur le même principe nous définissons des zones (vitres) de plus grande et des zones de plus petite disponibilité en proies, en comparant la disponibilité de chaque vitre à la disponibilité moyenne déterminée sur l'ensemble des supports du bâtiment. La disponibilité en proies est évaluée à partir du nombre d'interactions entre Araignées et proies relevées au cours de nos observations.
Des données récoltées, nous avons retiré des informations générales sur la répartition des Araignées dans le milieu, sur la capture des proies et le comportement de construction de la toile, ainsi que des précisions sur la nature des interactions entre les Araignées et leurs proies. Mais, très peu de données sur les interactions entre congénères ont été récoltées :
seulement six interactions observées se limitantàdes contacts via la soie, n'autorisant pas une
analyse correcte. Deux raisons principales à cette lacune. Premièrement, la difficulté de repérage et d'observation des Araignées au moyen d'un mince faisceau de lumière rouge tamisée dans un lieu plongé dans l'obscurité totale; deuxièmement, la méthode retenue
d'effectuer des passages rapides devant chaque site, résultant du grand nombre d'Araignées à
suivre et par ailleurs efficace pour obtenir un grand nombre d'informations sur la capture des proies, mais vraisemblablement peu adaptée à l'observation d'interactions entre congénères pouvant être très furtives.
Figure 6 : Photographie du site d'étude du "Premier cycle"
Pl ... Pl2 : Panneaux de la zone d'étude du site.
R : Point de référence (xo, Yo) pour la position spatiale des retraites des Araignées. Flèches grisées: Montants des panneaux, supports d'installation pour les Araignées.
II.l.2. Etude de la population naturelle "Premier cycle"
L'étude a été réalisée sur une population occupant le pourtour de panneaux situés sous les fenêtres d'une aile d'un bâtiment universitaire. Sur toute la longueur de l'aile, soit environ 54 m, se succèdent une trentaine de panneaux offrant les mêmes structures d'accueil, et séparés les uns des autres par des poteaux. Cependant la zone d'étude est restreinte aux douze premiers panneaux à compter de l'extrémité de l'aile; les Araignées étant absentes de la quasi-totalité des autres panneaux, et le douzième marquant la limite d'un "no spider's land". Les montants verticaux et horizontaux, métalliques, des panneaux constituent les supports potentiels où les Araignées peuvent construire leur retraite et fixer leur toile (Fig. 6). Le fond des panneaux est constitué d'une plaque de métal argenté. La longueur totale des supports potentiels de la zone d'étude est de 40,2 m.
Cette étude a eu lieu en Septembre-Octobre 2000. La population est alors constituée essentiellement de femelles adultes.
Comme pour l'étude précédente (cf. II. 1.1.), une cartographie de l'habitat à été réalisée afm de repérer les sites occupés par les Araignées, et permettre de déterminer les distances au plus proche voisin et d'analyser la répartition des Araignées le long des supports. Le point de référence (xo, Yo) étant situé en haut à droite du premier panneau (Fig. 6).
Trois journées d'observation ont été effectuées: 12 et 27 Septembre, 17 Octobre. Au début de chaque journée, les sites occupés par les femelles adultes sont repérés. A partir de là, six panneaux présentant des densités en Araignées apparemment différentes sont retenus pour les observations. La présence ou non de nouvelles ou d'anciennes toiles est relevée pour les sites occupés des panneaux retenus, ainsi que les caractéristiques des toiles présentes.
Les dimensions des panneaux sont telles qu'elles permettent à l'observateur de se positionner de façon à avoir l'ensemble des structures d'accueil et donc des Araignées d'un panneau dans son champ de vision, tout en étant à une distance permettant les observations (1,5 m), sans interférer avec les flux de proies potentielles. Des observations diurnes (de 12h00 à 19h00) et nocturnes (de 23h00 à 8hOO) sont effectuées sous forme de périodes de dix minutes passées successivement devant chacun des six panneaux retenus, soit en une heure une période d'observation de dix minutes pour chaque panneau. Pendant la nuit, des passages sont aussi effectués régulièrement devant chaque site occupé, afin de relever les paramètres temporels
Tableau S: Distributions observées et théoriques des distances au plus proche VOISIn
déterminées pour chaque journée d'observation des deux populations étudiées Gymnase
1° jour 2° jour 3° jour 4° jour Sejour 6° jour
N 59 N 64 N 66 N 67 N 67 N 63 P 0,7 P 0,7 P 0,7 P 0,7 P 0,7 P 0,7 Classes (cm) übs Th übs Th übs Th übs Th übs Th übs Th 0-18 8 6,6 10 7,7 10 8,2 12 8,4 6 8,4 12 7,5 18-36 12 5,8 15 6,8 20 7,1 18 7,4 22 7,4 15 6,6 36-54 Il 5,2 11 6,0 7 6,3 Il 6,4 12 6,4 16 5,8 54-72 10 4,6 12 5,2 Il 5,5 10 5,6 9 5,6 8 5,1 >72 18 36,8 16 38,3 18 38,9 16 39,2 18 39,2 12 38,0 df 4 4 4 4 4 4 X2 29,20 36,66 40,25 37,27 48,12 50,88 P <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 Premier cycle
1° jour 2° jour 3° jour
N 92 N 81 N 80 p 2,3 P 2,0 P 2,0 Classes (cm) übs Th übs Th übs Th 0-7 7 13,6 9 10,7 10 10,4 7-14 45 Il,6 37 9,2 24 9,0 14-21 22 9,9 17 8,0 25 7,9 21-28 5 8,4 7 7,0 12 6,8 28-35 5 7,2 1 6,1 1 6,0 35-42 0 6,1 4 5,3 1 5,2 >42 8 35,2 6 34,7 7 34,7 df 6 6 6 X2 143,38 121,76 95,44 p <0.001 <0.001 <0.001
La comparaison des distributions observées (Obs) des distances au plus proche voisin aux distributions théoriques (Th) obtenues pour des populations de mêmes densités (p) dont les individus sont répartis au hasard est réalisée par la méthode du "goodness of fit test".
Tableau 5: Distributions observées et théoriques des distances au plus proche VOlsm déterminées pour chaque journée d'observation des deux populations étudiées
Gymnase
1° jour 2° jour 3° jour 4° jour 5° jour 6° jour
N 59 N 64 N 66 N 67 N 67 N 63
P 0,7 P 0,7 P 0,7 P 0,7 P 0,7 P 0,7
Classes (cm) Obs Th Obs Th Obs Th Obs Th Obs Th Obs Th
0-18 8 6,6 la 7,7 10 8,2 12 8,4 6 8,4 12 7,5 18-36 12 5,8 15 6,8 20 7,1 18 7,4 22 7,4 15 6,6 36-54 Il 5,2 11 6,0 7 6,3 11 6,4 12 6,4 16 5,8 54-72 la 4,6 12 5,2 Il 5,5 10 5,6 9 5,6 8 5,1 >72 18 36,8 16 38,3 18 38,9 16 39,2 18 39,2 12 38,0 df 4 4 4 4 4 4 X2 29,20 36,66 40,25 37,27 48,12 50,88 P <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 Premier cycle
1° jour 2° jour 3° jour
N 92 N 81 N 80
p 2,3 P 2,0 P 2,0
Classes (cm) Obs Th Obs Th Obs Th
0-7 7 13,6 9 10,7 la 10,4 7-14 45 Il,6 37 9,2 24 9,0 14-21 22 9,9 17 8,0 25 7,9 21-28 5 8,4 7 7,0 12 6,8 28-35 5 7,2 1 6,1 1 6,0 35-42 a 6,1 4 5,3 1 5,2 >42 8 35,2 6 34,7 7 34,7 df 6 6 6 X2 143,38 121,76 95,44 p <0.001 <0.001 <0.001
La comparaison des distributions observées (Obs) des distances au plus proche voisin aux distributions théoriques (Th) obtenues pour des populations de mêmes densités (p) dont les
liés à la construction de la toile (début et durée de construction). Les observations nocturnes
sont facilitées par la présence, à une quinzaine de mètres du bâtiment, de deux lampadaires
qui assurent un éclairage diffus tout au long de la nuit permettant les observations générales sur la position des Araignées; une lampe de poche munie d'un filtre rouge étant utilisée pour les observations plus précises des interactions entre congénères ou de l'état de la toile et de l'avancement de sa reconstruction.
Tous les paramètres mesurés au cours de ces observations et à l'issue de la période nocturne ont été décrits pour l'étude de terrain précédente (cf. II.!.!.).
11.2. Structuration des populations naturelles
La méthode du "goodness of fit test" permet la comparaison par un Test du X2 de la
distribution observée des distances au plus proche voisin de la population étudiée à une distribution théorique obtenue pour une population de même densité dont les individus sont
répartis au hasard. Comme pour tout Test du X2, la fréquence théorique de chaque classe de
distance doit être supérieure ou égale àcinq. La taille de classe retenue pour notre analyse est
donc la plus petite distance au plus proche voisin permettant la formation du plus grand nombre de classes dont la fréquence théorique est supérieure ou égale à cinq.
Pour chaque journée d'observation de chacun des biotopes, la distribution observée des distances au plus proche voisin diffère significativement de la distribution théorique (Tab. 5).
Les fréquences observées significativement supérieures (Tests du X2 :P<O.05) aux fréquences
théoriques pour les distances au plus proche voisin de 18 à 54-72 cm pour le "Gymnase" et de
7 à21 cm pour le "Premier cycle", et inversement significativement inférieures (Tests du X2 :
P<O.05) aux fréquences théoriques pour les distances respectivement >72 cm et >28 cm,
montrent que les Araignées des deux populations naturelles étudiées ne sont pas réparties au hasard le long des supports mais présentent une répartition de type agrégatif. Néanmoins, pour la population du "Premier cycle", le fait que la fréquence observée pour des distances au
plus proche voisin de
°
à7 cm soit plus petite que la fréquence théorique semble indiquer, aumoins pour cette population, le maintien d'une distance minimale entre les Araignées.
Cette structure de population semble se maintenir au cours du temps, puisque l'on retrouve la
répartition agrégative à chaque journée d'observation; de plus la distance moyenne au plus
proche voisin ne diffère pas entre les différentes journées (ANOVA, "Gymnase" :F5,380=O,69 ;