B B..PP..88881155KKIINNSSHHAASSAA11 F FAACCUULLTTEEDDEESSSSCCIIEENNCCEESSAAGGRROONNOOMMIIQQUUEESS D DEEPPAARRTTEEMMEENNTTDDEEPPHHYYTTOOTTEECCHHNNIIEE THESE DE DOCTORAT N° . Domaine : Sciences Agronomiques
Spécialité : Sélection et amélioration des plantes Présentée et soutenue publiquement
Par
Jean-Pierre KABONGO TSHIABUKOLE, Ir. Diplômé d’Etudes Approfondies,
Le 07 Novembre 2018
EVALUATION DE LA SENSIBILITE AUX STRESS HYDRIQUES DU
MAÏS (Zea mays L.) CULTIVE DANS LA SAVANE DU SUD-OUEST
DE LA RD CONGO, CAS DE MVUAZI
Promoteur : Pr. MUMBA DJAMBA (UPN)Co Promoteurs : Pr. KABWE NKONGOLO (Université Laurentienne d’ONTARIO / CANADA),
Pr. KIZUNGU VUMILIA (UNIKIN)
JURY Président : Pr. LOMA TONGOMO (UPN)
Secrétaire : Pr. NYONGOMBE UTSHUDIENYEMA (UPN) Membres : Pr. MONZAMBE MAPUNZU (UPN)
Pr. KALAMBAIE BINM MUKANYA (UPN)
TABLE DES MATIERES INTRODUCTION GENERALE 7 1. Concepts 7 3. Hypothèses 10 4. Objectif général 11 5. Objectifs spécifiques 11
6. Organisation et subdivision de la thèse 11
CHAPITRE 1 14
SYNTHESE BIBLIOGRAPHIQUE 14
1.1. Le maïs tropical 14
1.1.1. Description botanique 14
1.1.2. Physiologie 14
1.1.2.1. Capture et utilisation du carbone 15
1.1.2.2. Répartition des matières assimilées 17
1.1.2.3. Avortement à la floraison 18 1.1.2.4. Remplissage du grain 19 1 .1.2.5. Dessiccation du grain 20 1.1.3. Production 21 1.1.4. Culture et fertilisation 21 1.1.4.1. Culture de maïs 21 1.1.4.2. Fertilisation de maïs 22 1.1.4.3. Besoins en eau 24
1.1.5. Importance dans l’alimentation 26
1.1.6. Place du maïs en RDC 28
1.1.7. Notion de la sécheresse en agriculture 29
1.1.8. Contraintes de la culture du maïs en Afrique subsaharienne 30
1.2. Les stress abiotiques affectant la culture de maïs 32
1.2.1. Stress hydrique 32
1.2.1.1. Symptômes de stress hydrique 32
1.2.1.1.1. Avant floraison 32
1.2.1.1.2. A la floraison 32
1.2.1.1.3. Après floraison 33
1.2.1.2. Situation à risque 33
1.2.2. Stress azoté et phosphoré 35
1.2.3. Stress thermique 36
1.2.4. Stress salin 38
1.2.5. Similitude entre les stress abiotiques 39
1.3. Mécanismes d’adaptation des plantes aux stress abiotiques 39
1.3.1. Esquive ou échappement 40
1.3.2. Evitement de la déshydratation des tissus 41
1.3.3. Résistance à la déshydratation des tissus 42
1.4. Les indices de tolérance aux stress hydriques 44
1.5.1. Simulation des cultures 46 1.5.2. Simulation de la surface foliaire et de l’indice de surface foliaire 46
1.5.3. Simulation de l’indice de surface foliaire 47
1.6. Milieu d’étude 48
1.6.1. Centre de recherche de Mvuazi 48
1.6.2. Climat de Mvuazi 48
1.6.3. Sol de Mvuazi 50
1.6.4. Végétation de Mvuazi 51
CHAPITRE 2 52
MESURE DES PARAMETRES DE TOLERANCE A LA SECHERESSE 52
2.1. Introduction 52
2.2. Matériels et méthodes 53
2.3. Analyses statistiques 54
2.4. Résultats et discussion 55
2.4.1. Analyse de la variance de paramètres étudiés 55
2.4.2. Régression linéaire 56
2.4.3. Corrélation entre les variables 57
2.4.4. Analyse en composantes principales (ACP) 58
2.5. Conclusion 61
CHAPITRE 3 62
LES INDICES DE TOLERANCE AUX STRESS HYDRIQUES DU MAÏS (Zea mays .L) CULTIVE
DANS LE SUD OUEST DE LA REPUBLIQUE DEMOCRATIQUE DU CONGO 62
3.1. Introduction 62
3.2. Matériels et méthode 63
3.2.1. Calcul des indices de stress 64
3.2.2. Analyses statistiques 65
3.3. Résultats et discussion 65
3.3.1. Relation entre Ri et Rs avec l’indice de tolérance aux stress (STI) 67
4.3.2. Relation entre Ri et Rs avec l’indice MP 68
3.3.3. Relation entre Ri et Rs avec l’indice MGP 69
3.3.4. Relation entre Ri et Rs avec l’indice SSI 70
3.3.5. Relation entre Ri et Rs avec l’indice ISR 72
3.3.6. Relation entre Ri et Rs avec l’indice TOL 72
3.3.7. Analyse en composantes principales (ACP) 74
3.4. Conclusion 75
CHAPITRE 4 77
ANALYSE DE LA REPONSE ADAPTATIVE DES VARIETES DE MAÏS (Zea mays L.) DE LA RD
CONGO AUX DEFICITS HYDRIQUES 77
3.2. Matériels et méthode 77
4.3. Analyses statistiques 80
4.4. Résultats et discussion 80
4.4.1. Hauteur de plant (HP) 80
4.4.2. Nombre de feuilles vertes (NFV) 81
4.4.3. Indice de surface foliaire (LAI) 82
4.4.4. Volume de système racinaire (VR) 84
4.4.5. Rendement en grain (g/m2) 85
4.4.5. Biomasse totale (BM) 86
4.5. Conclusion 88
CHAPITRE 5 89
SIMULATION DE LA CROISSANCE ET DE L’INDICE DE SURFACE FOLIAIRE DU MAÏS A QUALITE PROTEIQUE DANS LES CONDITIONS SAISONNIERES DE LA SAVANE DU SUD
OUEST DE LA RDC 89
5.1. Introduction 89
5.2. Matériels et méthode 90
5.2.1. Mesure des paramètres 91
5.2.2. Calcul du temps thermique 93
5.3. Résultats et discussion 95
5.3.1. Simulation de la croissance de maïs 95
5.3.1.1. La hauteur des plants 95
5.3.1.2. Nombre des feuilles 96
5.3.2. Simulation de l’Indice de surface foliaire 98
5.4. Conclusion 100
CHAPITRE 6 101
DISCUSSION GENERALE, CONCLUSION GENERALE ET PERSPECTIVES 101
6.1. Discussion générale 101
6.1.1. Mesure des paramètres de tolérance à la sécheresse 101
6.1.2. Les indices de tolérance aux stress 102
6.1.3. Analyse de la réponse adaptative de maïs au déficit hydrique 102
6.1.4. Simulation de la croissance et de l’indice foliaire de QPM 104
6.2. Conclusion générale et perspectives 105
REFERENCES BIBLIOGRAPHIQUES 109
LISTE DES TABLEAUX
Tableau 1.1.Quelques facteurs environnementaux affectant l’EC chez le maïs tropical de basses altitudes 16
Tableau 1.2. Les besoins en minéraux du maïs pour boucler le cycle 22
Tableau 1.3. Absorption des éléments majeurs au cours d’une saison 23
Tableau 1.4. Les 15 premiers producteurs de maïs en Afrique entre 2000 et 2014 27
Tableau 1.5. Evolution de la production de maïs par province (en tonnes) 28
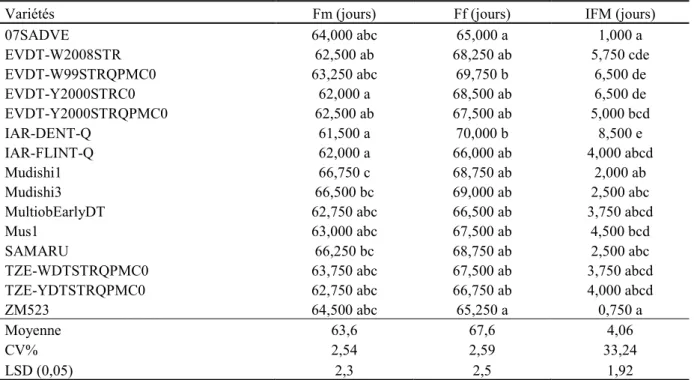
Tableau 2.1. Analyse comparée des variétés de maïs basée sur la floraison mâle et femelle (Fm et Ff) et
l’intervalle entre floraison (IFM) 55
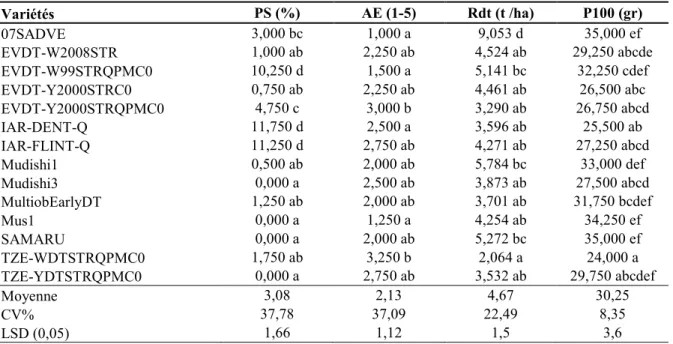
Tableau 2.2. Analyse comparée des variétés de maïs basée sur le % de plants stériles (PS), aspect des épis (AE),
rendement en grain (Rdt), et poids de 100 grains (P100) 56
Tableau2.3. Matrice de corrélation entre pairs des variables 57
Tableau 3.1. Origines et types de variétés de maïs testées pour les indices de tolérance aux stress 64
durant la saison de 2013 et de 2014 à Mvuazi 64
Tableau 5.1. Estimation des paramètres du modèle pour deux variétés de maïs au cours de deux saisons pour la
hauteur des plants. 95
Tableau5.2. Estimation des paramètres du modèle pour deux variétés de maïs au cours de deux saisons pour le
nombre de feuilles visibles. 97
LISTE DES FIGURES
Figure 0.1. Phénotype : Comportement de la plante en réponse à l’environnement 8 Figure 1.1. Remplissage des grains sur épi en fonction du niveau de stress (source : Kabongo et al., 2016) 32 Figure 1.2. Conséquence du déficit hydrique au moment de floraison : avortement des ovules. (Source : Kabongo
et al., 2016) 33
Source (Jean-Marc, 2015) 34
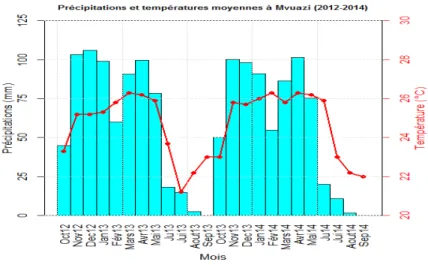
Figure 1.3. Pourcentage de perte de rendement par jour de stress hydrique à différentes époques de la saison 34 Figure 1.4. Evolution du rendement en matière sèche et absorption des éléments fertilisant N-P-K. 36 Figure 2.5. Situation géographique du Centre de Recherche de Mvuazi (Zone d’étude) 48 Figure 3.6. Situation climatique durant la période expérimentale dans le centre de Mvuazi et environs
(2012-2014) 49
Figure 2.1. Relation entre le rendement en grains des variétés et l’intervalle de floraison. 57 Figure 2.3. Biplot (axes F1 et F2 : 80,86 %) dispersion des variétés en fonction des variables étudiées 59 Figure 2.4. Effet du stress hydrique sur l’intervalle de floraison et le remplissage des épis de maïs 59 Figure 3.1. Moyenne des rendements et indice de sensibilité au stress en 2013et 2014 66
Figure 3.2. Relation entre Ri et Rs 67
Figure 3.3(a). Relation entre STI et Rendements Ri, (b). Relation entre STI et Rendements Rs 68 Figure 3.4. (a) Relation entre Rendements Ri et MP, (b). Relation entre Rendements Rs et MP 69 Figure 3.5(a). Relation entre MGP et Rendements Ri. (b). Relation entre MGP et Rendements Rs 70
Figure 3.6. Relation entre SSI et Rendements Ri et Rs 71
Figure 3.7 (a). Relation ISR et rendement Ri, (b) Relation ISR et rendement Rs 72 Figure 3.8(a). Relation entre TOL et Rendements Ri, Relation entre TOL et Rendements Rs 73 Figure 3.9. Répartition des génotypes générés par l'ACP des le rendement et indices stress 74 Figure 4.1. Installation du dispositif expérimental. Suivi et prise des données des paramètres d’étude 78
Figure 4.2. Mesures du volume des racines. 79
Figure 4.3. (a) Evolution de la croissance des tiges par variété en tous régimes hydriques confondus. (b). Evolution de la croissance des tiges par régime hydrique pour toutes les variétés confondues. Allongement de la tige (A), 50% floraison mâle (B), 50% de floraison féminine (C), 50% de formation d'épi (D), remplissage d'épi (desséchement de soie) (E), 50% de maturité de l’épi (F) (G), 50% de sénescence foliaire (H), sénescence totale (feuilles et tiges) (I) et dessiccation des grains (J). (100%WW=100%CR,
60%WW=60%CR, 30%WW=30%CR) 81
Figure 4.4. (a).Evolution du nombre de feuilles vertes visibles par variété en tous régimes hydriques confondus. (b). Evolution du nombre de feuilles vertes visibles par régime pour toutes les variétés confondues. Allongement de la tige (A), 50% floraison mâle (B), 50% de floraison féminine (C), 50% de formation d'épi (D), remplissage d'épi (desséchement de soie) (E), 50% de maturité de l’épi (F) (G), 50% de sénescence foliaire (H), sénescence totale (feuilles et tiges) (I) et dessiccation des grains (J).
(100%WW=100%CR, 60%WW=60%CR, 30%WW=30%CR) 82
Figure 4.5. (a). Evolution de LAI de variété en tous régimes confondus. (b).Evolution de LAI par régime hydrique pour toutes les variétés confondues. Allongement de la tige (A), 50% floraison mâle (B), 50% de floraison féminine (C), 50% de formation d'épi (D), remplissage d'épi (desséchement de soie) (E), 50% de maturité de l’épi (F) (G), 50% de sénescence foliaire (H), sénescence totale (feuilles et tiges) (I) et
dessiccation des grains (J). (100%WW=100%CR, 60%WW=60%CR, 30%WW=30%CR) 83
Figure 4.6. (a).Evolution du volume de racines par variété pour tous les régimes hydriques confondus.
(b).Evolution du volume de racine par régime hydrique pour toutes les variétés confondues. Allongement de la tige (A), 50% floraison mâle (B), 50% de floraison féminine (C), 50% de formation d'épi (D), remplissage d'épi (desséchement de soie) (E), 50% de maturité de l’épi (F) (G), 50% de sénescence foliaire (H), sénescence totale (feuilles et tiges) (I) et dessiccation des grains (J). (100%WW=100%CR,
60%WW=60%CR, 30%WW=30%CR) 84
Figure 4.7. (a) Moyennes de rendement par régimes hydriques pour toutes les variétés confondues. (b)
Figure 4.8. (a) Moyennes de Biomasses par régimes hydriques pour toutes les variétés confondues. (b)
Moyennes de biomasses par variété par régime hydrique. 86
Figure 5.1. Situation sur les moyennes des précipitations et des températures durant l’expérimentation 91 Figure 5.2. Courbe de simulation de la croissance foliaire et paramètres du modèle STICS 94 Figure 5.3. Simulation des stades phénologiques et reproducteurs selon STICS 94 Figure 5.4. Modélisation des cultures : Analyse automatique des données prises aux champs 95 Figure 5.5. Evolution de la hauteur des plants de deux variétés de maïs (Mudishi1 et Mudishi3). 96 Figure 5.6. Evolution du nombre des feuilles visibles de deux variétés de maïs (Mudishi1 et Mudishi3). 97 Figure 5.7. Simulation de l’indice foliaire de deux variétés de maïs par rapport au temps thermique des
INTRODUCTION GENERALE
1. ConceptsLa sécheresse est l'un des facteurs abiotiques les plus préjudiciables à travers le monde. Elle peut gravement entraver la productivité des cultures agricoles. Même dans les zones tropicales, où les pluies fournissent en moyenne 2600 mm d’eau annuellement, la réserve en eau du sol peut diminuer fortement durant la saison sèche et limiter la croissance de la végétation (Cornic et Massacci, 1996).
Le maïs est parmi les cultures céréalières de premier plan dans le monde, mais il est sensible à la sécheresse. Il est affecté par cette dernière à différents stades de croissance dans différentes régions du globe. Le pouvoir germinatif, la levée, l’établissement des peuplements des plantules, la croissance et les développements globaux, le développement pollinique, le développement de la soie, l’intervalle anthèse-soies, la pollinisation, le développement embryonnaire, le développement de l'endosperme et le développement du noyau sont les événements de la vie du maïs qui sont sérieusement entravés par le stress hydrique dû à la sécheresse. Le rendement en grains de maïs est aussi une des variables la plus sévèrement contrainte par la sécheresse qui se produit pendant ou peu avant la floraison ; la période la plus vulnérable pour le maïs (Magorokosho et al., 2003) et les pertes varient entre 21et 50% (Edmeades et al., 1995 ; Ashley, 1999).
Le stress hydrique est un accident relativement fréquent dans les sols à faible réserve hydrique, en culture pluviale ou insuffisamment irriguée. Il affecte plusieurs variables dans la physiologie de la plante, telles que la régulation de la température foliaire (Patel et al., 2001; Luquet et al., 2004), la conductance stomatique et la surface foliaire (Lowlor et Cornic, 2002), ainsi que la photosynthèse (Yuan et al., 2004) avec pour conséquence une limitation biochimique des chloroplastes qui servent à fixer le dioxyde de carbone (Tartieu et Simoneau, 1998). Les processus physiologiques qui affectent la croissance des feuilles sous contrainte de la sécheresse, telles que les propriétés mécaniques de la paroi cellulaire (Cosgrove, 2005; Muller et al., 2007) ou des propriétés hydrauliques de cellules en croissance (Tang et Boyer, 2002 ; Bouchabké et al., 2006) ont été largement étudiés. Le stress hydrique entraîne une modification de l’expression de nombreux gènes (Gaufichon et al., 2010). Cependant, les
plantes ont développé des nom dus à la sécheresse.
Figure 0.1. Phénotype : Comportemen Les germoplasmes de maïs ont de mieux faire face à la séchere plantes à échapper au stress h hydrique soit en réduisant les p qu'une légère réduction du pote la croissance des feuilles de ma l'élongation des cellules de p l'écoulement de l'eau du xylème Westgate et Boyer (1985) ; Tan du maïs s’était arrêtée lorsque auteurs (Sarah et Tardieu, 1997 des feuilles des plants de maïs s bien arrosés.
Afin de décrire la stabilité et l’é l’environnement et aux impacts des conditions de stress et de
nombreuses stratégies qui leur ont permis de fai
rtement de la plante en réponse à l’environnement
ïs ont des nombreuses caractéristiques qui permette écheresse. Une des stratégies adaptatives est la pré ress hydrique. Certains génotypes ont la capacité t les pertes d'eau ou en augmentant l'absorption d u potentiel hydrique du milieu des racines diminu de maïs (Boyer, 1970; Acevedo et al., 1979). Da
de plantes supérieures peut être inhibée par ylème dans les cellules environnantes d’élongation
Tanguilig et al., (1987) ont rapporté que la crois rsque le potentiel hydrique foliaire de base était < , 1997 ; Stone et al., 1998; Cakir, 2004) ont montr maïs soumis à la sécheresse étaient plus petites qu
et l’élasticité des cultivars vis-à-vis de l’expositio pacts du changement climatique, des études ont é et de non-stress (Ben Salem, 1995). Les appr
de faire face aux stress
rmettent aux accessions la précocité qui aide les té d'éviter le stress tion d'eau. Il est connu diminue immédiatement Dans ces conditions, e par l'interruption de gation (Nonami, 1998). a croissance des feuilles était < -10 bar. D'autres montré que les surfaces tes que celles des plants
osition aux éléments de s ont été conduites sous s approches permettant
d’estimer l’impact de la variabilité climatique sur les rendements ont été également étudiées (Nasr et al., 1998), à savoir : le développement de méthodes d’efficience d’utilisation de l’eau (Boussen et al.,2005), l’utilisation des régressions linéaires et des coefficients évaluant le degré de stabilité du rendement (Rezgui et al.,1999).
L’ajustement osmotique par accumulation d'osmolytes, l’autodéfense végétale par accumulation d'antioxydants, les régulateurs de croissance des plantes, les protéines des stress et les protéines des canaux d'eau, la transcription des facteurs et les voies de transduction du signal sont impliqués dans l'attribution de la tolérance à la sécheresse sur le maïs.
2. Problématiques
Le déficit hydrique est le problème majeur qui affecte la production et la qualité des céréales en conditions de savane. Plusieurs stratégies, y compris des changements dans les pratiques agricoles et le choix des cultivars plus tolérants à la sécheresse ont été adoptées pour résoudre ce problème (Ben Salem et al., 1997). Les résultats de plusieurs enquêtes précédentes ont montré que l’interaction du génotype et de l’environnement (G x E) pourrait être décrite en partie par les indices de tolérance aux stress (Maleki et al., 2008).
Des grands efforts ont été fournis par le CIMMYT, l’IITA et d'autres sociétés multinationales dans le développement du maïs tolérant à la sécheresse, tant sur les variétés à pollinisation ouverte (VPO) que sur les hybrides. Les hybrides tolérants à la sécheresse et les VPO sont cultivées dans de nombreux pays africains. Il y a un besoin d’améliorer encore le niveau d'adaptabilité à la sécheresse pour lutter contre le problème mondial de la sécurité alimentaire. La sélection pour la résistance à la sécheresse repose sur l'identification et la création de génotypes à grande capacité d’adaptation capables de produire un maximum de biomasse ou de grains en conditions de déficit hydrique. Plusieurs critères physiologiques et biochimiques devraient être ainsi identifiés dans le but de distinguer les variétés sensibles des variétés résistantes au stress hydrique.
La difficulté d'identifier et de caractériser les paramètres de la résistance au stress hydrique chez les plantes, à travers l'observation d'un caractère phénotypique complexe et de faible héritabilité, comme le rendement en conditions de déficit hydrique, a conduit à s'intéresser à
des critères morpho-physiologiques de la résistance à la sécheresse. Des approches analytiques, consistant à isoler et à étudier individuellement un mécanisme de résistance donné, via l'observation d'un paramètre particulier (critère de sélection) ont été proposées dans ce travail.
Le développement des variétés, par des méthodes classiques, étant longue et onéreuse (Banziger et al., 2006), l’utilisation des techniques de biotechnologie représente une alternative fiable pour la recherche sur la résistance aux stress abiotiques (Lutts et al., 1996). Cependant, il existe encore très peu d’informations sur la réponse adaptative vis-à-vis des conditions hydriques très limitées et les mesures directes pour apprécier le niveau de sensibilité variétale aux stress hydriques pour les variétés de maïs diffusées et cultivées dans la savane du sud ouest de la République Démocratique du Congo en général et dans la contrée de Mvuazi en particulier.
Les connaissances sur les effets du stress hydrique, les réalisations, les stratégies d'adaptation, et les outils de reproduction possibles sont les conditions préalables à tout plan de production. De ce fait, nous avons pris soin de compiler ces aspects dans ce travail.
3. Hypothèses
Suivant les problématiques évoquées ci-haut, nous tenterons de vérifier quelques hypothèses en rapport avec notre thème de recherche, à savoir :
Les génotypes sensibles à la sécheresse auraient un intervalle de floraison plus long que les génotypes tolérant la sécheresse ainsi les lignées à haute teneur en acides aminés seraient plus vulnérables à l’effet de sécheresse ;
L’étude des rapports entre les indices de tolérance aux stress et la différence entre le rendement potentiel et déficitaire, pourrait aider à classer les variétés en fonction de leur degré de tolérance à la sécheresse ;
L'analyse des caractéristiques morphologiques et phénologiques pourrait contribuer à notre compréhension des mécanismes d'adaptation des cultures de maïs au déficit hydrique ;
L’estimation des paramètres du modèle de Baret pour les nouvelles conditions de stress hydriques de savane, pourrait caractériser la croissance et le rendement saisonnier de deux variétés de maïs à protéique de qualité cultivée en RDC.
4. Objectif général
L’objectif général de ce travail est de déterminer la capacité d’adaptation variétale de différents génotypes de maïs aux conditions hydriques déficitaires et de proposer des itinéraires techniques susceptibles d’assurer la stabilité des rendements afin de contribuer à l’amélioration de la sécurité alimentaire et nutritionnelle de Mvuazi et ses environs dans le sud-ouest de la RDC.
5. Objectifs spécifiques
Les objectifs spécifiques de ce travail ont été choisis en fonction du sous thème à développer. Les deux premiers objectifs spécifiques 1 et 2 ont pour fondement le phénotypage et les deux autres 3 et 4 sont orientés vers la modélisation des cultures.
1. Renforcer les connaissances sur l'utilisation des mesures des paramètres phénotypiques dans la sélection des génotypes résistant à la sécheresse ;
2. Déterminer les indices de sélection et d’amélioration du rendement de maïs face aux conditions de déficit hydrique ;
3. Analyser les effets de l’impact du stress hydrique sur les variables phénotypiques des quelques variétés de maïs afin d’apprécier leur réponse adaptative dans des environnements d'eau contrastés ;
4. Développer et évaluer les approches logistiques et/ou exponentielles simulant le développement et l’indice de surface foliaire du maïs à qualité protéique cultivé dans la savane du sud ouest de la RDC.
6. Organisation et subdivision de la thèse
Hormis l’introduction générale, la conclusion générale et suggestions, le 1er chapitre traite la synthèse bibliographique et le milieu d’étude. Le 2e chapitre présente les mesures des
paramètres de tolérance à la sécheresse. Le 3e chapitre présente les indices de tolérance aux stress hydriques du maïs. Le 4e chapitre présente l’analyse de la réponse adaptative de maïs aux stress hydriques. Le 5e chapitre présente la simulation de la croissance et de l’indice de surface foliaire du maïs à qualité protéique dans les conditions saisonnières de la savane du sud ouest de la RDC et le 6e chapitre présente la discussion générale et la conclusion générale.
CHAPITRE 1
SYNTHESE BIBLIOGRAPHIQUE
1.1. Le maïs tropical1.1.1. Description botanique
Le maïs (Zea mays L.) est une céréale herbacée annuelle, à tallage faible à nul, du genre Zea, appartenant à la famille des Poacées (Graminées) tribu des Maydes. C’est la seule espèce cultivée de ce genre et la seule de grande importance économique de la tribu (Reeves et Mangelsdorf, 1942). Comme la plupart des Poacées tropicales, le maïs présente un métabolisme photosynthétique de type C4 (le premier glucide formé comportant quatre atomes de carbone), qui confère à la plante une efficience supérieure à celle des Poacées tempérées dans la conversion de l'énergie lumineuse (Gallais, 1984 ; Gay, 1984). C'est une plante de jours courts, dont les variétés tropicales sont souvent photopériodiques. Ce caractère oligogénique a pu être éliminé lors de l'adaptation de l'espèce aux latitudes tempérées.
Le maïs est une plante monoïque. Il porte deux types d’inflorescence : les fleurs mâles, groupées sur une panicule terminale ramifiée, et les fleurs femelles, associées sur un ou quelques épis insérés aux aisselles des feuilles. Bien que le maïs soit auto fertile, l'allogamie est prépondérante et atteint 95%. Elle résulte de la monoécie et de la protandrie de la plante. La forte allogamie du maïs entraîne la présence d'un lourd fardeau génétique dans les populations non sélectionnées, à l'origine d'une importante dépression de consanguinité et d'une forte vigueur hybride corrélative. Les hybrides issus des lignées homozygotes sont de 200 à 300 % supérieurs à leurs parents (Hallauer et Miranda, 1981).
1.1.2. Physiologie
La physiologie des cultures en général et du maïs en particulier a été décrite dans de nombreuses études détaillées (Fischer et al., 1987 ; Pearson et Hall, 1984; Hay et Walker, 1989 ; Squire, 1990 ; Evans, 1993).
Les aspects de la physiologie du maïs qui peuvent être manipulés pour l’amélioration de sa production et pour la sélection des variétés comprennent les points suivants :
1.1.2.1. Capture et utilisation du carbone
La production d’une culture dépend des radiations solaires et de leur conversion en biomasse. La quantité de radiation incidente qui est interceptée par la plante est déterminée par la surface foliaire, l’orientation des feuilles et leur durée de vie. L’indice de surface foliaire du maïs (ISF ou LAI, surface foliaire par unité de surface du sol) a jusqu’à une valeur d’environ 4, une incidence positive sur l’interception de radiations. Au-delà de cette valeur, la surface additionnelle a peu d’effets sur l’interception de la lumière. La densité de plantes est un facteur déterminant de LAI et de l’interception des radiations. Les variétés précoces produisent moins de feuilles pour intercepter les radiations et nécessitent, comparativement aux variétés tardives, une densité de semis plus forte pour un rendement optimum. La quantité totale de radiations interceptée pendant une saison dépend du temps nécessaire pour une interception maximale et de la durée de vie de la surface verte des feuilles. Par exemple, la fraction de la radiation totale interceptée pendant une saison était de 0,46 dans le cas d’un maïs tropical cultivé en sept lieux différents avec une faible fertilisation azotée (Muchow, 1994). Une culture ayant un LAI maximum d’environ 2 intercepte seulement 37 % de radiations reçues durant la saison, et une culture fortement azotée avec un LAI maximum d’environ 4,5 intercepte 58% des radiations. Après la floraison, le taux de sénescence affecte la capture de la lumière ; la sénescence peut être accélérée par les maladies, le stress hydrique, une faible fertilité du sol et des facteurs génétiques.
Il existe une variabilité génétique importante et hautement héréditaire pour l’orientation des feuilles. L’impact de l’architecture du couvert végétal sur l’interception des radiations et son utilisation ont fait l’objet de nombreuses études. Des simulations indiquent que des feuilles supérieures érigées associées à des feuilles basses horizontales correspondent à une utilisation optimum des radiations du couvert végétal. Cet effet est plus important en milieu tropical où l’angle solaire est plus important (Pearson et Hall, 1984). Mais cet effet est aussi plus faible pour la plantes en C4 comme le maïs que pour les plantes en C3 (Hay et Walker, 1989). Chez le maïs, la contribution des spathes à l’assimilation d’éléments nutritifs par l’épi est plus importante que celle des autres feuilles (Edmeades et al., 1979). Les feuilles érigées au dessus de l’épi permettent un meilleur éclairement des spathes, entrainant un bénéfice additionnel de l’architecture érigée des feuilles. L’éclairement des feuilles les plus basses, qui est aussi favorisé par les feuilles érigées de la partie haute de la plante, est important pour l’absorption continue d’éléments nutritifs durant le remplissage des grains. Les deux facteurs : l’espèce
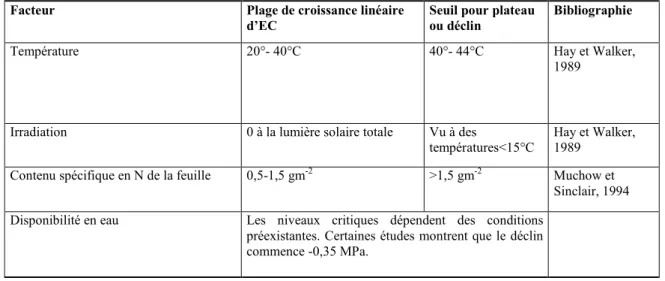
végétale et l’environnement agissent sur l’efficacité d’utilisation des radiations solaires (efficacité de conversion : EC). Le maïs présente un processus de photosynthèse en C4 qui permet une réponse continue à l’augmentation des radiations jusqu’au plein soleil et ce, à de faibles niveaux de photo-respiration. Ces caractéristiques correspondent à une bonne adaptation aux fortes températures et aux fortes intensités d’éclairement rencontrées dans beaucoup de régions tropicales. D’autres facteurs ayant une grande influence sur l’EC sont l’eau et la disponibilité en élément nutritifs (Tableau 1.1). Le taux maximal de photosynthèse chez le maïs tropical se situe entre 30°C et 40°C (Norman et al., 1995). L’effet de la température sur l’EC est relativement faible dans une plage de 20°C à 40°C. Toutefois, les températures en dehors de la plage d’adaptation du cultivar (en deçà de 15°C et au-delà de 44°C pour les maïs tropicaux de basses altitudes) peuvent réduire l’EC.
Tableau 1.1.Quelques facteurs environnementaux affectant l’EC chez le maïs tropical de basses altitudes
Facteur Plage de croissance linéaire
d’EC Seuil pour plateau ou déclin Bibliographie
Température 20°- 40°C 40°- 44°C Hay et Walker,
1989
Irradiation 0 à la lumière solaire totale Vu à des
températures<15°C Hay et Walker, 1989 Contenu spécifique en N de la feuille 0,5-1,5 gm-2 >1,5 gm-2 Muchow et
Sinclair, 1994 Disponibilité en eau Les niveaux critiques dépendent des conditions
préexistantes. Certaines études montrent que le déclin commence -0,35 MPa.
Source : (Lafitte, 2000)
Les radiations d’ondes couvertes journalières varient d’environ 10 MJ m-2 j-1 (radiation solaire incidente totale en mégajoule, dans une plage de longueur d’onde de 0,4 à 3 micromètres par mètre carré par jour) en région nuageuse à 25 MJ m-2 j-1 en région semi-aride et la saison sèche. Comme indiqué précédemment, en l’absence de stress, le maïs semé à forte densité peut intercepter environ 55% des radiations totales reçues pendant la saison. Muchow (1994) a montré que pour des maïs non stressés, les efficacités de conversion varient dans une plage de 1,2 à 1,6 g de biomasse au-dessus du sol par MJ de radiation solaire interceptée. Dans des bonnes conditions d’environnement sans stress, le maïs tempéré peut avoir un taux de
croissance supérieur à 500 kg/ha/j pendant des périodes de plusieurs semaines, entrainant une productivité élevée. En régions tropicales, avec des températures plus élevée et des jours plus courts, le taux de croissance maximum rencontré varie de 250 à 350 kg/ha/j (Agus et al., 1983).
1.1.2.2. Répartition des matières assimilées
Le maïs, comme les autres plantes, tend à maintenir un équilibre fonctionnel entre la masse des racines et des parties aériennes de la plante. Si un des éléments tirés du sol comme l’eau ou un des éléments nutritifs vient à manquer, plus d’éléments nutritifs migreront vers la racine et la croissance racinaire sera favorisée par rapport à la partie aérienne de la plante. Dans le cas où les radiations limitent la croissance (ombre ou conditions nuageuses), plus d’éléments nutritifs seront consacrés à la croissance de la partie aérienne et le rapport racine/partie aérienne décroit.
Le rendement du maïs dépend de la quantité de biomasse qui est affecté au grain. Ceci se reflète dans l’indice de récolte (IR) (IR = la masse grain par rapport à la biomasse de la partie aérienne de la plante). L’IR d’un maïs cultivé dans les conditions favorables varie de 0,2% des variétés locales non améliorées à 0,3-0,4% des variétés améliorées et plus de 0,5% des variétés tempérées précoces. L’IR du matériel tempéré était déjà élevé dans les années 30 (environ 0,45), aussi les améliorations ont-elles plus résulté d’une augmentation de la biomasse que d’une amélioration de l’IR. Les possibilités d’augmenter et de stabiliser l’IR chez les maïs tropicaux sont encore importantes (Hay, 1995). L’indice de récolte est aussi très sensible aux facteurs environnementaux comme la température (pour des raisons inconnues) et aux conditions particulières qui peuvent réduire la quantité de matière assimilée par la plante qui sera transféré vers les organes reproducteurs (environ dix jours avant et après anthèse). A la floraison, l’effet de ces stress est catastrophique (disproportionnée avec la réduction de disponibilité de l’élément). Ils peuvent avoir pour conséquence une forte réduction du nombre de grains et même l’absence totale de grains sur certains épis. La conséquence de stress à d’autres stades de développement, tend à être plus directement proportionnelle à la réduction de l’efficacité de conversion et/ou de l’interception de radiation (Paliwal, 2002).
1.1.2.3. Avortement à la floraison
La température est le premier facteur qui agit sur le développement du maïs. Les cultivars sont classés soit précoces ou tardifs selon la somme de température nécessaire à l’accomplissement de certaines étapes de leur développement. Les besoins en température sont mesurés en somme de température cumulée au-dessus d’un minimum et au dessous d’un optimum nécessaire pour le développement (Isaa, 2013). Ces températures sont des sommes de degré-jours. La floraison est souvent utilisée comme l’événement du développement qui caractérise un cultivar en rapport avec sa précocité. Les cultivars de régions tropicales d’altitudes paraissent avoir des besoins en température pour le développement minimum et/ou optimum, plus bas que les cultivars adaptés aux basses terres tropicales. Le rendement dépend grandement de la quantité de radiation accumulée par la culture par unité de température par unité de temps. Une culture de maïs tropical a généralement un rendement plus bas que le maïs tempéré parce que les températures sous les tropiques sont plus élevées et qu’il accomplit son cycle en un temps plus court. De la même façon, les cultivars précoces ont des rendements généralement moins élevés que les tardifs. Quand le rendement est exprimé sur une base journalière, les rendements des cultivars précoces sont équivalents ou quelque fois supérieurs à ceux des cultivars tardifs et leur indice de récolte est souvent supérieur (Isaa, 2013).
La photopériode peut aussi affecter le temps nécessaire à la floraison du maïs. Le maïs est classé comme une plante de jours courts. Après une période juvénile insensible au photopériodisme, la floraison et retardée par des photopériodes supérieures à 12,5 heures (Kiniry et al., 1983). Il existe une variabilité génétique pour la longueur de la photopériode critique au-dessous de laquelle la floraison n’est pas affectée. La plupart de cultivars tropicaux sont photosensibles, mais le degré de sensibilité varie énormément (de 1 à 12 jours de retard de l’anthèse par heure d’allongement de la durée du jour). Les cultivars tempérés ont tendance à être à la fois plus précoces et moins sensibles à la photopériode, ce qui leur permet de compléter leur cycle dans un laps de temps relativement court.
Le maïs est une plante unique du fait de la séparation de ses fleurs dans l’espace, les mâles sur la panicule terminale et les épis (femelles) latéraux. La panicule accomplit la majorité de développement avant la période de rapide développement des épis. Il semble que l’épi, en tant qu’organe formé en dernier, soit comparativement moins compétitif dans l’utilisation
d’éléments nutritifs. Une influence hormonale associée à la dominance apicale peut favoriser la croissance de la panicule et des feuilles supérieures, au détriment des pousses latérales surtout lorsque la densité des plantes est élevée. La sélection de panicule de taille réduite chez le maïs tropical a été associée à une augmentation du nombre de grains par plante et du rendement (Ficher et al., 1987). Ceci appuie l’hypothèse selon laquelle la panicule et l’épi sont en compétition pour les éléments nutritifs durant la phase critique de floraison et du rendement.
Beaucoup d’études ont montré que la disponibilité en éléments nutritifs aux alentours de la floraison est un facteur critique pour la production du rendement en grain. Ceci est difficile à comprendre, car la capacité photosynthétique instantanée du maïs est souvent à son maximum vers la floraison, et dans las autres tissus (Goldsworthy, 1984). Ce phénomène peut être dû au fait que les hormones produites à cette période par les premières fleurs affectent la croissance de suivantes. Les fleurs ayant un avantage initial, maintiennent cet avantage durant les premières étapes de la croissance du grain, entrainant ainsi un certain taux d’avortement des fleurs qui se forment plus tardivement. Le maïs serait plus sensible que les autres cultures à cet effet, peut être parce que les fleurs du sommet sont fécondées en dernier et que la séparation d’inflorescence peut réduire leurs chances d’être fécondées. Même quand la pollinisation est réussie, une mauvaise alimentation en éléments nutritifs peut entrainer l’avortement des fleurs fertilisées ou des épis entiers (Westgate, 1994).
1.1.2.4. Remplissage du grain
Une fois le nombre de grains établis, le rendement final dépend de la quantité d’éléments nutritifs stockés ou en cours de production. Il n’existe pas chez le maïs de connexions vasculaires entre les grains et la rafle. Les hydrates de carbone et les matières assimilables s’accumulent dans l’espace libre sous les grains en cours de développement et migrent dans les grains en suivant un gradient de diffusion ; une conséquence importante de ceci est que des déséquilibres dans la fourniture de différents constituants du grain peuvent limiter son développement. Par exemple, dans le cas de faible disponibilité en azote, la croissance du grain dépend d’une stricte stœchiométrie entre carbone (C) et azote (N) (Below, 1997). Si les hydrates de carbone solubles s’accumulent en excès dans le grain, la pression osmotique au niveau du pédicelle peut devenir trop élevée et empêcher toute nouvelle entrée de C, jusqu’à
ce que d’autres éléments d’hydrates de carbone solubles en constituent de grains, diminuant ainsi la pression osmotique (Porter et al., 1987).
Bolaños (1995) a montré que chez des maïs tropicaux de maturité semblable, la durée relative de la période de remplissage des grains est plus longue chez les hybrides que chez les variétés en pollinisation libre. Le rendement était lié à la durée de remplissage du grain.
1 .1.2.5. Dessiccation du grain
Sous les tropiques, le maïs est généralement récolté vers 20% d’humidité, c'est-à-dire à une humidité beaucoup plus basse que celle où la maturité physiologique est généralement atteinte (30 et 35%). Dans les régions tempérées, la vitesse de dessiccation est un facteur critique pour le choix d’une variété car la récolte doit êtres achevée avant l’hiver. Les variétés tempérées sont caractérisées par de spathes fines et ouvertes à maturité pour accélérer la dessiccation. Sous les tropiques, l’état sanitaire et la protection du grain sont généralement plus importants que la vitesse de dessiccation, et les maïs tropicaux sont caractérisés par des spathes épaisses et serrées avec une bonne couverture du sommet de l’épi.
Les spathes ouvertes favorisent l’entrée des insectes et de maladies, ce qui peut être particulièrement dommageable quand les grains sont stockés à la ferme, dans des lieux où le contrôle de la température et de l’humidité est réduit. L’implication physiologique de ces différences est que, chez les maïs tropicaux, la spathe représente une partie importante de la matière sèche produite. Les éléments nutritifs s’accumulent dans les spathes comme ils s’accumulent dans l’épi et la panicule ; elles entrent donc en concurrence directe avec ceux-ci. Les spathes servent aussi à stocker des hydrates de carbone solubles qui peuvent être utilisés pour le remplissage du grain. L’impact relatif de ces deux rôles des spathes sur le rendement en grain n’est pas très clair (Paliwal, 2002).
1.1.3. Production
En ASS, le maïs est produit principalement pour l'alimentation humaine, soit frais ou produit transformé. En outre, le maïs est produit pour l'alimentation animale et les utilisations industrielles telles que l'amidon, la farine, l'éthanol, le sirop de cuisson et croustillante. Selon le FAOSTAT (2013) un total de 885,3 millions de tonnes métriques de maïs a été réalisé à travers le monde. Sur ce total, l'Afrique a contribué à 65,7 millions de tonnes soit 7,2%. La faible contribution de l'Afrique à la production de maïs est principalement due au faible rendement moyen par unité de surface, au manque de connaissances scientifiques et pratiques adéquates (Law-Ogbomo et Remison, 2008) et à la faible utilisation de fertilisants (Fakorede et al., 2001). Selon l'étude réalisée par CIMMYT (2009), la demande pour le maïs dans les pays en développement dépassera celle de blé et de riz à l'horizon 2020. Par ailleurs, entre les périodes 1995 et 2020 la consommation mondiale et de l’Afrique subsaharienne du maïs devrait augmenter de 50% et 93% respectivement, indiquant l'importance de la culture à la fois en Afrique et dans le monde.
1.1.4. Culture et fertilisation
1.1.4.1. Culture de maïs
Le maïs présente une large diversité agro-morphologique. Son cycle du semis à la maturité varie de deux à onze mois, le nombre de ses feuilles de 8 à 48, la hauteur de sa tige de 0,6 à 6 mètres. Certaines variétés produisent plus de quatorze talles par plante. L'épi, long de 2,5 à 30 centimètres, peut comporter 8 à plus de 20 rangées de grains). La couleur des grains va du blanc au noir, en passant par le jaune, l'orange, le rouge, le vert et le bleu. Les rendements moyens sont de l'ordre de 0,5 tonne par hectare dans les zones défavorables, mais atteignent plus de 12 tonnes par hectare dans les plaines du Middle West américain ; le record mondial étant de 25 tonnes par hectare. Malgré cette diversité, toutes les variétés de maïs appartiennent à la même espèce et sont inter-fertiles (Paliwal, 2002).
Le maïs possède une plasticité adaptative remarquable, qui, associée à un potentiel de production supérieure à celui des autres céréales, explique son succès dans les pays développés. Après les Etats-Unis, il a conquis des vastes étendues en Europe, et progresse à
présent fortement en Asie. Il s'agit d'une importante source de protéines et d'énergie pour la consommation humaine et animale, et une source de matières premières pour l'industrie (Paliwal, 2002).
Le maïs tropical est à son tour, d’après son environnement, divisé en 3 sous-classes : basses terres, moyenne altitude et hautes terres (Dowswell et al., 1966). Pour sa culture, le maïs doit être en plein soleil pour que soit garantie une photosynthèse efficace. Ainsi, la durée du jour et l'ensoleillement déterminent la période de croissance. Généralement, chaque cultivar est adapté aux contraintes spécifiques de l'endroit où il est cultivé. Des températures élevées sont requises depuis le semis jusqu'à la floraison, tandis que durant la maturation, des conditions un peu plus fraîches peuvent être acceptées.
1.1.4.2. Fertilisation de maïs
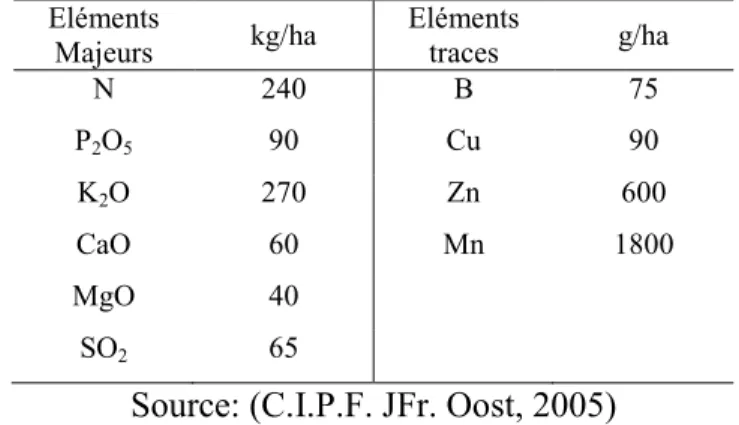
Une culture de maïs qui produit quatre tonnes de grains nécessite environ 100 kg d’azote (N), 18 kg de phosphore (P) et 68 kg de potasse (K) (Sanchez, 1976). Les besoins du maïs pour boucler son cycle sont approximatifs aux valeurs présentées dans le tableau 1.2.
Tableau 1.2. Les besoins en minéraux du maïs pour boucler le cycle Eléments
Majeurs kg/ha Eléments traces g/ha
N 240 B 75 P2O5 90 Cu 90 K2O 270 Zn 600 CaO 60 Mn 1800 MgO 40 SO2 65
Source: (C.I.P.F. JFr. Oost, 2005)
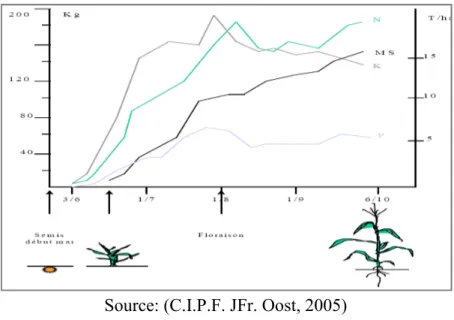
Chacun de ces éléments a son rôle pour la culture. Un apport optimum de ceux-ci contribue à atteindre le meilleur rendement. L’apport de l’azote influence non seulement la croissance juvénile du maïs mais également sa teneur en azote brute digestible en fin de saison ; le phosphore intervient dans le mécanisme de la maturation et influencera donc directement la teneur en matière sèche ; quant à la potasse, elle conduit à la formation de l’épi et des grains.
Le maïs a un développement très rapide, l’élément fertilisant doit donc être disponible au moment où la plantes en a besoin. La majeure partie des éléments nutritifs est prélevée durant la période relativement courte qui couvre la formation de l’épi et plus précisément 10 jours avant l’apparition des fleurs jusqu’à 25 à 30 jours après. Durant cette période, la plante absorbera 70% à 75% d’azote et 2/3 de ses besoins en phosphore et potasse. On estime que l’absorption des éléments majeurs se réparti sur la saison comme l’indique le tableau 1.3.
Tableau 1.3. Absorption des éléments majeurs au cours d’une saison 1er mois 2e mois 3e mois 4e mois
N faibles 40% 50% 10%
P faibles 25% 50% 25%
K faibles 70% 30% -
Source: (C.I.P.F. JFr. Oost, 2005)
Le système racinaire est capable d’absorber d’éléments nutritifs pendant tout le cycle de la plante, mais le taux d’absorption décroit pendant la fin du remplissage des grains, en raison de la sénescence des feuilles de la base de la plante. La réponse du maïs à la fertilité azotée est généralement positive et linéaire jusqu’à un taux d’absorption relativement plus élevé que pour les autres cultures. Le système racinaire du maïs, relativement grossier, n’est pas très compétitif comparé à des espèces ayant des racines plus fines, comme c’est le cas pour certaines graminées fourragères. La concentration en azote des feuilles est relativement faible (1 à 4%) (Pearson et Hall, 1984) comparée à des céréales en C3 comme le blé. L’efficacité de l’utilisation de N dans la photosynthèse est plus grande chez le maïs, aussi sa concentration dans les feuilles comparativement plus faible, ne limite-elle pas sa productivité par rapport aux autres cultures (Sinclair et Horie, 1989). A des niveaux trop élevés de fertilisation azotée l’efficacité de conversion (EC) est fortement diminuée. Une culture à une fertilisation azotée élevée, qui produit 32% plus qu’un témoin, avait une EC de 0,7 g/MJ (gramme de matière sèche produite par mégajoule de radiation interceptée active pour la photosynthèse), comparée à 1,3g/MJ pour le témoin dans l’étude conduite par Muchow (1994). La sensibilité de l’EC à des disponibilités en azote faible est plus forte que la sensibilité de développement de la surface foliaire (Muchow et Sinclair, 1994).
La quantité d’azote qui migre des tissus végétatifs vers l’épi durant le remplissage des grains peut varier dans des grandes proportions. On considère que 20 à 60% d’azote total des grains provient de l’absorption réalisée avant l’anthèse. L’azote stocké dans les tiges est le premier à être transféré vers l’épi. La quantité d’azote mobilisée dépend à la fois de la variété, de la quantité et du moment d’application. Les indices d’azote récolté (kg de N dans le grain/kg de
N dans la plante entière, sans les racines) sont plus élevés que les indices de matière sèche récoltée et peuvent être de l’ordre de 0,6 à 0,8 chez les maïs tardifs tropicaux cultivés dans des conditions optimales (Issa, 2013). Le modèle de distribution du phosphore dans la plante est semblable à celui de l’azote, excepté qu’une plus grande proportion des besoins en phosphore (P) se manifeste après la floraison. La majorité du potassium nécessaire à la culture est absorbée avant la floraison et la plupart du potassium reste dans les pailles après la récolte. Il ne semble qu’il y ait pas de différences significatives dans la nutrition minérale des maïs tempérés et tropicaux, ainsi les scientifiques travaillant en milieux tropicaux peuvent bénéficier des résultats obtenus pour les maïs tempérés.
1.1.4.3. Besoins en eau
Seuls quelques trois pourcents de radiations incidentes arrivant sur la culture sont utilisées pour la photosynthèse. Une partie de l’énergie restante chauffe le feuillage et cette chaleur est dissipée par la transpiration (évapotranspiration d’eau par les feuilles). Un millimètre d’eau est évaporé par 2,4MJ de radiation solaire. Un maïs bien arrosé transpire environ 350 g d’eau par gramme de matière sèche produite au-dessus du sol (Hay et Walker, 1989) auxquels il faut ajouter l’eau évaporée par le sol. Le grain constitue une fraction de la matière sèche produite, ce qui signifie qu’une culture bien arrosée utilise de 800 à 1000 g d’eau par gramme de grain produit.
Comme le dioxyde de carbone pénètre dans les feuilles à travers la même ouverture (les stomates) que l’eau quittant les feuilles, il existe une bonne relation entre la quantité d’eau transpirée et la matière sèche produite par la culture. Des facteurs environnementaux ont une action sur le degré d’ouverture de stomate, permettant ainsi une diminution de la quantité d’eau perdue par la culture. En fonction des radiations, de température et d’humidité relative, une culture de maïs transpirant librement, transpire environ 80 à 90% de l’évaporation potentielle. Dans ces conditions, la température de la feuille et généralement légèrement inférieure à celle de l’air et la quantité d’eau utilisée par la transpiration par unité de surface de la culture. Quand la transpiration est réduite par la fermeture des stomates, la culture se déconnecte de l’environnement (Jones, 1993) ; la température de feuilles augmente alors, ce qui peut avoir des effets nocifs sur le métabolisme de la plante. De plus, une fermeture prolongée des stomates réduit la quantité de CO2 disponible pour la photosynthèse, diminuant
ainsi l’efficacité de conversion. Les facteurs environnementaux affectant la fermeture des stomates sont l’humidité du sol et la demande de transpiration.
Le progrès génétique réalisé sur la productivité induit mécaniquement un besoin en eau accru, sachant qu'il est quasiment certain que l'efficience intrinsèque de l'eau n'a pas été modifiée entre les variétés anciennes et modernes de maïs (Barrière, 2001). Ainsi, en conditions non limitant en eau, nutriments et lumière, un maïs (ou un sorgho) produit 40 kg de matière sèche (MS) par mm d'eau utilisé par hectare, alors qu’un blé ou un ray-grass ne produit que 25 kg MS/mm/ha, et une luzerne (ou une légumineuse) 20 kg MS/mm/ha (Marine, 2000).
Au cours de la période du 1950 à 2000, l'évolution des variétés de maïs se caractérise par des progrès exceptionnels sur la productivité (et inconnus sur l’ensemble des autres plantes cultivées), en stabilité de la productivité, en résistance à la verse et aux pourritures de tiges (Barrière et Emile, 2000a).
Le maïs demande beaucoup d'eau, en raison de la grande quantité de matière organique produite lors de la croissance. Le besoin en eau de maïs cultivé dans les plaines est d’au moins 500 mm de précipitations bien réparties tout au long de la saison (Norman et al., 1995). Il exige un maximum d'humidité durant la période de la formation de l'aigrette et des soies. Son coefficient de transpiration est de plus ou moins 280 à 350, ce qui signifie que pour la synthèse d'un gramme de matière sèche, le maïs transpire 280 à 350 grammes d'eau. Pendant la période de croissance intensive, comme les deux semaines qui se situent avant l'apparition de l'aigrette, une seule plante de maïs transpire 2 à 4litres d'eau en 24 heures. Il s'ensuit qu'un champ d'un hectare comptant 50.000 plantes produit 100 à 200 tonnes d'eau par jour.
Concernant les conditions climatiques et les méthodes de culture, le maïs consomme 60 à 90 tonnes d'eau pour produire 100kg de grains. Les conditions d'humidité favorables au maïs sont réunies, lorsque l'humidité du sol est en surplus au niveau des racines et si le total des précipitations, devant atteindre 400mm au moins, est favorablement distribué durant la période de croissance. La teneur en humidité du sol pour la croissance et le développement favorable à un rendement élevé doit atteindre 60 à 70% de la capacité au champ. Dans des conditions de sécheresse, le taux de croissance est diminué, la période de fécondation est retardée, et la formation ainsi que le remplissage des grains sont considérablement réduits, entraînant par là une diminution de rendement (Ristanovic, 2001).
La profondeur de semis est de 5 à 8cm si l’humidité est adéquate à cette profondeur. La levée rapide et régulière des plantules se fait alors en quatre ou cinq jours. Cette durée augmente
lors du semis plus profonds. Dans certains environnements, par exemple dans les hautes terres du Mexique, la semence est normalement mise à des profondeurs de 12 à 15cm dans le but d’atteindre une humidité suffisante pour assurer une bonne germination (Paliwal, 2002).
1.1.5. Importance dans l’alimentation
L'attractivité de maïs comme aliment de base est largement due à son rôle diversifié comme une source de nourriture pour les humains et les animaux. La suite du traitement conduit à son utilisation comme épaississants alimentaires, des édulcorants et des huiles
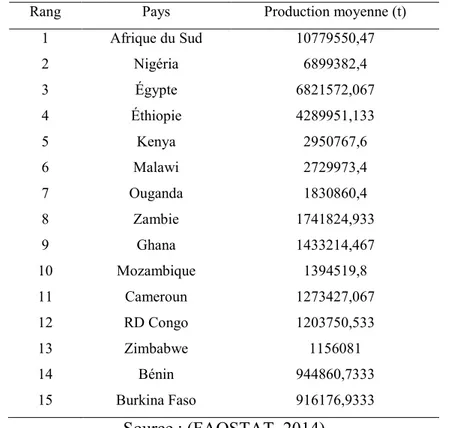
La consommation annuelle du maïs par habitant en Afrique australe va de 138 kg à 195 kg au Swaziland en Afrique du Sud (CIMMYT, 1999), tandis qu’en Afrique orientale elle varie de 40 kg au Burundi à 105 kg au Kenya. En Afrique le maïs est cultivé par les agriculteurs de petite et moyenne dimension qui cultivent 1ha ou moins dans des conditions de systèmes d’intrants extrêmement faibles avec des rendements moyens du maïs à 1,3 t ha-1 (Bänziger et Diallo, 2004). Les pays d'Afrique subsaharienne ne produisent pas assez de maïs pour répondre à leurs besoins et doivent donc importer environ trois millions de tonnes de maïs par an. L’Afrique du Sud conduit la production de maïs du continent, suivie par le Nigéria (Tableau 1.4). Le rendement moyen par unité de surface est de 5,2 tonnes ha-1 pour le total mondial, 1,9 tonne ha-1 pour l'Afrique en 2011 (FAOSTAT, 2013). L'exception est l’Egypte, où le système de production est soutenu par l’irrigation (FAOSTAT, 2010).
Tableau 1.4. Les 15 premiers producteurs de maïs en Afrique entre 2000 et 2014
Rang Pays Production moyenne (t)
1 Afrique du Sud 10779550,47 2 Nigéria 6899382,4 3 Égypte 6821572,067 4 Éthiopie 4289951,133 5 Kenya 2950767,6 6 Malawi 2729973,4 7 Ouganda 1830860,4 8 Zambie 1741824,933 9 Ghana 1433214,467 10 Mozambique 1394519,8 11 Cameroun 1273427,067 12 RD Congo 1203750,533 13 Zimbabwe 1156081 14 Bénin 944860,7333 15 Burkina Faso 916176,9333 Source : (FAOSTAT, 2014)
Contrairement au monde développé où le maïs est principalement produit pour l'alimentation animale, la consommation humaine du maïs en ASS est estimée à environ 70% (Aquino et al., 2001). La recherche indique que 20% des calories alimentaires mondiales et 15% de toutes les protéines vivrières sont fournies par le maïs. Cependant, la qualité du maïs normal et la plupart de protéine de céréales est pauvre car elle manque des acides aminés essentiels, la lysine et le tryptophane (Bhatia et Rabson, 1987).
La carence en acides aminés essentiels dans le maïs normal provoque une grave malnutrition en protéines et des problèmes connexes pour les personnes ayant des besoins élevés en protéines, par exemple, les jeunes enfants, les femmes enceintes ou allaitantes et les malades dans les communautés où le maïs est un aliment de base et souvent une source majeure de protéine (Pixley et Bjarnason, 2002). En Afrique subsaharienne où le système de culture du maïs est dominant, le taux de malnutrition en particulier pour les enfants d'âge préscolaire est élevé. Hyman et al. (2008) ont rapporté que la prévalence du retard de croissance est de plus de 40% dans les zones où le maïs est un régime dominant. En outre, la proportion de personnes pauvres (qui vivent en dessous de 2$ US par jour) dans les communautés agricoles de maïs de l’ASS est d'environ 65% (Wood et al., 2010) ce qui implique que les sources de protéines comme la viande, le lait et les œufs sont inabordables.
1.1.6. Place du maïs en RDC
Après le manioc, le maïs est la principale denrée alimentaire produite en RDC, loin devant le riz. Sa consommation est surtout importante dans les savanes du nord du pays et dans les provinces du sud : le Katanga et les deux Kasaï. Il est produit dans l’ensemble du pays mais quatre provinces représentent près de 70% de la production nationale : le Katanga (23%), le Bandundu (17%), le Kasaï Occidental (17%) et le Kasaï Oriental (13%). Le maïs est surtout produit par les exploitations traditionnelles de très petite taille, centrées sur l’autoconsommation et la sécurité alimentaire familiale, qui n’utilisent aucun intrant et qui cultivent le maïs le plus souvent en association avec le manioc et l’arachide.
Il existe cependant des exploitations de plus grande taille produisant en culture pure pour le marché, surtout dans le Katanga et les provinces du Kasaï.
Le tableau 1.5 présente l’évolution de la production de maïs en tonne par province entre 1995 et 2014. Le manque de données entre 1997 et 1999 se justifierait par l’instabilité politique et économique dans le pays.
Tableau 1.5. Evolution de la production de maïs par province (en tonnes)
Les activités de recherche sur le maïs n’ont pas bénéficié des appuis récemment apportés au manioc et au riz. L’INERA a continué à produire de très petites quantités de semences de pré-base et de pré-base de maïs. La production des semences commerciales dites certifiées est actuellement entre les mains d’un nombre important d’acteurs disparates (fermes privées, fermes semencières dites autonomes et réseau d’ONG). Mais le manque de suivi et de contrôle ne permet pas d’en assurer la qualité. Les activités d’amélioration variétale et de multiplication des semences demandent à être soutenues.
1.1.7. Notion de la sécheresse en agriculture
Tardieu (1996) définit la sécheresse comme étant une période prolongée sans précipitation qui peut résulter dans la réduction de la teneur en eau du sol et, ainsi, causer le déficit hydrique chez la plante. Cette sécheresse peut également être définie en termes soit de statut de l’eau externe aux alentours de la plante (sol, air) ou bien le statut de l’eau interne dans les tissus. Ceci signifie qu’il y a une balance négative entre l’offre ou la disponibilité en eau et la demande de la plante.
Le changement climatique pourra continuer à accroitre encore le nombre de personnes souffrant de la faim au cours des prochaines décennies, en raison des nouveaux défis posés à l’agriculture et à la production vivrière. Partout dans le monde, les températures pourraient augmenter de 6°C d’ici 2050. Des sécheresses et des inondations plus graves et plus fréquentes ainsi qu’une pression plus forte des ravageurs et des maladies sont les principaux défis induits par le changement climatique à l’agriculture et à la production vivrière (FAO, 2007).
D’après la FAOSTAT (2010), 90% des paysans en Afrique dépendent uniquement de la pluie, qui est devenue imprévisible, pour stabiliser la production des cultures. Peu d’agriculteurs ont accès á l’irrigation. Seulement 4 % de terres de champs en Afrique subsaharienne sont irriguées comparativement á 37 % en Asie.
Les changements climatiques observés ces dernières années et qui sont caractérisés par l’alternance de la sécheresse et des inondations ; le raccourcissement de la saison de pluie et la mauvaise répartition de cette dernière, viennent aggraver une situation qui était déjà très
préoccupante. Et cette situation ne fera qu’accentuer le déficit de la production du maïs, si des mesures et des stratégies efficaces ne sont pas développées et mise en place.
Une production agricole fiable nécessite un environnement stable. Une variabilité climatique accrue, conjuguée à une tendance au réchauffement systématique, menace la stabilité de cet environnement à prédisposition agricole corollaire à une insécurité alimentaire. Les outils de gestion des risques climatiques destinés à accroître la résilience consistent notamment en de meilleurs systèmes d’alerte rapide et des régimes d’assurance contre les intempéries, qui informent, préviennent, réduisent et répartissent les risques de façon à ce que les agriculteurs et les secteurs commerciaux et industriels agro-alimentaires ne pâtissent pas dans une mauvaise saison.
L’Afrique tire seulement 10% de sa production alimentaire de terres irriguées et c’est essentiellement dans le Nord, où l’eau se fait de plus en plus rare. Environ 95% des terres cultivées d’Afrique subsaharienne relèvent de l’agriculture pluviale. Ce continent ferait bien de développer l’irrigation pour rattraper l’Asie. L’agriculture est actuellement le secteur qui consomme le plus d’eau (environ 70%). Comme la demande d’autres usagers de premier plan est à la hausse, il est important d’augmenter l’efficacité de techniques de gestion hydrique pratiquée actuellement ou innovées.
1.1.8. Contraintes de la culture du maïs en Afrique subsaharienne
Plusieurs facteurs biotiques et abiotiques limitent la production de maïs et de la productivité dans les pays de l'Afrique subsaharienne (Badu-Apraku et al., 2003). Les facteurs biotiques limitant la production de maïs dans la région comprennent les insectes nuisibles, les maladies et les mauvaises herbes parasites. Les principaux facteurs biotiques et abiotiques limitant la production de maïs vont de la fécondité à la sécheresse de sol, et ceux parmi les défis les plus importants de la production de maïs, la sécurité alimentaire et la croissance économique de l’Afrique (CIMMYT, 2003 ; Bänziger et Diallo, 2004). Des faibles niveaux d'azote (N) dans les sols comme un facteur majeur limitant le rendement retrouvé souvent dans les champs des agriculteurs dans les tropiques où les engrais ne sont pas couramment utilisés et de la matière organique est rapidement minéralisée.
La majorité des agriculteurs dans les tropiques produit du maïs dans des conditions pluviales et sont vulnérables à la sécheresse. Bien que la sécheresse à toute étape de la croissance et du développement des cultures affecte la production, le plus grand impact se produit autour de la floraison (Edmeades et al., 1992). L'incidence de stress hydrique dans la culture du maïs devrait augmenter en partie due au changement climatique et le déplacement du maïs à des environnements marginaux par les cultures à haute valeur ajoutée (Bänziger et al., 2000). L'adoption de cultivars utilisant plus efficacement l’azote ainsi que tolérant les sécheresses récurrentes que connaît une région pourra atténuer les défis de stress abiotiques dans la culture du maïs (Diallo et al., 2004).
Plusieurs études ont montré qu'une bonne compréhension des facteurs environnementaux et génétiques à l'origine de l'interaction ainsi que l'évaluation de leur importance dans le système Génotype et Environnement (G x E) pourraient avoir un grand impact sur l'amélioration des plantes (Basford et Cooper, 1998). L’interaction G x E se produit le plus souvent lorsque les génotypes sont évalués à travers des environnements (Becker et Leon, 1988 ; Kang, 1998). Magari et Kang (1993) ont constaté que la contribution des différents facteurs environnementaux à la stabilité de rendement du maïs a eu des bons résultats dans les essais multi-locaux. Les effets de G x E sont plus apparents dans les essais d'environnements multiples qui ont trois objectifs principaux :
a) d'estimer avec précision les niveaux de rendement prévisionnels basés sur des données expérimentales ;
b) de déterminer la stabilité de rendement et l'adaptation des génotypes dans des environnements ; et
c) de fournir des indications fiables pour sélectionner les meilleurs génotypes ou traitements agronomiques pour la plantation dans les années à venir à de nouveaux sites (Crossa, 1990).
1.2. Les stress abiotiques affectant la culture de maïs 1.2.1. Stress hydrique
Le déficit hydrique est une contrainte permanente de la production agricole. Il est à l'origine des pertes de production agricole dans de nombreuses régions. Les risques du manque d’eau sont et deviendront de plus en plus fréquents et persistants, à l'avenir, par suite des changements climatiques causés par l'effet de serre (Witcombe et al., 2009).
Passioura (2004) définit le déficit hydrique comme étant les circonstances dans lesquelles les plantes accusent une réduction de croissance et de production suite à une alimentation hydrique insuffisante dite stress hydrique.
1.2.1.1. Symptômes de stress hydrique 1.2.1.1.1. Avant floraison
- Enroulement des feuilles du cornet. La plante se protège en fermant ses stomates pour limiter l’évapotranspiration ;
- Coloration bleue des feuilles du cornet
1.2.1.1.2. A la floraison
- Retard de la floraison femelle ; - Problème de fécondation
Dans le cas de stress hydrique ou thermique sévères, le délai entre l’émission du grain de pollen et l’émergence des soies (Intervalle de floraison) augmente et en conséquence, une faible quantité de grain de pollen viable au moment où les soies sont réceptives et donc une fécondation réduite. Selon la période de stress, l’épi peut présenter une bonne fécondation à la base et faible au sommet, ou une répartition dispersée des grains, comme le montre la figure 1.1.
1.2.1.1.3. Après floraison
- Avortement des grains
- Echaudage des grains du haut de l’épi ou quelques fois sur ou plusieurs rangs (épi en banane) (figure1.2.)
- Perte de PMG
Figure 1.2. Conséquence du déficit hydrique au moment de floraison : avortement des ovules. (Source : Kabongo et al., 2016)
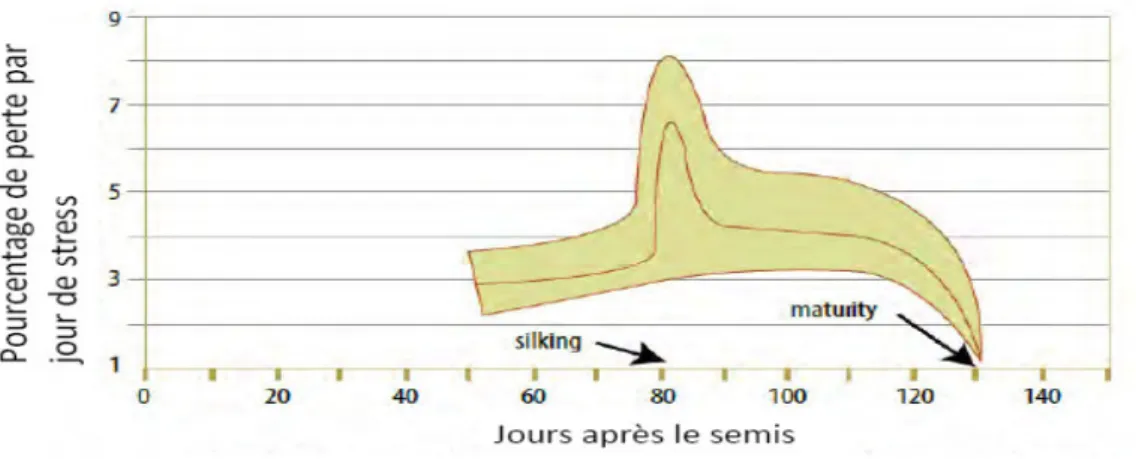
1.2.1.2. Situation à risque
- Petites terres et toutes les situations non ou mal irriguées, à faibles réserve hydrique
- Retard dans la mise en place du système d’irrigation - Semis de maïs après précédent récolte tardive
La figure 1.3, montre l’évolution de perte de rendement par jour de stress hydrique à différentes époques de saison. Les pertes sont plus importantes lorsque la plante est fortement stressée au moment de floraison femelle.