Time-optimal control of piecewise affine bistable gene-regulatory networks: preliminary results
Texte intégral
Figure
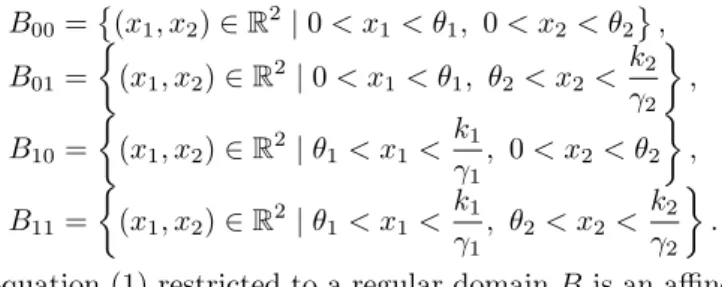
Documents relatifs
CAPN3 嫩度 Tendreté 2006 CSIRO/MLA/Beef CRC, Australie Genetic Solutions P/L CAST 嫩度 Tendreté 2002 CSIRO/MLA/Beef CRC, Australie Genetic Solutions P/L GH1 大理石纹 Persillé
Abstract: As a first approach to the study of systems coupling finite and infinite dimensional natures, this article addresses the stability of a system of ordinary
c Transfer curves (channel and gate current as a function of applied gate bias) for an HgTe nanocrystal array deposited on a nanotrench fabricated on a LaF 3 substrate used as a
Elle ne modifie en rien les termes quant aux modalités d’intervention financière dont disposent les Départements pour impulser une politique publique en faveur des sports de nature
When faces were presented centrally, regions along the ventral visual pathway (occipital cortex, fusiform gyrus, bilateral anterior temporal region) were more activated by fear than
This text is based on notes (taken by R. Thangadurai) of three courses given at Goa University on December 15 and 16, 2014, on Algebraic independence of Periods of Elliptic
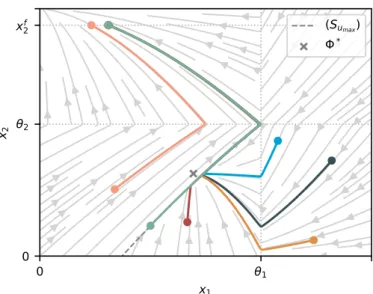
The focus of this paper is the synthesis of suboptimal state feedback controllers for continuous-time optimal control problems (OCP) with piecewise-affine (PWA) dynam- ics and
Average baseline latencies to first lever press for AM, MD, and PM sessions, and average latencies for sessions that followed AM, MD, and PM skip session probes, as well as the
