Does structural sensitivity alter complexity–stability relationships?
25
0
0
Texte intégral
Figure
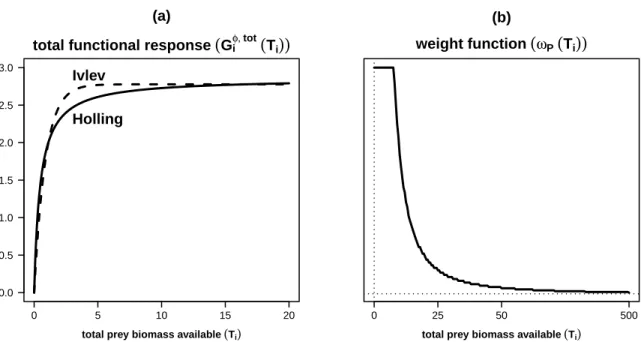



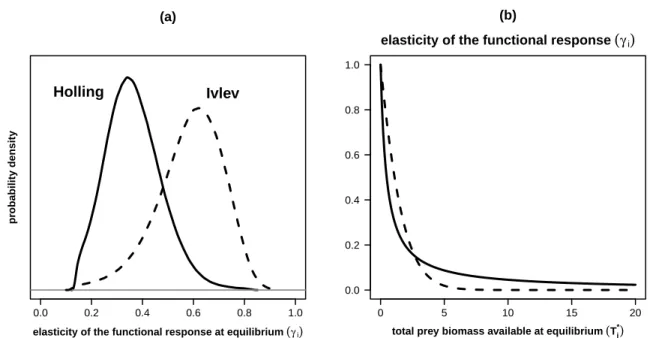
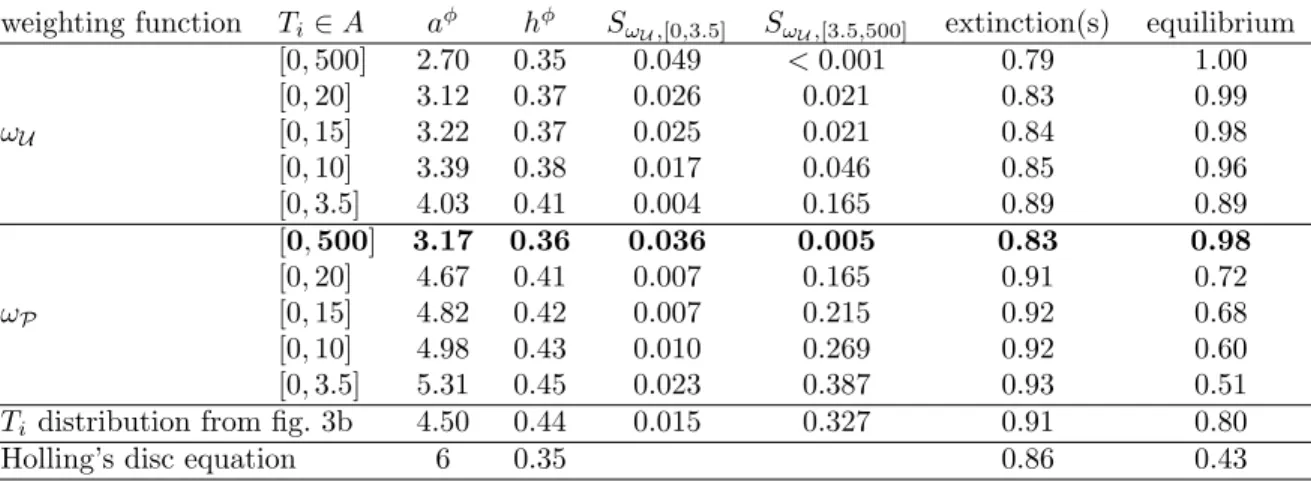
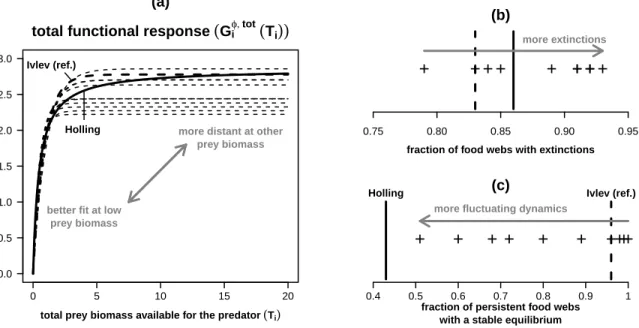
+5

Documents relatifs
[r]
Dans tout ce qui suit, les longueurs seront exprimées en unités de longueur
[r]
This report has been prepared by the World Health Organization Regional Office for the Western Pacific for governments of Member States in the Region and for the
[r]
[r]
[r]
[r]
