Modelling forest recolonization by an epiphytic lichen using a landscape genetic approach
17
0
0
Texte intégral
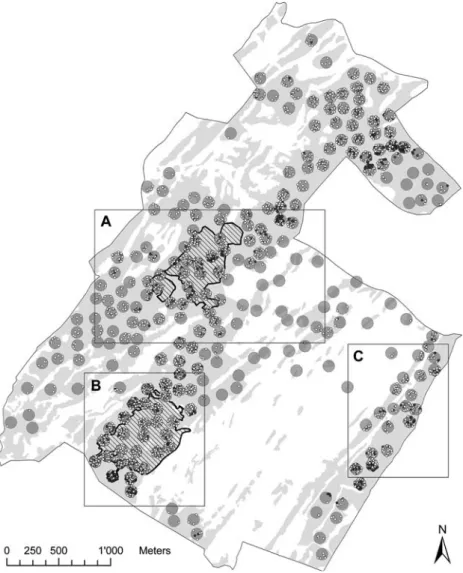
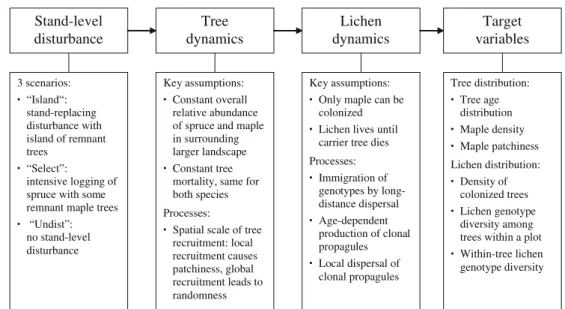
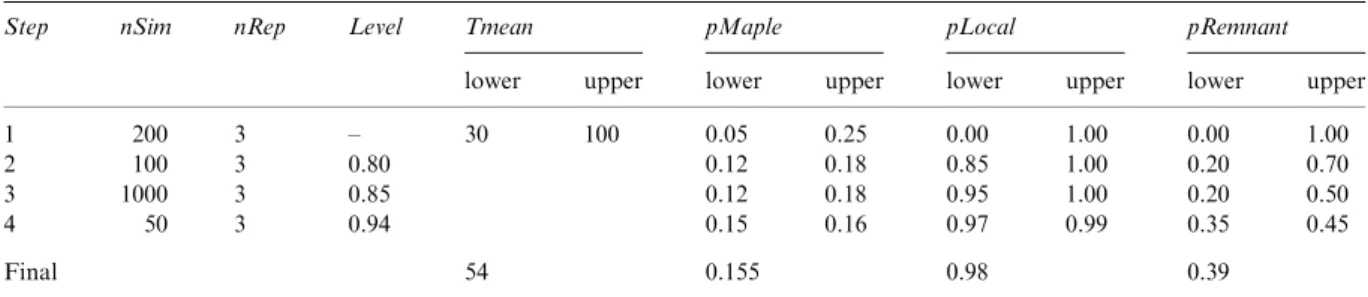
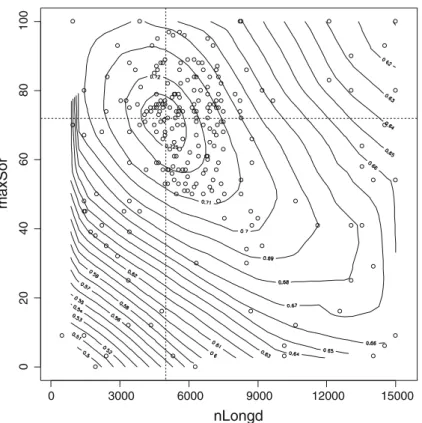
Figure


+5


Documents relatifs