HAL Id: tel-01556472
https://tel.archives-ouvertes.fr/tel-01556472
Submitted on 5 Jul 2017HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Modélisation de l’organisation fonctionnelle de la lecture
chez le sujet sain : Etude en EEG-HR par reconstruction
de sources et modèles causaux dynamiques.
Applications aux patients épileptiques
Gaëtan Yvert
To cite this version:
Gaëtan Yvert. Modélisation de l’organisation fonctionnelle de la lecture chez le sujet sain : Etude en EEG-HR par reconstruction de sources et modèles causaux dynamiques. Applications aux patients épileptiques. Psychologie. Université de Grenoble, 2012. Français. �NNT : 2012GRENS044�. �tel-01556472�
Université Joseph Fourier / Université Pierre Mendès France / Université Stendhal / Université de Savoie / Grenoble INP
THÈSE
Pour obtenir le grade de
DOCTEUR DE L’UNIVERSITÉ DE
GRENOBLE
Spécialité:SciencesCognitives,Psychologie & Neurocognition
Arrêté ministériel : 7 août 2006 Présentée par
Gaëtan Yvert
Thèse dirigée par Monica Baciu et Olivier David
préparée au sein du Laboratoire de Psychologie et NeuroCognition et Grenoble Institut des neurosciences dans l'École Doctorale Ingénierie pour la Santé, la Cognition et l’environnement
Modélisation de l’organisation fonctionnelle de
la lecture chez le sujet sain. Etude en EEG-HR
par reconstruction de sources et modèles
causaux dynamiques. Applications aux patients
épileptiques.
Thèse soutenue publiquement le 18 juin 2012, devant le jury composé de :
M. Jean-Luc Schwartz, Directeur de Recherches au CNRS,
GIPSA-LAB, Département Parole & Cognition, CNRS UMR 5216 & Grenoble Universités, France. (Président)
Mme. Fanny Meunier, Directeur de Recherches au CNRS, laboratoire Dynamique du Langage, UMR5596, Lyon, France. (Rapporteur)
M. Christoph Michel, Professor, Functional Brain Mapping, University Medical Center, Genève, Suisse. (Rapporteur)
Mme. Catherine Tallon-Baudry, Directeur de Recherches au CNRS, LENA CNRS UPR640, France. (Examinateur)
M. Jeremy Mattout, Chargé de Recherches, INSERM U1028 -
CNRS UMR5292, Equipe DYCOG, Lyon, France. (Examinateur)
Mme. Monica Baciu, Professeur, Université Pierre Mendès-France, LPNC, UMR-CNRS 5105, Grenoble, France. (Directeur de thèse)
M. Olivier David, Chargé de Recherches, Fonctions cérébrales et neuromodulation, INSERM U 836, La tronche. (Directeur de thèse)
2
3
T
ABLE DES MATIERES
!"#$%&'()*)))))))))))))))))))+,)-./011,233,1/.)4235.66.)7777777777777777777777777777777777777777777777777)*8! !"! #$%&'()**$+,,$*('%-+,.'//'%0',%)12'3,%""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%44! "#! $%&'()*'+),(!-.*!')%.*!/)*0.11.*!2%)3')%.*!###########################################################################!44! 5#! $%&'()*'+),(!6)7%'%86)90.!-.!1'!/,).!/.(+%'1.#####################################################################!4:! ;#! ;,(*+%08+),(!2'%!1<.=27%).(8.!-.!1'!2>%'3)-.!6)7%'%86)90.!###############################################!4?! !!"! #$%&'()**$+,,$*('%-+,.'//'%0',%5)3,%"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%46! "#! @(+%,-08+),(!#########################################################################################################################!4A! 5#! B'!%.8,((')**'(8.!/)*0.11.!-.*!1.++%.*!C!-.!1<,DE.+!F!1'!1.++%.!##############################################!4A! ;#! B.!+%')+.3.(+!,%+6,&%'26)90.!#############################################################################################!4G! H#! H.!1<,%+6,&%'26.!F!1'!26,(,1,&).!########################################################################################!I4! J#! B'!%78027%'+),(!*73'(+)90.!################################################################################################!IA! K#! ;,(810*),(!*73'(+)90.########################################################################################################!:G! !!!"! 7&)(',,.,%3)890):*%'3%&'()**$+,,$*('%-+,.'//'%"""""""""""""""""""""""""""""""""""""""""""""""""""%;<! "#! @(+%,-08+),(!#########################################################################################################################!LM! 5#! ;,((.=),(*!D,++,3N02!.+!+,2N-,O(C!P.1'+),(*!'('+,3)90.*!.+!Q,(8+),((.11.*!################!L4! ;#! R,-S1.*!+67,%)90.*!.+!8,320+'+),((.1*!############################################################################!LT! H#! R78'()*3.!-.!1<)(+7&%'+),(!-.*!2%,8.**0*!D,++,3N02!.+!+,2!-,O(!##################################!LT! J#! @(Q10.(8.!-.*!8,((.=),(*!+,2N-,O(!-'(*!1'!%.8,((')**'(8.!/)*0.11.!#################################!L:! K#! ;,(810*),(!*0%!1.!3,-S1.!+67,%)90.!-.!Q'8)1)+'+),(!+,2N-,O(!############################################!LU! !"#$%&'()9! ):;<=0>060?2.)77777777777777777777777777777777777777777777777777777777777777777777777777777777)@*! !"! #=>/'(3&)9'*(>8?$/)@&$8?+'%""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%AB! "#! $%)&)(.!-.*!8,0%'(+*!3.*0%7*!#############################################################################################!?I! 5#! B.*!2,+.(+).1*!7/,907*!########################################################################################################!?:! ;#! R7+6,-,1,&).!-<'890)*)+),(!-.*!-,((7.*############################################################################!?L! !!"! #$%&'()*,3&.(3+)*%0'%,).&(',%""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%A6! "#! B.!2%,D1S3.!-)%.8+!###############################################################################################################!?A! 5#! B.!2%,D1S3.!)(/.%*.!############################################################################################################!AM! ;#! B'!%.8,(*+%08+),(!-.!*,0%8.*!.(!2%'+)90.!###########################################################################!AA! !!!"! C)0>/+,$3+)*%($.,$/'%0D*$5+E.'%FGHC%I%GD*$5+(%H$.,$/%C)0'//+*@J%""""""""""""""""""%6K! "#! @(+%,-08+),(!#########################################################################################################################!AG! 5#! V67,%).!#################################################################################################################################!UM! ;#! ;6,)=!-.*!3,-S1.*!C!H;R!.(!2%'+)90.!#################################################################################!UT! !L"! C>3?)0)/)@+'%'M8>&+5'*3$/'%"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%NO!
4 "#! ;6,)=!.+!3)*.!.(!21'8.!-.*!2'%'-)&3.*!###############################################################################!U:! 5#! W%,87-0%.!.=27%)3.(+'1.!####################################################################################################!U?! !"#$%&'()A))))))))))))))$,-<2.).BC;-2D.1<,6.)77777777777777777777777777777777777777777777777777777777777777)EE! !"! !*3&)0.(3+)*%"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%NK! !!"! P>,./3$3,%I%""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%NK! "#! "%+)81.!###################################################################################################################################!UG! 5#! P7*037!-.*!%7*01+'+*!###########################################################################################################!GM! !!!"! P>,./3$3,%,.&%/=+*Q/.'*('%0'%/$%Q&>E.'*('%/'M+($/'%"""""""""""""""""""""""""""""""""""""""""""""""""%KO! "#! P7*01+'+*!8,32,%+.3.(+'0=!###############################################################################################!G:! 5#! P7*01+'+*!*0%!1.*!%.8,(*+%08+),(*!-.!*,0%8.*!#######################################################################!G:! !"#$%&'()F))))))G23/533201)>.3)-;356<,<3)35-)6.3)35H.<3)3,213)77777777777777777777777777777777777)I@! !"! !*3&)0.(3+)*%"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%K6! !!"! #'%&>,'$.%0'%/$%&'()**$+,,$*('%-+,.'//'%0',%5)3,%""""""""""""""""""""""""""""""""""""""""""""""""%K6! "#! B'!%.8,((')**'(8.!,%+6,&%'26)90.!.+!1.=)8'1.!####################################################################!GU! 5#! B.!-78,-'&.!26,(,1,&)90.!##################################################################################################!GG! ;#! P78027%'+),(!*73'(+)90.!#################################################################################################!4M4! !!!"! #'%Q)*(3+)**'5'*3%0'%('%&>,'$.%""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%4<B! "#! B'!/,).!1.=)8,N*73'(+)90.!#################################################################################################!4M:! 5#! H.!1<.=)*+.(8.!-<0(.!8,((.=),(!%'2)-.!,88)2)+,NQ%,(+'1.!##################################################!4ML! ;#! H.!1<)32,%+'(8.!-.*!2%,E.8+),(*!+,2N-,O(!#######################################################################!4M?! H#! R,-S1.*!Q,(8+),((.1*!+67,%)90.*!-0!%7*.'0!-.!%.8,((')**'(8.!/)*0.11.!#########################!4MA! !L"! #$%()**'(3+-+3>%,3&.(3.&$/'%*'.&)9$*$3)5+E.'%,).,93'*0$*3%('%&>,'$.%"""""""""""""""%444! "#! X.0%,N'('+,3).!-.*!%7*.'0=!-.!*0D*+'(8.*!D1'(86.*!######################################################!44T! 5#! PY1.*!Q,(8+),((.1*!8,((0*!-.*!Q')*8.'0=!-.!Q)D%.*!D1'(86.*!#############################################!44I! L"! H)*(/.,+)*%,.&%/'%&>,'$.%/$*@$@+'&%"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%446! !"#$%&'()J)))))))))(C26.C32.).<)-;0-?,123,<201)K01/<2011.66.)77777777777777777777777777777777777)**I! !"! !*3&)0.(3+)*%""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%4R<! "#! Z)*+,%)90.!##########################################################################################################################!4TM! 5#! B<72)1.2*).!C!R'()Q.*+'+),(!.+!+67%'2).!##############################################################################!4TM! ;#! B<72)1.2*).!-0!1,D.!+.32,%'1!#############################################################################################!4T:! !!"! #$%&>)&@$*+,$3+)*%Q)*(3+)**'//'%0$*,%/=>8+/'8,+'%3'58)&$/'%""""""""""""""""""""""""""""""""%4RO! "#! B'!21'*+)8)+7!87%7D%'1.!########################################################################################################!4T:! 5#! B.*!-)QQ7%.(+*!378'()*3.*!-.!21'*+)8)+7!############################################################################!4TL!
5 ;#! B.*!-)QQ7%.(+*!2'++.%(*!-.!%7,%&'()*'+),(!2'%!*0D*+)+0+),(!###############################################!4T?! H#! K'8+.0%*!)(Q10.(['(+!1.!+>2.!-.!%7,%&'()*'+),(!#################################################################!4TU! !"#$%&'()@)))#CC62/,<2013),5B)C,<2.1<3);C26.C<2L5.37)7777777777777777777777777777777777777777777)*AA! #1,6M3.)C-;N0C;-,<02-.).<);<5>.)>.)/,3)601?2<5>21,6.)777777777777777777777777777777777777777777)*AA! !"! !*3&)0.(3+)*%""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%4BO! !!"! P>,./3$3,%"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%4B;! "#! P.8%0+.3.(+!######################################################################################################################!4IL! 5#! P7*01+'+*!)(-)/)-0.1*!2%7,27%'+,)%.*!#################################################################################!4IL! ;#! J+0-.!1,(&)+0-)('1.!2%7!.+!2,*+!,27%'+,)%.!86.\!0(.!2'+).(+.!#########################################!4IG! !!!"! G+,(.,,+)*%"""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""""%4OA! "#! H)*80**),(!*0%!1.*!%7*01+'+*!-0!&%,02.!-.!2'+).(+*!2%7N,27%'+,)%.!###################################!4:?! 5#! H)*80**),(!*0%!1<7+0-.!1,(&)+0-)('1.!##################################################################################!4:A! ;#! ;,(810*),(!*0%!1'!%7,%&'()*'+),(!#######################################################################################!4LM! !"#$%&'()O)))))))))!01/653201)?;1;-,6.)777777777777777777777777777777777777777777777777777777777777777777)*J9!
6
INTRODUCTION
GENERALE
« Si le cerveau était assez simple pour être compris, Nous serions trop bêtes pour le comprendre » L. Watson « C’est le langage qui créé l’homme. » J. d’Ormesson « La lecture est à l’esprit ce que l’exercice est au corps. » J. Addison
L’écriture est un système de représentation graphique permettant de transmettre de l’information sans le support de la voix. L’importance de cette invention a fait d’elle le marqueur entre la préhistoire et l’histoire. Les connaissances humaines pouvant alors être transmises de génération en génération sans déformation.
De très nombreuses théories furent développées pour expliquer cette faculté humaine, mais il fallut attendre le 19ème siècle pour que cette faculté soit définitivement rattachée au cerveau plutôt qu’au divin. La première observation reliant le langage et le cerveau fut montrée par Paul Broca en 1861, puis lui succédèrent d’autres médecins comme Carl Wernicke (1848 - 1905) ou Joseph Jules Déjerine (1849 - 1919). Tous montrèrent qu’une lésion d’une région ciblée du cortex conduisait à des troubles spécifiques langagiers, de production, de compréhension ou encore de lecture, les neurosciences modernes étaient nées.
La présence de ces régions langagières spécifiques à différentes opérations linguistiques conduit rapidement à l’hypothèse d’un réseau de régions corticales interconnectées et dont l’interaction dynamique produit le langage. Certaines de ces régions permettront la production, la compréhension de la parole ou encore la lecture ce qui sera le thème principal de cette thèse.
La lecture est intrinsèquement reliée à l’écriture. Or l’écriture n’est apparue il y a qu’environ 5000 ans, ce qui ne représente qu’une fraction de seconde sur l’échelle de l’évolution biologique. Cette observation suppose donc que le substrat neuronal sur lequel repose la lecture était préexistant. En particulier, on peut supposer que les structures corticales permettant la reconnaissance des objets interviennent dans celle des mots écrits. Cependant, les lettres et les mots ne sont pas tout à fait des objets comme les autres. En effet, leur reconnaissance complète nécessite trois opérations cognitives. Tout d’abord, la reconnaissance orthographique (i.e., le décodage des lettres) suivis du décodage phonologique (i.e., l’association à leur son) et de la récupération sémantique (i.e., l’accès à leur sens).
7 Il a longtemps été considéré que ces trois étapes se faisaient de façon sérielle progressive, les unes après les autres. Cependant, de l’expérience de tout normo-lecteur expérimenté et de récentes études, il semble qu’il existe une voie directe rapide de l’accès au sens des mots uniquement à partir de leur forme orthographique ou lexicale sans avoir besoin d’effectuer un décodage phonologique. Au cours des dernières décennies, différentes études ont montré seulement de façon indirecte ou théorique cette voie directe d’accès lexico-sémantique.
Depuis l’émergence des moyens de neuroimagerie moderne, de très nombreuses études se sont intéressées à la localisation de ces différentes aires fonctionnelles de la reconnaissance visuelle des mots, notamment en Imagerie par Résonnance Magnétique fonctionnelle (IRMf) ou en Tomographie par Emission de positons (TEP). Cependant, bien qu’ayant une très bonne résolution spatiale, ces techniques ne peuvent pas mettre en évidence la dynamique temporelle de l’intégration des différentes opérations cognitives nécessaire au décodage du mot. A l’inverse, l’électroencéphalographie (EEG) ou la magnétoencéphalographie (MEG) possèdent une résolution temporelle excellente de l’ordre de la milliseconde. Cependant la localisation spatiale des régions corticales ayant générée l’activité électrique enregistrée au niveau du scalp est restée pendant très longtemps un obstacle considérable à la mise en évidence du réseau fonctionnel sous tendant la lecture.
Malgré un très grand nombre d’études sur le sujet, il n’existe aujourd’hui que peu de consensus au sein de la communauté scientifique sur ce réseau en raison de nombreux résultats contradictoires et de la complexité du problème à résoudre. En effet, la compréhension du système de lecture nécessite la connaissance non seulement du « où ? » et du « quand ? » sont intégrés les différents processus, mais aussi du « comment ?», c'est-à-dire quel est le lien fonctionnel entre ces différentes régions.
Bien que récente, cette conception des fonctions cognitives sous forme de réseau d’interaction dynamique entre régions fonctionnelles semble faire aujourd’hui l’unanimité. Elle touche son paroxysme avec la conception hodotopique qui tend à montrer que la fonction d’une région est plus dépendante de ses connexions avec les autres régions fonctionnelles que de sa localisation corticale.
8 Mais que se passe-t-il si l’une des régions fonctionnelles du réseau langagier est endommagée ? Ceci peut être le cas lors de la survenue d’une lésion cérébrale, qu’elle soit transitoire (tels que un accident vasculaire cérébral ou un traumatisme crânien) ou chronique (tels que l’épilepsie). Grâce à son extraordinaire plasticité, le cerveau est capable de se réorganiser, de « déplacer » la région fonctionnelle vers une région saine au niveau du même hémisphère cérébrale (ipsilatérale) ou de l’autre hémisphère (controlatérale). Cependant, les mécanismes de cette réorganisation restent encore aujourd’hui très mal compris ainsi que les facteurs qui vont l’influencer.
L’objectif de cette thèse est double. Tout d’abord, il sera de mettre en évidence le réseau cortical de la reconnaissance visuelle des mots, en mettant en exergue les différents processus cognitifs et leurs substrats neuroanatomiques. Le but sera, non seulement de mettre en évidence où et quand ces processus sont intégrés, mais aussi de mettre en évidence les mécanismes d’intégration au sein de ce réseau, c'est-à-dire de révéler la connectivité et l’interaction entre les différentes régions fonctionnelles lors de la reconnaissance visuelle des mots. Le second objectif sera d’observer les modifications au niveau de ce réseau dans le cas où une région fonctionnelle est lésée et en particulier dans le cas de l’épilepsie focale temporale avant et après opération chirurgicale.
La première partie de cette thèse sera consacrée à l’état de l’art sur la reconnaissance visuelle, en premier lieu sur la reconnaissance visuelle d’objets, mettant en évidence le substrat neuronal initial sur lequel repose la lecture. Dans un deuxième temps, nous verrons l’état de l’art de la reconnaissance visuelle des mots et des trois étapes fondamentales à leurs décodages. La dernière partie sera consacrée à la présentation des derniers avancements sur la modélisation de la reconnaissance visuelle et du réseau permettant l’intégration fonctionnelle.
L’observation de la dynamique temporelle de ce réseau nécessite l’utilisation de l’électro-encéphalographie. La localisation spatiale des régions intégratives par EEG est restée pendant de nombreuses années un problème inextricable. Cependant, depuis une vingtaine d’années, d’énormes progrès ont été effectués dans le développement d’algorithmes permettant la reconstruction au niveau cortical des sources qui ont généré les signaux recueillis sur le scalp. Ces aspects méthodologiques seront abordés dans le second chapitre de
9 cette thèse. De plus, la mise en évidence du réseau et de son interaction fonctionnelle nécessite l’utilisation de méthodologies avancées et en particulier l’utilisation de modèles causaux dynamiques permettant de révéler l’organisation de ce réseau.
Le troisième chapitre sera consacré aux résultats obtenus au cours de cette thèse en s’appuyant sur la littérature et la méthodologie décrite dans les chapitres précédents. Le quatrième chapitre présentera une discussion générale permettant d’intégrer ces résultats dans le contexte plus large de la reconnaissance visuelle et de proposer un modèle neuro-anatomique de la reconnaissance visuelle des mots.
Le cinquième chapitre sera consacré à l’étude de ce réseau chez les patients épileptiques ayant un foyer épileptique au niveau du lobe temporal gauche qui nous le verrons occupe une place prépondérante dans le traitement des mots écrits. Il s’ouvrira sur une introduction générale sur l’épilepsie, puis sur les mécanismes de plasticité qui permettent une réorganisation des régions fonctionnelles
Le sixième chapitre présentera les résultats obtenus chez des patients épileptiques en bilan chirurgical pour montrer l’effet de la présence d’une lésion au niveau d’une région fonctionnelle. Puis nous verrons le cas d’une patiente suivie de façon longitudinale pré et post opératoire pour observer la réorganisation induite pas la chirurgie curative.
Le septième et dernier chapitre fera une synthèse générale de ce travail de thèse, sur l’organisation du réseau de la lecture chez les sujets sains et les patients épileptiques et permettra de mettre en avant les perspectives qui découlent de ce travail de thèse.
10
CHAPITRE 1
La reconnaissance visuelle
11
I.
La reconnaissance visuelle des objets
A. Organisation des aires visuelles primaires
a. V1
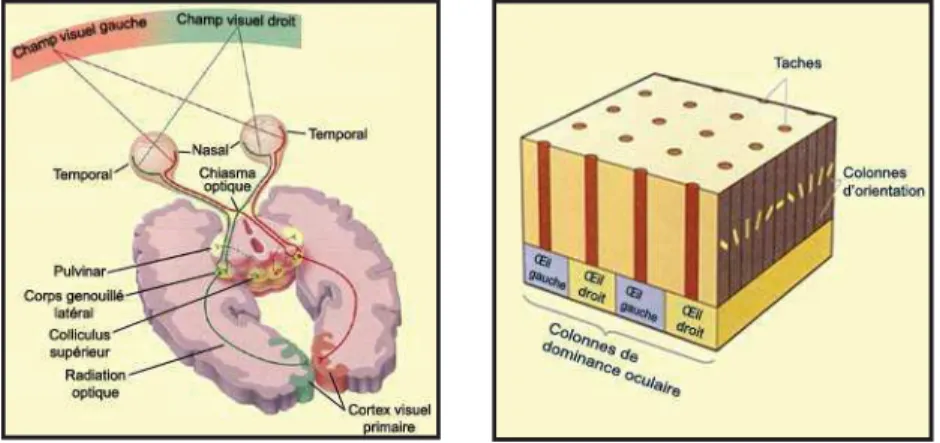
A la présentation d’un stimulus visuel, les informations provenant de la rétine vont être majoritairement envoyées, via le corps genouillé latéral au niveau du cortex occipital sur le cortex visuel primaire : V1. L'architecture fonctionnelle de V1 est rétinotopique1 (Hubel and Wiesel 1968). V1 se caractérise par sa structure striée en colonne où l’on distingue des colonnes de dominance oculaire et des colonnes d’orientation. Les colonnes de dominance oculaire représentent des bandes larges de tissu cérébral où se projettent alternativement l’œil gauche et l’œil droit. Les colonnes d’orientation représentent des bandes fines de tissu cérébral à l’intérieur desquelles les neurones sont sélectifs à la même orientation et à leur position dans le champ visuel définissant leur champ récepteur.
Figure 1: A gauche, schéma de description du système visuel de la rétine aux différentes structures sous-corticales et sous-corticales. A droite, représentation schématique de l’organisation rétinotopique de V1.
En résumé, l’analyse au niveau du cortex visuel primaire V1 est très basique. V1 extrait les traits qui composent une image en fonction de leur position et de leur orientation (Hubel and Wiesel 1977). 2
1
En d'autres termes, le champ visuel se projette point par point sur le cortex visuel primaire. Cette représentation, dite rétinotopique, permet à V1 de conserver une topographie du champ visuel sur la rétine.
2
Nous verrons par la suite que le traitement au niveau de V1 n’est pas uniquement bottom-up mais est aussi soumis à de forte influence top-down.
12 b. Les aires visuelles extrastriées
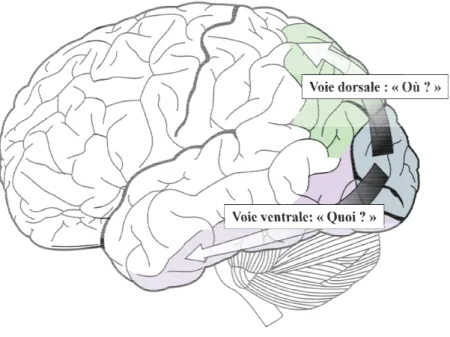
A partir de V1, l’information visuelle est projetée vers les aires visuelles extrastriées (e.g. V2, V3, V4 et V5). Au fur et à mesure, les champs récepteurs (portions du champ visuel) s’élargissent en combinant les traits des étages inférieurs rendant la rétinotopie de plus en plus grossière. Ainsi, au sein du cortex visuel, la représentation de l’image va devenir de plus en plus complexe et globale. En d’autres termes, le cerveau « éclate » l’image perçue en composantes élémentaires pour par la suite la recomposer progressivement dans les étages supérieurs du cortex visuel. On distingue alors classiquement deux grandes voies du traitement visuel: la voie dorsale et la voie ventrale (Figure 2).
c. Voie dorsale et voie ventrale
i. La voie dorsale
La voie dorsale, classiquement appelée la voie du « où ?» ou « pariétale » du fait de sa projection dans le cortex pariétal postérieur, est principalement impliquée dans la perception du mouvement et dans la perception pour l'action (Mishkin, Ungerleider et al. 1983). Au sein de cette voie, l’information visuelle chemine successivement à travers les aires V1, V2, V3, V3A et V5 et atteint les aires du cortex pariétal postérieur. Cette voie est quasi-exclusivement constituée des projections des cellules magnocellulaires. Ces cellules ont la propriété d’une part d’être sensible aux basses fréquences spatiales et d’autre part de répondre de manière très rapide lors d’une stimulation visuelle (Goodale and Milner 1992). Ainsi, le traitement de l'information dans la voie dorsale est extrêmement rapide. Nous verrons dans la discussion de cette thèse le rôle potentiel de cette voie dans la perception des mots et en particulier de son rôle dans l’attention spatiale.
ii. La voie ventrale
La voie ventrale, classiquement appelée la voie du « quoi ? » ou « temporale » du fait de sa projection dans le cortex inférotemporal, est principalement impliquée dans la perception de la forme et des couleurs (Mishkin, Ungerleider et al. 1983). Au sein de cette voie, l’information visuelle chemine successivement à travers les aires V1, V2, V4, et atteint le cortex inférotemporal (aire TE). Tout au long de ce cheminement le long de la voie ventrale, les champs récepteurs des neurones vont augmenter progressivement, combinant au fur et à mesure les éléments identifiés dans V1 pour former des représentations de plus en plus complexes. Les neurones de V1 répondront à de petites barres orientées, les neurones de
V2 combineront ces barres en formes plus complexes de suite (Tamura and Tanaka 2001
sur cette voie du traitement de l’information
Figure 2 : Schéma classique des deux voies de reconnaissance ventrale du « quoi ?».
V2 combineront ces barres en formes plus complexes comme des « T » ou des « Tamura and Tanaka 2001). Nous allons, dans la suite de cette section, nous sur cette voie du traitement de l’information visuelle et sur son fonctionnemen
: Schéma classique des deux voies de reconnaissance visuelle avec la voie dorsale du «
13 » ou des « Y » et ainsi . Nous allons, dans la suite de cette section, nous centrer
sur son fonctionnement.
B. Organisation hiérarchique de la voie ventrale
La reconnaissance visuelle des o hiérarchique à l’image d’une pyramide (Figure 3). L pyramide au niveau de V1 et se propage le long de l
(Figure 2). Ce cheminement s’accompagne de deux propriétés essentielles à la r visuelle des objets : l’accroissement de la complexité représentationne perceptive.
a. Du simple au complexe
A chaque étage de la pyramide, les réponses des neu intégrés en des représentations de plus en plus com s’accroissent d’un facteur 2 à 3 par étape pour arr
champ visuel. De façon intéressante, cette organisa opérer à tous les niveaux de complexité de façon bi uniquement ascendante mais aussi descendante. Ainsi le cerveau va pouvoir recréer l’information manq
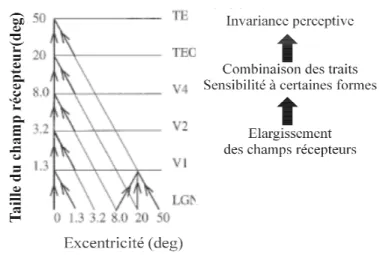
Figure 3 : Schéma de l’organisation pyramidale hiérarchique champ récepteur des neurones des différentes région
du stimulus. Au fur et à mesure du cheminement de l accroissent leur champ récepteur et deviennent de p Geniculate Nucleus, corps genouillé latéral (d’après Rolls, 2000).
Organisation hiérarchique de la voie ventrale
La reconnaissance visuelle des objets repose sur un principe d’organisation hiérarchique à l’image d’une pyramide (Figure 3). L’information visuelle entre à la base de la pyramide au niveau de V1 et se propage le long de la voie ventrale jusqu’au pôle temporal
s’accompagne de deux propriétés essentielles à la r : l’accroissement de la complexité représentationne
Du simple au complexe
A chaque étage de la pyramide, les réponses des neurones inférieurs sont combinées et intégrés en des représentations de plus en plus complexes. Les champs récepteurs s’accroissent d’un facteur 2 à 3 par étape pour arriver à des champs récepteurs de l’ordre du champ visuel. De façon intéressante, cette organisation hiérarchique combinatoire semble opérer à tous les niveaux de complexité de façon bidirectionnelle, c'est
uniquement ascendante mais aussi descendante. Ainsi, avec seulement un morceau du puzzle, le cerveau va pouvoir recréer l’information manquante en réactivant les niveaux inférieurs.
: Schéma de l’organisation pyramidale hiérarchique neuronale le long de la voie ventrale. Taille du champ récepteur des neurones des différentes régions corticales de la voie ventrale en fonction de l’excentricité du stimulus. Au fur et à mesure du cheminement de l’information dans la pyramide hiérarchique, les neu accroissent leur champ récepteur et deviennent de plus en plus indépendant de la position spatiale. LG
Geniculate Nucleus, corps genouillé latéral ; TEO : jonction occipito-temporale; TE : cortex temporal inférieur
14 bjets repose sur un principe d’organisation
’information visuelle entre à la base de la a voie ventrale jusqu’au pôle temporal s’accompagne de deux propriétés essentielles à la reconnaissance : l’accroissement de la complexité représentationnelle et l’invariance
rs sont combinées et plexes. Les champs récepteurs iver à des champs récepteurs de l’ordre du rarchique combinatoire semble directionnelle, c'est-à-dire non pas , avec seulement un morceau du puzzle, uante en réactivant les niveaux inférieurs.
neuronale le long de la voie ventrale. Taille du ventrale en fonction de l’excentricité ’information dans la pyramide hiérarchique, les neurones lus en plus indépendant de la position spatiale. LGN : Lateral : cortex temporal inférieur
b. Invariance perceptive
Au fur et à mesure de l’élévation dans la hiérarchi moins en moins sensibles à la taille, à la position, à l’illu formes qu’ils codent.
A la suite des travaux d’Hubert et Wiesel (1968) su mettant en évidence la sensibilité de ces neurones
comme Robert Desimone, Charles Gross, Keiji Tanaka long de la voie ventrale visuelle. L’une des expéri controversées fut effectuée par Tanaka et ses colla extrême sélectivité des neurones situés au niveau d présenté, le neurone choisi répond uniquement la fo sélectivité est d’autant plus remarquable qu
niveau, le neurone va préférer une forme précise, m position change de plusieurs degrés ou encore que l complètement différentes (Sary, 1993
Figure 4: enregistrement des neurones
ces neurones à des formes complexes, seule la vue d Tanaka, 2001).
Aussi étonnant que cela puisse paraitre, cette séle l’invariance perceptive pure pour atteindre un nive
En effet, en 2005, Quiroga et ses collaborateurs en inférotemporaux antérieur chez un patient épileptiq
Invariance perceptive
Au fur et à mesure de l’élévation dans la hiérarchie, les neurones vont devenir de oins sensibles à la taille, à la position, à l’illumination, à la déformation des
A la suite des travaux d’Hubert et Wiesel (1968) sur les aires visuelles du chat, mettant en évidence la sensibilité de ces neurones aux traits élémentaires, des chercheurs comme Robert Desimone, Charles Gross, Keiji Tanaka s’intéressèrent aux neurones en aval le long de la voie ventrale visuelle. L’une des expériences les plus marquantes et des plus controversées fut effectuée par Tanaka et ses collaborateurs en 2001. Ils montrèrent une extrême sélectivité des neurones situés au niveau du cortex temporal inférieur. Dans le cas présenté, le neurone choisi répond uniquement la forme d’une chaise (Figure 4). Cette sélectivité est d’autant plus remarquable qu’elle s’accompagne d’une forte invariance. A ce niveau, le neurone va préférer une forme précise, même si la taille de celle
position change de plusieurs degrés ou encore que la luminosité change créant des ombres s (Sary, 1993 ; Ito, 1995 ; Vogels, 2002).
: enregistrement des neurones inférotemporal du macaque. Ceci présente une remarquable sélectivi ces neurones à des formes complexes, seule la vue d’une chaise active fortement ce neurone (d’après T
Aussi étonnant que cela puisse paraitre, cette sélectivité semble même dépasser l’invariance perceptive pure pour atteindre un niveau de représentation beaucoup plus abstrait. En effet, en 2005, Quiroga et ses collaborateurs enregistrèrent l’activité de neurones inférotemporaux antérieur chez un patient épileptique, ils trouvèrent un neurone répondant
15 e, les neurones vont devenir de
mination, à la déformation des
r les aires visuelles du chat, ires, des chercheurs s’intéressèrent aux neurones en aval le ences les plus marquantes et des plus teurs en 2001. Ils montrèrent une u cortex temporal inférieur. Dans le cas rme d’une chaise (Figure 4). Cette ’elle s’accompagne d’une forte invariance. A ce ême si la taille de celle-ci varie ou que sa a luminosité change créant des ombres
du macaque. Ceci présente une remarquable sélectivité de ’une chaise active fortement ce neurone (d’après Tamura et
ctivité semble même dépasser au de représentation beaucoup plus abstrait. trèrent l’activité de neurones ue, ils trouvèrent un neurone répondant
16 spécifiquement à Jennifer Aniston ! Le plus étonnant est que ce neurone déchargeait qu’on lui montre Jennifer Aniston en portrait, en buste ou en pied, mais aussi en caricature ou en lui présentant simplement son nom écrit! (Quiroga, 2005).
Ce résultat ne tend pas à montrer que tout Jennifer Aniston est contenue dans un neurone, mais souligne l’importance de la pyramide hiérarchique et le réseau associé à la reconnaissance de cette personne. De plus il montre la bidirectionnalité de cette pyramide permettant ainsi de retrouver notamment le nom de la personne à partir de son visage et vice et versa ou même à partir d’indice partiel (tel que simplement ses yeux).
C. Construction par l’expérience de la pyramide hiérarchique
Le cortex temporal inférieur possède de fortes capacités de plasticité et d’apprentissage. Cette structure hiérarchique pyramidale semble donc se forger par l’expérience et l’apprentissage, et donc avec une interaction forte avec le monde extérieur. L’organisation de cette pyramide est permise par la cooccurrence fréquente de différents traits saillants ou à une échelle au-dessus de formes. Cependant, ces formes peuvent être complètement arbitraires et artificielles, seules leur cooccurrence et la fréquence d’apparition semblent jouer un rôle dans l’apprentissage des formes. Baker et ses collaborateurs ont appris à des singes à reconnaitre des formes étranges composés d’un segment avec deux formes particulières à chacun des bouts. Après apprentissage, certains neurones du cortex temporal inferieur répondaient spécifiquement à la combinaison de ces formes. Leurs réponses au groupement de ces deux formes dépassaient largement la somme des réponses aux formes isolées. Ces neurones ont appris la régularité de la conjonction de ces deux formes montrant ainsi les capacités de plasticité et d’apprentissage du cortex temporal inférieur (Baker, 2002).
Cette capacité d’apprentissage semble être permise par les conjonctions non seulement spatiales mais aussi temporelles. Dans une expérience de Miyashita et collaborateurs, des fractales n’ayant aucun rapport entre elles étaient présentées. S’il n’existait pas de redondance dans la présentation, chacune des fractales était codées séparément uniquement sur leurs traits visuels. Cependant, s’il existait une régularité dans l’ordre de présentation, une fractale A toujours suivie d’une fractale B, il existait des neurones qui répondaient aux deux fractales dont le seul point commun ait été d’avoir été associé dans le temps (Miyashita, 1991).
Cette double capacité d’apprentissage de régularité de cooccurrence à la fois spatiale et temporelle semble permettre la création de représentation invariante avec un très haut niveau d’abstraction.
17
II.
La reconnaissance visuelle des mots
A. Introduction
L’homme a inventé l’écriture et donc a fortiori la lecture il y a environ 5000 ans, ce qui ne représente qu’une fraction de seconde sur l’échelle de l’évolution génétique. Ainsi, le substrat neuronal sur lequel repose la lecture doit s’effectuer à partir de structures cérébrales préexistantes et notamment celle impliquées dans la reconnaissance visuelle des objets introduites précédemment. Dans cette perspective, les lettres pourraient être associées à de petits objets composées de trait élémentaires, dont la combinaison forme des graphèmes, dont l’association formerait des mots puis des phrases, puis des textes, etc.
Une telle vision refléterait ainsi ce que nous avons vu précédemment : l’augmentation de complexité de la représentation selon la pyramide hiérarchique temporale. Grâce à l’apprentissage, nous sommes devenus des experts de la reconnaissance de ces objets ainsi que de leur combinaison, pour rendre automatique l’ensemble des processus de lecture.
Dans un premier temps, nous nous intéresserons à la reconnaissance visuelle des lettres, composant élémentaire des mots. Nous nous intéresserons par la suite aux mots écrits à travers les trois étapes nécessaires à leur décodage : l’orthographe, la phonologie et la sémantique. Nous nous intéresserons au substrat neuro-anatomique sous tendant ces différentes opérations cognitives puis nous nous intéresserons au réseau constitué par ces différentes régions fonctionnelles.
B. La reconnaissance visuelle des lettres : de l’objet à la lettre
Si l’homme n’a pas évolué biologiquement pour l’écriture et la lecture, alors ce sont les lettres qui ont été formées pour être comprise facilement par l’homme (Changizi, Zhang et al. 2006). Dans presque tous les systèmes d’écritures, le nombre de traits pour former une lettre est d’environ 3 (Changizi and Shimojo 2005). Ce faible nombre de traits permet non seulement une écriture simple et rapide mais aussi un décodage simple et rapide de leur forme.
Comme nous l’avons vu précédemment, le système visuel est organisé de façon hiérarchique : le cortex visuel primaire (V1) ne distingue que les orientations des traits, puis ceux-ci sont combinés le long de la voie ventrale pour créer des représentations de plus en plus complexes. Ainsi, les trois traits composants les lettres pourraient ainsi être rapidement assemblés et identifiés au niveau de la pyramide hiérarchique neuronale du système visuel.
Dans cette idée, Dehaene et ses collaborateurs prop Detectors) qui tente d’attribuer aux différentes ré capacités d’intégration selon cette hiérarchie croissante d’un neurone visuel à leur combinaison sous forme d de bigrammes puis de mots (Dehaene, Cohen et al. 2005
Figure 4 : Modèle LCD (Local Combination Detectors) montrant neuronale d’un simple champ récepteur d’un neurone visuel, bigramme (Dehaene, 2005).
Dans cette idée, Dehaene et ses collaborateurs proposent le modèle LCD (Local Combination Detectors) qui tente d’attribuer aux différentes régions de la voie occipito
tés d’intégration selon cette hiérarchie croissante de complexité, du champ récepteur d’un neurone visuel à leur combinaison sous forme de lettres qui se combine alors sous forme
Dehaene, Cohen et al. 2005).
: Modèle LCD (Local Combination Detectors) montrant la pyramide hiérarchique d’intégration le d’un simple champ récepteur d’un neurone visuel, à sa combinaison sous forme de lettres puis de
18 osent le modèle LCD (Local Combination gions de la voie occipito-temporale leurs de complexité, du champ récepteur e lettres qui se combine alors sous forme
la pyramide hiérarchique d’intégration à sa combinaison sous forme de lettres puis de
19
C. Le traitement orthographique
a. Approche psycholinguistique
i. De la régularité orthographique
Quelque soit la langue ou l’écriture, il existe certaines lettres qui reviennent plus souvent dans les mots que d’autres, c’est ce qu’on appelle la « fréquence graphémique ». Par exemple, le « e » en français est très courant le « w » beaucoup moins (pour avoir idée des fréquences, voir le nombre de points que vaut une lettre au scrabble !). De la même manière, certaines combinaisons de lettres, sous forme de bigrammes ou trigrammes sont beaucoup plus récurrentes que d’autres (« le » vs « rk » ou « ion » vs « obn »). On parlera alors de « régularité orthographique ».
Or, le cerveau semble être capable d’extraire la régularité de l’orthographe dans une langue donnée et notamment la redondance des chaines de lettres. L’expérience devenu classique de Miller et al. (Miller, 1954) met en évidence l’utilisation par le sujet des redondances statistiques des chaines de lettres en anglais. Ils présentèrent aux sujets des chaines de 8 lettres dont les régularités orthographiques variaient d’une condition à l’autre et faisaient varier aussi le temps de présentation. La tache demandée aux sujets était de reporter le plus rapidement possible les lettres vues. Les auteurs ont créé quatre « ordres » (ou conditions expérimentales) dans lesquelles les relations entre les lettres variaient (voir Figure 5).
A l’ordre 0, les lettres étaient tirées au hasard parmi les 26 lettres de l’alphabet. A l’ordre 1, les lettres respectaient la distribution statistique (fréquence graphémique) de la langue anglaise indépendamment les unes des autres. A l’ordre 2, le tirage de la lettre n+1 dépendait de la lettre n suivant la distribution statistique de l’anglais (régularité orthographique). A l’ordre 4 étaient prises en compte les 3 lettres précédentes. Les pseudo-mots créés ressemblaient ainsi de plus en plus à de vrais pseudo-mots.
Les résultats montrent classiquement une amélioration des performances avec le temps de présentation. Mais ils montrent aussi un effet très important de la régularité statistique des chaines de caractères. Dans les conditions d’ordre 4 ou d’ordre 2, les sujets ont des performances bien plus élevées que dans les deux autres conditions, ceci tend donc à montrer que les sujets ont utilisé la régularité orthographique de l’anglais, qui a été internalisée lors de l’apprentissage de la lecture. On peut voir aussi que cet effet ne se base pas essentiellement à
20 l’échelle de la lettre (ordre 1), mais à un niveau hiérarchique plus élevé dans le traitement de l’information, à l’échelle au moins du bigramme (ordre 2) et encore plus à l’échelle du trigramme/quadrigramme.
Ainsi la cooccurrence fréquente de lettres au cours de l’apprentissage permet l’émergence de leurs représentations commune au sein du cortex temporal inférieur3. Autrement dit, on voit ici émerger l’effet de la pyramide hiérarchique observée sur la reconnaissance visuelle des objets, où les étages supérieurs ont une influence descendante sur le traitement des niveaux inférieurs.
Figure 5 : A gauche, liste des chaines de lettres aux différentes propriétés de régularité orthographique. A droite, taux de bonnes réponses en fonction de la durée de présentation des différentes catégories de chaines de caractères (tirée de Miller, 1954).
ii. L’effet de supériorité lexicale
L’utilisation automatique des régularités orthographiques dans une langue donnée par un normo-lecteur a été validée par l’expérience de Reicher et de ses collaborateurs en 1969. Dans cette expérience, il était demandé aux participants de reporter une lettre qui était très brièvement présentée (à la limite de la perception consciente entre 35 et 85 ms) puis masquée (#). Cette lettre était soit présentée dans un mot, soit de façon isolée, soit dans un non-mot construit comme une anagramme de mot existant sans respecter les règles orthographiques.
3
21 Ils observèrent que les performances de report des participants étaient bien meilleures dans le cas où celle-ci était présentée dans le contexte d’un mot par rapport aux deux autres conditions. Ceci montre que la capacité du sujet à extraire de l’information sur la lettre est augmentée. En effet, dans l’exemple ci-dessous (Figure 6), l’information portée par les lettres WOR est nulle étant donné qu’il pourrait s’agir du mot WORK, mais en présence de ces lettres le cerveau du sujet est capable d’extraire plus d’informations.4
A la suite des expériences de Miller (1954), cette expérience met en exergue l’organisation hiérarchique bidirectionnelle de la reconnaissance visuelle des mots. Plus la régularité orthographique va augmenter en atteignant son paroxysme dans le cas des mots, plus le sujet sera capable de reporter de lettres ou de les reconnaitre à un seuil proche de l’invisible. Ceci tend à montrer l’importance et l’influence des informations descendantes provenant de régions de hauts niveaux sur les régions perceptives de bas niveaux.
Figure 6 : Paradigme de l’expérience de Reicher (1969) montrant la supériorité de l’effet lexical. La détection d’une lettre au seuil de perceptibilité sera augmentée par son appartenance à un mot par rapport à une condition isolée ou dans une chaine de caractères ne respectant pas la régularité orthographique.
4
Il est intéressant de noter que cet effet s’étend à d’autres conditions de façon graduée, la performance des sujets s’améliore aussi si la lettre est présentée dans le cas d’un pseudo-mots ou même d’une chaine de caractères à condition que ceux-ci respectent la régularité orthographique et les statistiques de la langue.
22 b. Représentation neuro-anatomique de l’orthographe
En Octobre 1887, Monsieur C., négociant retraité, se rendit soudainement compte qu’il ne parvenait plus à lire un seul mot de son journal. Les mots et les lettres étaient devenus des symboles ayant perdu tous leurs sens. Cependant, Monsieur C. pouvait parler, distinguer avec netteté les objets ou les gens et même écrire. Voici, le premier cas rapporté par Déjerine (1892) d’un trouble sélectif de la lecture : l’alexie pure. Cette observation neuropsychologique permit de mettre en évidence l’existence d’une région cérébrale capable de reconnaitre les lettres détruites chez Monsieur C.. En effet, à son décès, le docteur Déjerine observa lors de l’autopsie, que le patient avait une lésion au niveau de l’hémisphère gauche dans sa partie postérieure, plus spécifiquement dans une région située entre le lobe occipital et le lobe temporal inférieur (voir Figure 7). Déjerine mit en avant le principe de déconnexion entre régions cérébrales et supposera alors, étant donné que le patient pouvant toujours écrire, que « le centre visuel des lettres » se situait au niveau du gyrus angulaire et que c’est la déconnexion entre cette région et les régions visuelles qui avait conduit à l’alexie. Bien qu’aujourd’hui on sache que la localisation du « centre visuel des lettres » était erronée, son intuition sur le lien entre déficit et déconnexion entre aires cérébrale n’a jamais été autant d’actualité.
Figure 7 : A gauche, dessin anatomique de la lésion de Monsieur C. ayant provoqué l’alexie réalisé par Joseph-Jules Déjerine (1892), image du milieu. A droite, localisation de la « VWFA » par IRMf (Cohen, Dehaene et al. 2000).
23 Grâce au moyen d’imagerie moderne comme l’IRMf, il a été possible d’identifier la localisation de cette région sensible à la forme visuelle des lettres au niveau de la région occipito-temporale gauche (Figure 7). La spécialisation de cette région pour la reconnaissance des lettres et des mots lui fut conférée par Cohen et Dehaene qui la dénommèrent la Visual Word Form Area. (VWFA) (Cohen, Dehaene et al. 2000; Dehaene, Le Clec et al. 2002; Cohen and Dehaene 2004). Il a été possible de différencier plusieurs types d’alexie. Les lésions les plus postérieures de la VWFA induisent à une incapacité totale à reconnaitre les lettres, alors qu’une lésion plus antérieure peut préserver l’identification isolée des lettres mais pas des mots entiers. Les patients présentant ce dernier type de lésions sont capables d’accéder au sens de mots par l’intermédiaire de l’épellation séquentielle des lettres composant le mot (Cohen, Jobert et al. 2004; Dehaene, Cohen et al. 2005). Ceci tend donc à montrer qu’il existe au sein de cette VWFA différentes étapes de traitement de la lettre au mot rappelant implicitement l’organisation croissante de la reconnaissance visuelle des objets.
L’un des moyens de mettre en évidence les capacités fonctionnelles d’une région est d’utiliser l’effet de facilitation. Lorsque l’on soumet deux fois de suite un sujet à un même stimulus, l’activation dans la région sensible à ce stimulus sera diminuée lors de la seconde présentation. Si les deux stimuli possèdent une propriété commune, alors on observera un effet de facilitation dans la région sensible à cette propriété commune. En général, on utilise des taches d’amorçage subliminal, c'est-à-dire que le premier stimulus est présenté en dessous du seuil de perception consciente. Le premier stimulus est appelé « l’amorce », le second « la cible »
En utilisant cette méthode, Dehaene et al. (2002) montrèrent un effet de facilitation au niveau des régions occipitales si l’amorce et la cible étaient les mêmes stimuli, à savoir des mots écrits dans la même casse5 (chat/chat). Par ailleurs, ils montrèrent un effet de facilitation au niveau de la VWFA, si l’amorce et la cible étaient les mêmes mots mais écrits dans des casses différentes (chat/CHAT).
Ils proposèrent ainsi l’existence d’une facilitation induit par la répétition physique au niveau des régions occipitales et une facilitation indépendante de la casse au niveau du gyrus fusiforme.
5
24 Figure 8: cartographie par IRMf des régions sensibles aux mots écrits dans une expérience d’amorçage subliminal et avec variation de la casse. A gauche, le gyrus occipital droit est activé pour les mots mais est sensible à la variation de la casse entre l’amorce et la cible. A droite, le gyrus temporal inférieur gauche est activé et n’est pas sensible à la variation de casse (Cohen, 2002)
Dans le cadre de l’organisation hiérarchique de la voie ventrale précédemment présentée, ce résultat montre donc que la partie postérieure du gyrus temporal inférieur fait déjà partie d’un niveau hiérarchique relativement élevé. En effet, non seulement cette région possède des propriétés d’invariance perceptive mais de plus son niveau d’abstraction est relativement élevé. A ce niveau de la pyramide, le système ne fait plus de distinction entre les minuscules et les majuscules, qui pourtant ne possèdent pour la plupart aucun trait physique commun. En effet, les correspondances entre les minuscules et les majuscules sont surtout d’ordre culturel. Ceci montre donc la nécessité de l’apprentissage de ces correspondances et qu’à ce niveau hiérarchique elles sont représentées par les mêmes assemblages de neurones.
Cependant, en raison des arguments évolutionnistes cités précédemment, la spécialisation d’une région chez l’espèce humaine pour la reconnaissance unique des lettres est peu probable. De plus, différentes équipes ont montré que cette région était activée par d’autres items que des lettres, dénonçant ainsi le mythe de la VWFA (Price and Devlin 2003; Kherif, Josse et al. 2010). Voir encadré « le babouin lecteur » en fin de chapitre.
25 c. Etudes électrophysiologiques de la représentation orthographique
Les études en IRMf ont permis d’identifier la localisation spatiale du traitement orthographique des mots et de mettre en évidence les différents niveaux hiérarchiques activés. Cependant, de part la faible résolution temporelle de l’IRMf, ils n’ont pu mettre en évidence le décours temporel de la cascade d’activation au sein de la voie ventrale. De nombreuses études ont été menées en vue d’établir cette chronologie en utilisant des méthodes électrophysiologiques, soit chez le sujet sain avec l’EEG ou la MEG, soit chez le patient épileptique à l’aide d’électrodes implantées le long de la voie ventrale.
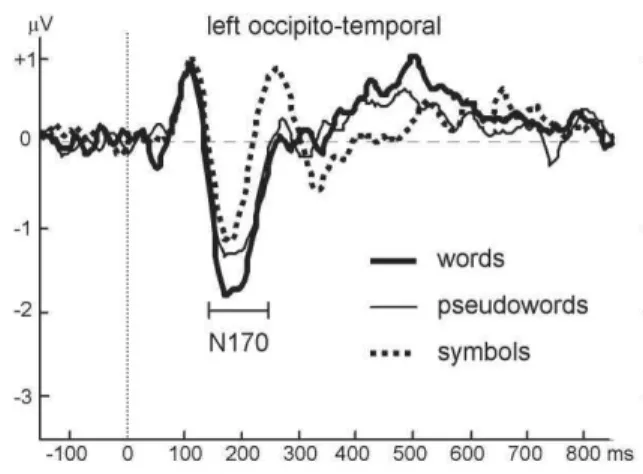
i. La N170
La N170 est une onde négative dont le maximum est généralement atteint au niveau de l’électrode PO7, c'est-à-dire au niveau de la région occipitale inférieure gauche. Bien que sa dénomination représente une onde négative ayant son maximum d’amplitude à 170 ms, il existe une forte variabilité sur la latence de cette onde dans la littérature d’électrophysiologie. En effet, certaines équipes trouvent cette onde à une latence de 120 ms (Compton, Grossenbacher et al. 1991; Spironelli and Angrilli 2007) et d’autres plus tardive jusqu’à 200 ms (Cohen, 2002). Des auteurs proposent ainsi que cette onde soit en réalité légèrement plus précoce et atteindrait son maximum vers 150 ms (Mondini, Angrilli et al. 2008; Dien 2009). Cette dénomination, légèrement confondante, a été choisie pour être mise en rapport avec la N170 générée par présentation de visages, onde beaucoup plus étudiée à l’époque (Bentin, Mouchetant-Rostaing et al. 1999). La latéralisation hémisphérique de la N170 est fonction du matériel présenté, en effet, elle est observée à droite pour les visages et à gauche pour le matériel verbal.
Concernant son rôle fonctionnel, la plupart des auteurs proposent que cette onde soit, lors du traitement verbal, le reflet d’une étape sub-lexicale. Cependant, ce rôle fonctionnel ne fait pas consensus en raison de résultats contradictoires entre différentes études.
La première équipe à mettre en évidence un lien entre la N170 et le traitement du matériel verbal fut celle de Compton (1991). Vers 125 ms, ils observèrent une augmentation de l’amplitude de cette onde pour des chaines de consonnes par rapport à des mots. Bien que ce résultat ait été reproduit quelques années plus tard (McCandliss, Posner et al. 1997), deux autres études ne trouvèrent pas de différence entre les mots, les pseudo-mots et les chaines de consonnes (Bentin, 1999 ; Simon, 2004). Cependant, ils montrèrent une augmentation de cette amplitude dans les trois cas cités précédemment par rapport à des formes, des symboles ou
26 des pseudo-lettres, confirmant ainsi le lien entre la N170 et le traitement des lettres. Par la suite, certains trouvèrent qu’il existait une différence d’amplitude de cette onde entre les mots et les pseudo-mots (mots > pseudo-mots) (Maurer, Brandeis et al. 2005) tandis que d’autres trouvèrent une différence d’amplitude dans le sens inverse (pseudo-mots > mots), bien que celle-ci ne soit pas significative (Hauk and Pulvermüller 2004)(Hauk, 2006). Les tentatives d’évaluation de l’effet de la familiarité sur l’amplitude de cette onde furent eux aussi très contradictoires malgré un contrôle de la longueur des mots et de la fréquence des bigrammes et des trigrammes (Hauk, 2004 ; Penolazzi, 2007 ; Hauk, 2006). De plus, Brem et ses collaborateurs ont montré que cette onde pouvait devenir semblable à celle provoquée par des mots sur des chaines de pseudo-caractères après un entrainement intensif (Brem, Lang-Dullenkopf et al. 2005), montrant ainsi l’effet de la familiarité sur cette onde lors du traitement d’un matériel verbal.
Figure 9 : Modulation de l’amplitude de la N170 enregistrée au niveau de PO7 en fonction du matériel verbal présenté. (d’après Maurer, 2005)
La divergence de ces résultats sur la N170, tant sur son amplitude que sur sa latence, peut être expliquée par différents facteurs. Notamment le matériel verbal utilisé (chaines de caractères, pseudo-mots, mots, etc.), les contrôles utilisés (pseudo-lettres, repos, etc.) mais également la tâche réalisée par les participants (lecture active/passive, catégorisation lexicale, etc.). Par ailleurs, il semble que la langue étudiée puisse jouer un rôle en fonction de sa transparence grapho-phonémique (cf, section II.C). Enfin, les divergences dans les résultats de la littérature peuvent venir de l’influence de processus top-down venant moduler ces activations. En effet, il a été montré que bien que relativement précoce la N170 était modulée par l’attention spatiale (McCarthy and Nobre 1993; Awh, Anllo-Vento et al. 2000) qui aurait
27 un rôle d’amplificateur de l’activité neuronale afin d’améliorer le « rapport signal/bruit » et ainsi extraire plus d’informations ou les informations les plus pertinentes pour la reconnaissance du stimulus (Hillyard, Vogel et al. 1998).
En conclusion, le lien entre la N170 et le traitement orthographique, en particulier une étape sub-lexicale, semblent établis. Cependant, de nombreuses ombres restent à éclaircir pour comprendre son rôle fonctionnel exact.
ii. La N250 ou le « recognition potential »
La N250 ou « recognition potential » (RP) est une onde négative avec une latence entre 200 et 250 ms évoquée lors de la présentation de mots. Elle fut découverte en 1990 par Rudell et ses collaborateurs (1990) et de nombreux travaux se sont ensuite intéressés à sa localisation et à son rôle fonctionnel. (Rudell and Hua 1996; Rudell and Hua 1997; Hinojosa, Martín-Loeches et al. 2000; Martín-Loeches, Hinojosa et al. 2001; Martín-Loeches, Sommer et al. 2005). Dans leur expérience princeps, Rudell et ses collaborateurs montrèrent une différence d’amplitude de cette onde chez des normo-lecteurs anglais lorsqu’on leur présentait des mots anglais par rapport à des mots chinois. Cette différence était inversée dans le cas de lecteurs chinois. Ils localisèrent cette onde au niveau occipital. Des études complémentaires ont pu montrer sa sensibilité à certains facteurs expérimentaux comme la dégradation des stimuli ou la difficulté des mots présentés (Rudell and Hua 1997; Rudell, Hu et al. 1999), tous deux augmentant la latence du RP, alors que la présentation d’une amorce cohérente diminue sa latence (Rudell and Hua 1996; Hinojosa, Martín-Loeches et al. 2004). L’amplitude du RP augmente avec le niveau linguistique des stimuli, en atteignant son maximum pour les mots (Martín-Loeches 2007). De plus, ils montrèrent que le RP était affecté par l’attention sélective et la tâche demandée (Rudell and Hua 1996).
Enfin, cette équipe a mis en évidence la corrélation entre la latence du RP et les capacités de lecture des sujets (Rudell and Hua 1997). De ces différents résultats, il fut stipulé que la N250 était un marqueur de la perception et de la reconnaissance des mots.
La localisation des générateurs de cette onde fut tout d’abord situé dans le cortex occipital et en particulier dans les aires extrastriées. Cependant, cette localisation a été très controversée et il semblerait aujourd’hui que cette onde puisse être associée plus au lobe temporal et en particulier à sa partie postérieure, nommée par Dien et ses collaborateurs la « Language formulation area » (Dien 2009). Cette localisation semble plus cohérente avec le principe d’augmentation de la représentation corticale des stimuli écrits le long de la voie
ventrale. Ainsi, suite à une étape sub
ms, N170), une seconde étape lexicale serait effectuée. Cette dernière en correspondance avec le lexique mental au niveau
vers 200/ 250 ms.
Figure 10: Modulation de l’amplitude de la
d. Approche computationnelle de la représentation or
Le modèle le plus influ
est celui de Rumelhart et McClelland
1986)(Rumelhart & McClelland, 1981, 1982). Ce modèle rep basé sur une organisation hiérarchique du traitemen
en 3 niveaux de hiérarchie distincte. Les connexions entre ces différent l’intérieur même de ces niveaux pourront être excit
11).
ventrale. Ainsi, suite à une étape sub-lexicale au niveau du gyrus fusiforme gauche (vers de étape lexicale serait effectuée. Cette dernière serait celle de la mise en correspondance avec le lexique mental au niveau du lobe temporal inférieur gauche et ce
: Modulation de l’amplitude de la N250 en fonction du matériel présenté (d’après
Approche computationnelle de la représentation or
« On ne peut lire si on ne connaît les lettres, seulement les lettres sont effacées par le sens.
Le modèle le plus influent dans la représentation de l’étape orthographiqu
rt et McClelland (Rumelhart 1977; Rumelhart and McClelland (Rumelhart & McClelland, 1981, 1982). Ce modèle représenté dans la Figure 11 est basé sur une organisation hiérarchique du traitement de l’information et peut être décomposé
rchie distincte. Les connexions entre ces différent l’intérieur même de ces niveaux pourront être excitatrices (->) ou inhibitrices (
28 lexicale au niveau du gyrus fusiforme gauche (vers 150 de étape lexicale serait effectuée. Cette dernière serait celle de la mise du lobe temporal inférieur gauche et ce
d’après Hinojosa, 2000)
Approche computationnelle de la représentation orthographique
On ne peut lire si on ne connaît les lettres, seulement les lettres sont effacées par le sens. » Alain
ent dans la représentation de l’étape orthographique et lexicale Rumelhart and McClelland résenté dans la Figure 11 est t de l’information et peut être décomposé rchie distincte. Les connexions entre ces différents niveaux ou à >) ou inhibitrices (-o) (voir Figure