Hématopoïèse et réponse immunitaire cellulaire chez Drosophila melanogaster
154
0
0
Texte intégral
Figure

+7
Outline
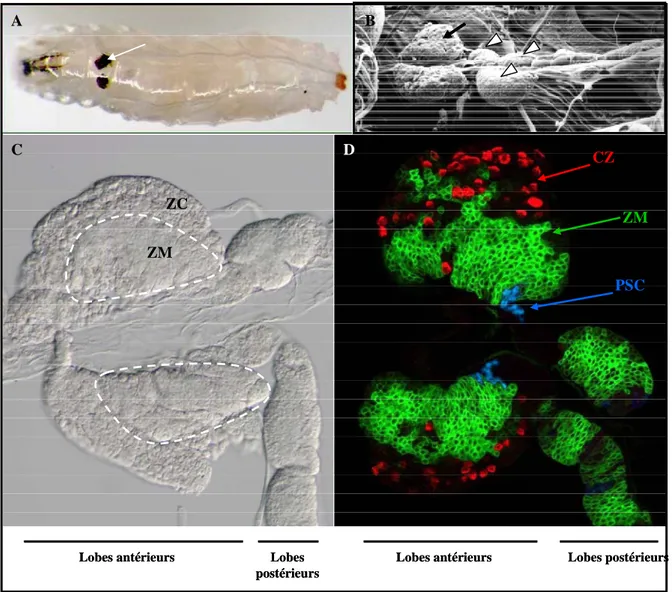
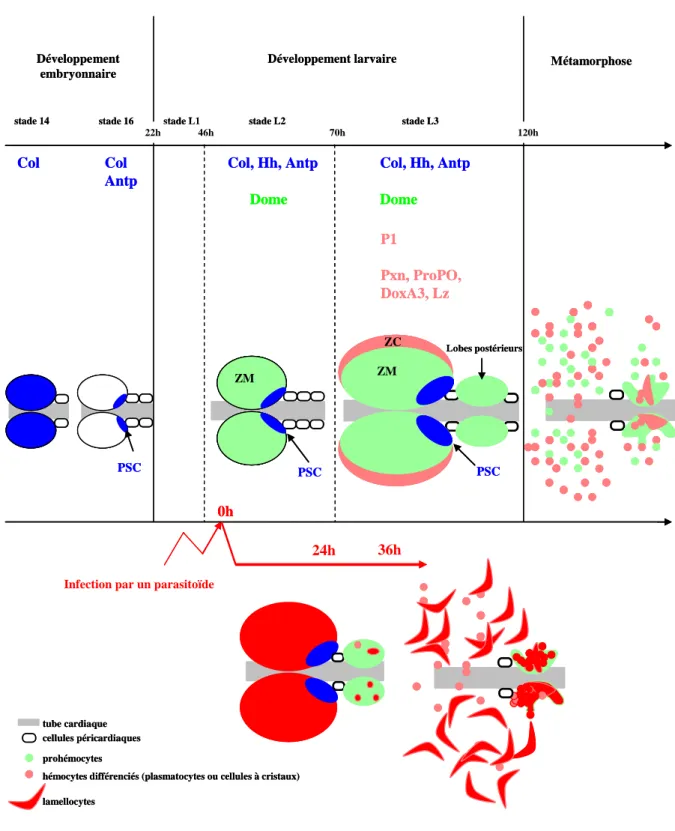
Localisation de la glande lymphatique dans la larve Vue dorsale d’une larve au troisième stade Les lobes antérieurs
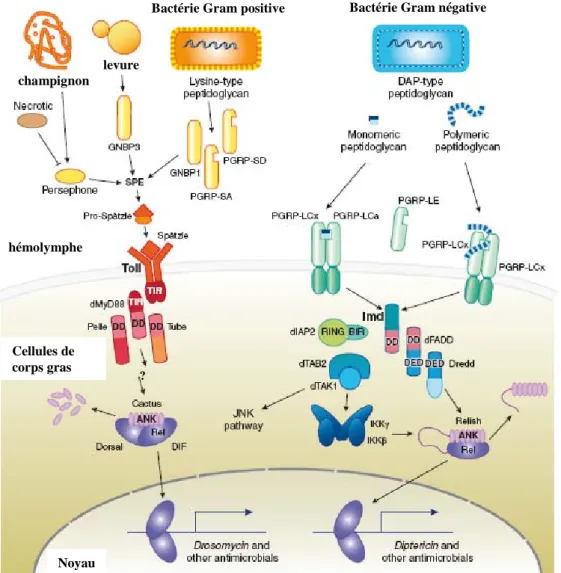
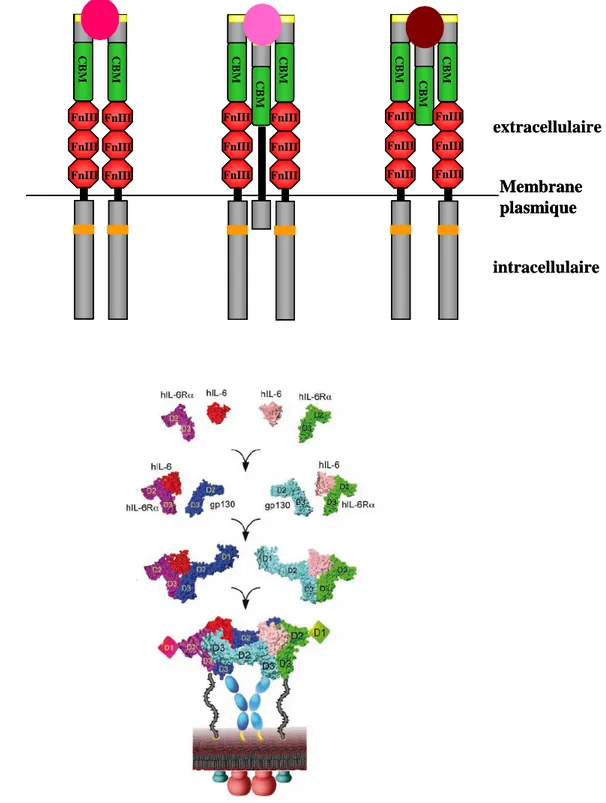
Article : Control of blood cell homeostasis in Drosophila larvae by the Posterior Signalling Centre
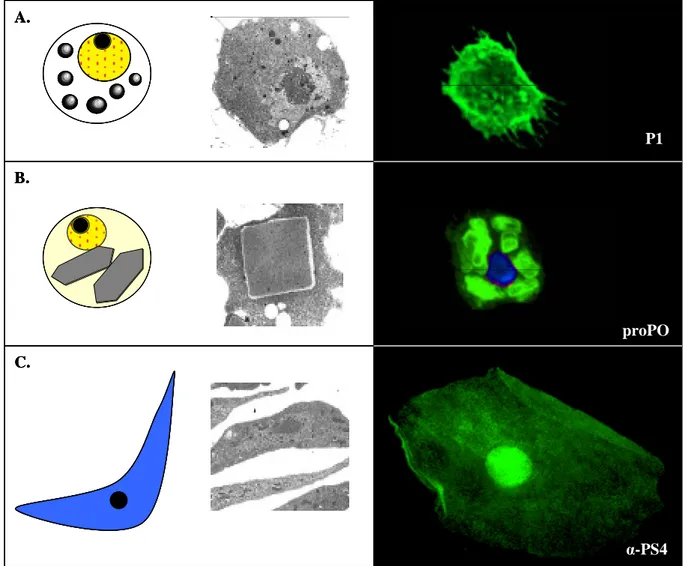
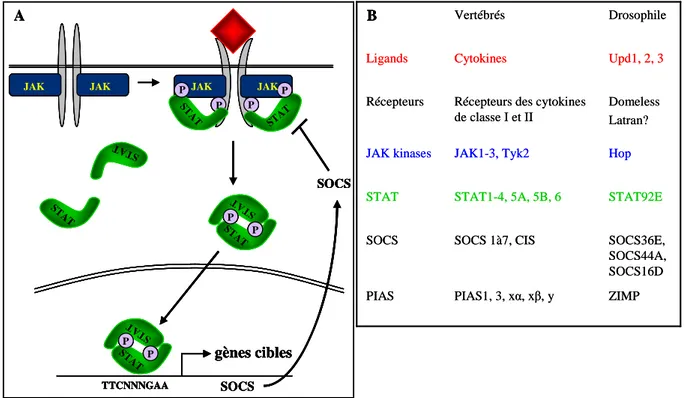
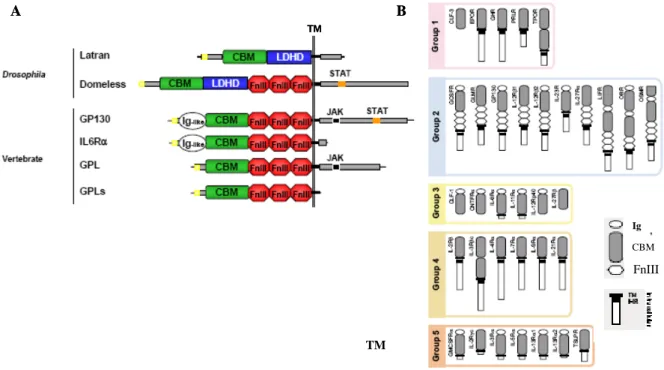
Manuscrit : A short receptor down-regulates JAK/STAT signalling to control the Drosophila cellular immune response.
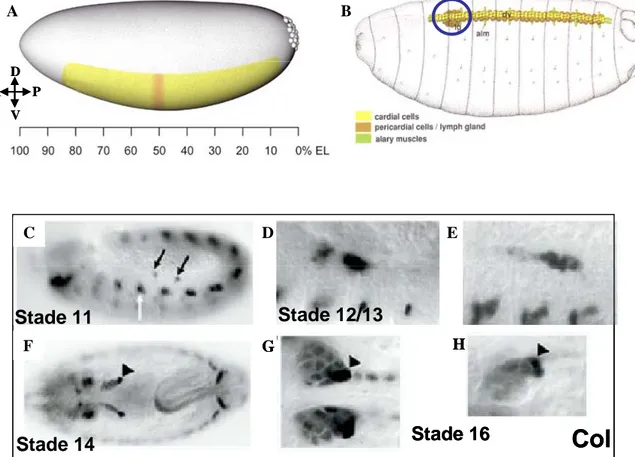
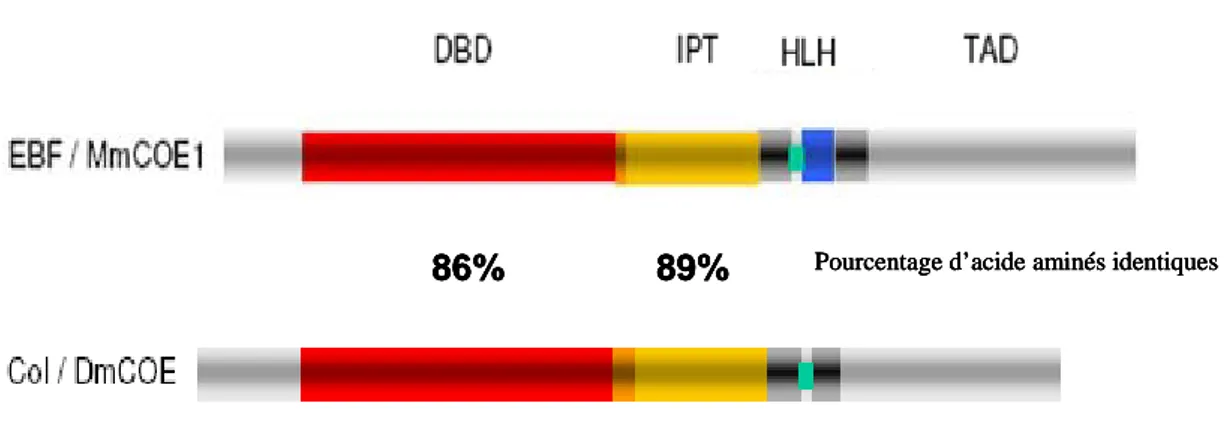
I l’expression de dome-MESO, le rapporteur de l’activité de la voie de signalisation
CONCLUSIONS GENERALES
Documents relatifs
