Toothed whales in the northwestern Mediterranean: Insight into their feeding ecology using chemical tracers
Texte intégral
Figure
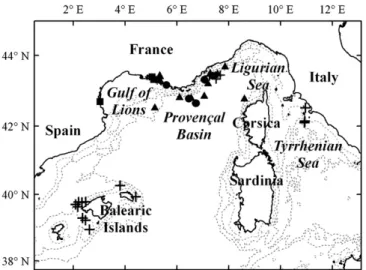
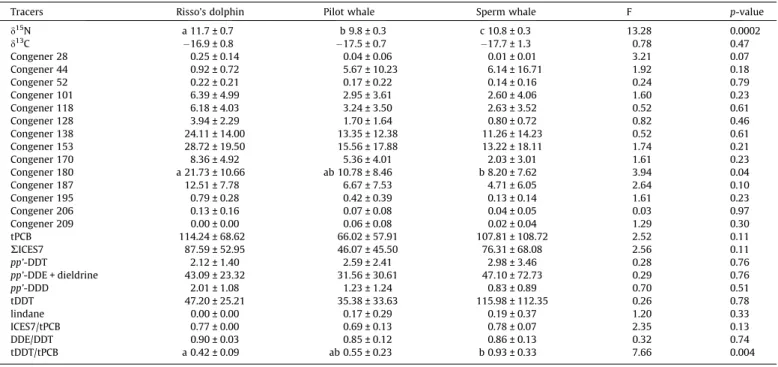
Documents relatifs
In this work, bulk oxycarbides sintered by Spark Plasma Sintering (SPS) from well controlled powders are used as references in order to evaluate IBA
142 with the IncN reference plasmid R46 demonstrated that they both contained the typical IncN plasmid scaffold, including the replicon gene (repA), the stbA-stbB-stbC genes
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
De plus, les frais d’entretien sont moins élevés dans les installations utilisant l’acide sulfurique, c'est le procédé le plus utilisé, tandis que la deuxième
d’appréciation de la faute de médecins et du lien causal avec le dommage dans le cadre de poursuites pénales du chef d’atteinte à la vie et à l’intégrité physique”, op.
The central form of these algorithms is to use a distri- buted procedure, such as either of the algorithms of chapter two, to determine for each commodity
The food and feeding ecology of the poorly known southern long-finned pilot whale (Globicephala melas edwardii) was investigated using ecological tracers (muscle and
whale (Megaptera novaeangliae) groups with a calf and other social group types in the South 789. Lagoon, New Caledonia (n
