© Julien Lapointe, 2020
Impact de l'hypoventilation volontaire sur l'entraînement
de la capacité à répéter des sprints chez les joueurs de
basketball
Mémoire
Julien Lapointe
Maîtrise en kinésiologie - avec mémoire
Maître ès sciences (M. Sc.)
ii
Résumé
Dans les sports collectifs et les sports de raquette, la capacité à répéter des sprints
(repeated-sprint ability, RSA) est un facteur déterminant de la performance. L’accumulation d’efforts
à haute intensité avec des récupérations souvent incomplètes mène à un état de fatigue neuromusculaire. C’est cette tolérance à la fatigue que les entraîneurs visent à améliorer avec différentes modalités d’entraînement. Avec la quête incessante d’optimisation de la performance sportive, l’ajout de stress physiologique lors des entraînements est devenu un incontournable afin de forcer les systèmes physiologiques à s’adapter.
L’entraînement de la RSA en milieu hypoxique (i.e., un environnement appauvri en oxygène) permet d’améliorer ce déterminant davantage qu’un entraînement similaire effectué en normoxie. Cependant, les camps d'entraînement en altitude et/ou l'utilisation de générateurs de mélanges gazeux hypoxiques nécessitent une logistique et des équipements spécifiques qui peuvent être contraignants. La technique d’hypoventilation volontaire (VHL) permettrait de surmonter cette problématique tout en ayant des effets ergogéniques similaires. Sommairement, cette technique consiste à bloquer la respiration en fin d’expiration, puis d’effectuer un effort de haute intensité (i.e., sprints). Cependant, dans la littérature, il y a très peu de mesures physiologiques permettant d’expliquer les gains de performance à la suite de l’entraînement en VHL. De plus, les entraînements de VHL ont été jusqu’à présent très peu spécifiques aux demandes des sports collectifs. L’étude insérée dans ce mémoire a pour objectif principal de mieux comprendre les effets ergogéniques potentiels du VHL et, comme objectif secondaire, d’évaluer la faisabilité de cette technique combinée à des changements de direction à haute vitesse.
Les résultats rapportés dans ce mémoire démontrent une diminution du score de détérioration de la performance lors d’un test de RSA après un entraînement VHL de 4 semaines. Les adaptations physiologiques sous-jacentes à ce gain de performance sont un meilleur maintien du recrutement des unités motrices initiales, une meilleure réoxygénation lors des périodes de récupération et une diminution de la concentration de potassium extracellulaire. La technique de VHL permet donc d’optimiser l’entraînement de la RSA tout en permettant d’introduire des changements de direction spécifiques aux demandes du sport. Des facteurs périphériques et centraux expliqueraient ces gains de performance.
iii
Abstract
In team sports and racket sports, repeated sprint ability (RSA) is a critical determinant of performance. The repetition of short and intense sprints with incomplete recovery leads to the development of neuromuscular fatigue. For coaches, the main goal of training is to improve fatigue tolerance with different training methods. In this never-ending quest for training optimisation, the use of extreme environments has become very popular among sport scientists to increase the stress placed on the athletes.
Performing RSA training in hypoxia (i.e., with less oxygen) can enhance several physiological factors and thereby improve sprint endurance more than the same training performed in normoxia. However, attending a training camp at terrestrial altitude and/or using hypoxic generators requires specific logistics and equipment that can be prohibitive. The voluntary hypoventilation (VHL) technique could overcome this problem while potentially having similar ergogenic effects. Briefly, this technique consists of blocking one’s breathing at the end of a normal expiration before attempting to sprint. However, the physiological changes associated with this technique have been poorly investigated. The study presented in this thesis aimed at understanding varied physiological adaptations following VHL training, and to assess the feasibility of performing VHL during abrupt changes in direction.
The current results demonstrate an improvement in sprint endurance following a 4-week VHL training. Physiological adaptations underlying this performance gain included a better maintenance of initial recruitment of motor units, greater muscle re-oxygenation during recovery periods and a decrease in extracellular potassium concentration. We conclude that the VHL technique can be successfully coupled with rapid changes of direction specific to the demands of the sport. Both peripheral and central mechanisms appear to underly these adaptations
iv
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières ... iv
Liste des tableaux ... vii
Liste des illustrations... viii
Liste des abréviations ... x
Remerciements... xii
Avant-propos ... xiii
Introduction ... 1
Chapitre 1 Revue de littérature ... 3
1.1. Capacité à répéter des sprints ... 3
1.1.1. Définition ... 3
1.1.2. Test de capacité à répéter des sprints et déclin de la performance ... 3
1.1.3. Facteurs limitants de la capacité à répéter des sprints ... 6
1.2. L’entraînement de la capacité à répéter des sprints ... 14
1.2.1. L’entraînement des qualités neuromusculaires ... 14
1.2.2. L’entraînement en hypoxie ... 16
1.3. Entraînement en apnée ... 23
1.3.1. L’hypoventilation volontaire à volume pulmonaire réduit ... 23
1.3.2. Les effets aigus du VHL en sprints répétés ... 25
1.3.3. Les effets de l’entraînement en VHL ... 27
1.4. Techniques de mesure des paramètres physiologiques ... 30
v
1.4.2. Électromyographie de surface ... 32
Chapitre 2 : Problématique d’étude et hypothèse de travail ... 35
Chapitre 3 : Article scientifique ... 36
3.1. Titre et résumé en français ... 36
3.2. Abstract ... 38
3.3. Introduction ... 39
3.4. Materials and Methods ... 40
3.4.1. Participants ... 40
3.4.2. Experimental Design ... 41
3.4.3. Repeated-sprint ability training... 42
3.4.4. Repeated-Sprint Ability Test ... 42
3.4.5. 30-15 Intermittent Fitness Test ... 43
3.4.6. Near-infrared Spectroscopy Measurements ... 43
3.4.7. Electromyographic Acquisition and Analysis ... 44
3.4.8. Blood Sampling... 45 3.4.9. Statistical Analysis ... 45 3.5. Results ... 46 3.5.1. Performance ... 46 3.5.2. Muscle Oxygenation ... 46 3.5.3. Electromyographic Activity ... 47
3.5.4. Blood Sample Analysis ... 47
3.5.5. Perceptual Exercise Responses ... 48
3.6. Discussion ... 48
3.7. Conclusion ... 53
3.8. References ... 53
vi
3.10. Figures ... 60
Chapitre 4 : Discussion ... 65
Conclusion ... 71
vii
Liste des tableaux
Tableau 1-1. Effets comparés de l’entraînement de sprints répétés en hypoxie vs en
normoxie sur les performances et les réponses physiologiques ... 19 Tableau 1-2. Effets comparés de l’entraînement de sprints répétés en hypoventilation
volontaire vs en normoxie sur les performances et les réponses physiologiques ... 28 Table 3-1. Mean changes in performance and perceptual exercise responses in the
repeated-sprint ability (RSA) and the 30-15IFT tests after repeated-repeated-sprint training performed with voluntary hypoventilation at low lung volume (VHL) or normal breathing (CTL). ... 57 Table 3-2. Mean changes in blood parameters following the RSA test after repeated-sprint
training performed with voluntary hypoventilation at low lung volume (VHL) or normal breathing (CTL). ... 58
viii
Liste des illustrations
Figure 1-1. Évolution des puissances moyennes au cours des 10 sprints du test RSA de type boucle fermée. Le protocole de test était de 10 sprints de 6-s sur ergocycle avec 30-s de récupération passive entre les répétitions (Mendez-Villanueva, Hamer, et Bishop 2008).. ... 5 Figure 1-2. L’influence de l’élévation du K+ extracellulaire sur la dépolarisation de la membrane (1) et la stimulation des groupes III/IV des nerfs afférents diminuant l’influx nerveux central (2). (Christiansen, 2019) ... 7 Figure 1-3. Contribution des voies métaboliques lors d’une séance de sprints répétés du premier sprint (a) au dernier (b). (Girard et al., 2011) ... 8 Figure 1-4. Régulation du H+ musculaire par le système tampon (Bishop et al., 2011) ... 10 Figure 1-5. Le développement de la fatigue neuromusculaire durant un exercice de sprints
répétés. (Collins et al., 2018) ... 12 Figure 1-6. Résumé des composantes de la RSA et des paramètres clés d’entraînement, inspiré de Bishop et al. (2011) ... 15 Figure 1-7 Principales réponses aiguës de la réduction de la disponibilité en oxygène
(hypoxie) lors d’un exercice de sprints répétés. O2 : oxygène, PCr : phosphocréatine, [H+] : concentration d’ions hydrogènes, [Lac-] : concentration d’ions lactates et [K+] : concentration d’ions potassiums. Inspiré de (Girard et al., 2017).. ... 17 Figure 1-8 Changements dans l’expression des acides ribonucléiques messagers (mRNA)
de certains gènes de transcriptions avant et après un entraînement de sprints répétés en hypoxie (Faiss et al., 2013). ... 22 Figure 1-9 : Schéma des volumes respiratoires. En rouge, le volume cible lors de l’utilisation de la technique d’hypoventilation volontaire à volume pulmonaire réduit. (Widmaier et al., 2009) ... 24 Figure 1-10. Représentation schématique du déplacement des rayons infrarouges émis entre
l’émetteur et le récepteur du NIRS. La profondeur du signal dépend de l'épaisseur de la peau et du tissu adipeux. Plus cette couche est mince, plus les rayons pénètrent dans le muscle. Et à l’inverse, plus elle est large, moins les rayons pénètrent le muscle. Schéma inspiré de Hamaoka & McCully (2019) ... 31 Figure 1-11. L’évolution de la désoxyhémoglobine ([HHb]) exprimée en pourcentage de la
valeur de repos lors d’un exercice de sprints répétés de 10 x 6-s sur ergocycle avec 30-s de récupération. (Racinai30-s et al., 2007)... 32 Figure 1-12. Schéma représentant le fonctionnement de l’électromyographie de surface
ix
Figure 1-13. Signaux de différents paramètres d’EMG. À gauche, fréquences du spectre de puissance du signal EMG. À droite, signaux bruts d’activité électrique (en haut) et signaux corrigés d’EMG (en bas). (Konrad, 2005) ... 34 Figure 3-1. Layout of repeated- sprint exercise with changes of direction (COD) on a basketball court.. ... 60 Figure 3-2. Completion time for every sprint of the RSA test performed with normal breathing (CTL) and voluntary hypoventilation at low volume (VHL) before and after four weeks of repeated-sprint training. Data are presented as means ± SE. *, indicates small effect between groups. ... 61 Figure 3-3. Peak (A) and nadir (B) values of normalised deoxyhemoglobin concentration ([HHb]) over 11 sprint/recovery cycles with normal breathing (CTL) and voluntary hypoventilation at low lung volume (VHL) before and after four weeks of training. Data are presented as means ± SD, expressed as a percent of the first sprint/recovery cycle. **, indicates moderate effect between groups; †, indicates large effect between groups.. ... 62 Figure 3-4. Percentage difference and qualitative interference in the change in NIRS variables from Pre- to Post- in VHL compared to CTL.. ... 63 Figure 3-5. Changes in normalised EMG amplitude (RMS) (A) and median power frequency (MPF) (B) of the biceps femoris and gastrocnemius muscles during 12 sprints with normal breathing (CTL) and voluntary hypoventilation at low lung volume (VHL) before and after four weeks of training. Data are presented as means ± SD, expressed as a percent of sprint 1. †, indicates large effect between groups.. ... 64
x
Liste des abréviations
[Ca++] : Concentration d'ions calcium
[cHCO3-] : Concentration d'ions bicarbonates calculée
[cHgb] : Concentration d'hémoglobine calculée [Cl-] : Concentration d'ions chlorures
[cTCO2] : Concentration totale de CO2
[Glu] : Concentration de glucose
[HbO2] : Concentration d’oxyhémoglobine
[HHb] : Concentration de désoxyhémoglobine
[HHb]max : Concentration maximale de désoxyhémoglobine
[HHb]min : Concentration minimale de désoxyhémoglobine
[K+] : Concentration d'ions potassium
[Lac-] :Concentration d'ions lactates [Na+] : Concentration d'ions sodium
[tHb] : Concentration totale d'hémoglobine
[tHb]max : Concentration totale d'hémoglobine maximale
[tHb]min : Concentration totale d'hémoglobine minimale
∆[HbO2] : Amplitude de changement de l'oxyhémoglobine
∆[tHb] : Amplitude de changement de l'hémoglobine totale 30-15IFT : 30-15 intermittent fitness test
ATP : Adénosine triphosphate
BE(b) : Excès de base du sang (base excess of blood)
BE(ecf) : Excès de base des fluides extracellulaires (base excess of extra cellular fluid) BF : Biceps femoris
Ca++ : Ions calcium
CLM : Course contre la montre
CMJ : Saut en contre-mouvement (counter mouvement jump) CNS : Système nerveux central (central nervous system) CO2 : Dioxyde de carbone
COD : Changements de direction (change of direction) CTL : Groupe contrôle
EMG : Électromyographie de surface ES : Taille d'effet (effect size)
FC : Fréquence cardiaque
FI : Indice de fatigue (fatigue index) FiO2 : Fraction d’O2 inspiré
GAS : Gastrocnemius lateralis H+ : Ions hydrogène
HbO2 : Oxyhémoglobine
HCO3- : Ions bicarbonates
Hct : Hématocrite
HHb : Désoxyhémoglobine K+ : Ions potassium
Lac- : Ions lactates
xi
MPF : Médiane du spectre de fréquence (median power frequency) Na+ : Ions sodium
NaHCO3 : Sodium bicarbonate
NIRS : Spectroscopie proche-infrarouge (near-infrared spectrocopy) NKA : Pompe d’ions sodium/potassium-adénosine triphosphate NS : Changement non significatif
O2 : Oxygène
PCO2 : Pression partielle de CO2
PCr : Phosphocréatine pH : Potentiel hydrogène Pi : Phosphate inorganique
Pmax : Puissance maximale
Pmoy : Puissance moyenne
P-MRS : Résonance magnétique par spectroscopie du phosphate PO2 : Pression partielle d'O2
Rép : Répétitions
RMS : Activité électrique totale (root mean square)
RPE : Échelle de perception de l'effort (rating of perceived exertion) RS : Sprints répétés (repeated sprints)
RSA : Capacité à répéter des sprints (repeated-sprints ability) RSAbest : Meilleure performance lors du test de RSA
RSAmean : Moyenne des performances lors du test de RSA
RSASdec : Socre de détérioration de la performance lors du test de RSA
RSH : Sprints répétés en condition hypoxique (repeated sprints hypoxia) RSN : Sprints répétés en condition normoxique (repeated sprints in normoxia)
S : Performance d'un sprint
SD : Écart-type (standard deviation)
Sdec : Score de détérioration de la performance
SpO2 : Saturation artérielle en O2
s: seconde
TOI : Indice d'O2 du tissu (tissu oxygen index)
VCO2 : Volume de CO2 expiré
VE : Ventilation
VHL : Hypoventilation volontaire à volume pulmonaire réduite VIFT : Vitesse finale lors du test 30-15 Intermittent Fitness Test
Vmax : Vitesse maximale
VO2 : Consommation systémique d’O2
VO2max : Consommation maximale d’O2
YYIR1 : Yo-Yo intermittent fitness test level 1 YYIR2 : Yo-Yo intermittent fitness test level 2
xii
Remerciements
Tout d’abord, je tiens à remercier ma famille proche (Marianne, Étienne, Marc et Andréa). Vous êtes une source de motivation et de persévérance dans laquelle j’ai pu puiser lors de la réalisation de mon projet. Merci pour votre écoute, intérêt et temps qui m’a permis d’explorer différentes perspectives. Bien entendu, merci à ma copine Marie d’avoir su me soutenir et me conseiller tout au long de ce travail.
Ensuite, je veux remercier les membres de l’équipe SciencePerfo, Léandre, Maxime et Simon, pour votre temps et votre implication dans l’acquisition des données de performance. Merci aux étudiants aux études supérieures, Pénélope, Jean-François, Shannon et Valérie pour votre aide lors des séances de test. Merci aussi à mes amis qui m’ont aidé comme cobaye lors de mes séances de testing.
Merci aux athlètes et entraîneurs du Rouge et Or basketball pour votre participation et votre confiance.
Finalement, j’aimerais remercier mon directeur de recherche, François Billaut. Tu as été une ressource incroyable tout au long du processus. Merci d’avoir partagé tes connaissances, ta philosophie de travail et surtout ta passion. Tes questions ont su me faire progresser et faire de moi un chercheur.
xiii
Avant-propos
L’article inséré dans ce mémoire s’intitule Impact of breath-hold training on muscle
oxygenation, myoelectrical changes, systemic [K+] and repeated-sprint ability in
basketball players. L’article a été révisé par les pairs et a été publié le 3 avril 2020 dans la
revue indexée Frontiers in Sports and Active Living dans la section Elite Sports and
Performance Enhancement. Je suis l’auteur principal et j’ai agi en tant que coordonnateur
à chacune des étapes du processus de recherche (rédaction du protocole de recherche, approbation éthique du projet, recrutement des participants, expérimentations, analyse des données et rédaction).
François Billaut, Ph. D., professeur-chercheur au département de Kinésiologie de l’Université Laval, est le directeur de mes travaux de recherche. Il a grandement contribué à toutes les étapes de la réalisation du présent projet.
Pénélope Paradis-Deschênes, étudiante au doctorat au département de Kinésiologie de l’Université Laval, a contribué lors de la phase expérimentale pour la collecte de données et lors de la phase d’analyse des données.
Xavier Woorons, Ph. D., professeur-chercheur à l’unité de Recherche Pluridisciplinaire Sport Santé Société à l’Université de Lille, l’Université de Artois et l’Université du Littoral Côte d’Opale, France. Il a contribué au développement de la question de recherche originale, à l’analyse et l’interprétation des données et à la révision de l’article.
Frédéric Lemaître, Ph. D., professeur-chercheur à la Faculté des Sciences du Sport à l’Université de Rouen, France. Il a contribué au développement de la question de recherche originale, à l’interprétation des données et a agi en tant que réviseur critique lors de la soumission de l’article.
1
Introduction
Dans tous les sports collectifs, y compris le basketball, les joueurs doivent réaliser un éventail varié d’efforts à intensité élevée ou maximale incluant des sprints, des sauts et des déplacements latéraux. Le tout est séparé par des périodes de marche ou de course moins intenses de quelques secondes à quelques minutes permettant à l’athlète de récupérer quelque peu. Pour améliorer la condition physique des joueurs et retarder l’apparition de la fatigue neuromusculaire, les programmes d’entraînements visent à améliorer, entre autres, la capacité à répéter des sprints (repeated-sprints ability, RSA). La RSA est maintenant bien établie comme étant un déterminant critique de la performance en sports collectifs et est influencée par des facteurs physiologiques variés tels que la déplétion des substrats énergétiques, l’accumulation de sous-produits métaboliques, des changements ioniques et de l’excitabilité de la membrane musculaire et la modification des stratégies de recrutement musculaire (Billaut & Bishop, 2009; Girard et al., 2011). Dans la quête incessante d’optimisation de l’entraînement, les scientifiques du sport et préparateurs physiques ont recours à des stratégies variées afin d’augmenter le stress imposé aux athlètes lors des séances d’entraînement. Par exemple, les entraînements par intervalles de haute intensité, les aides nutritionnelles et les environnements extrêmes sont autant de possibilités devenues très populaires. Au cours de la dernière décennie, toutefois, l’utilisation d’un stress hypoxique (i.e. un environnement appauvri en oxygène) a fait l’objet d’un grand nombre d’études scientifiques expérimentales et a montré un fort potentiel d’amélioration de la RSA comparativement à un entraînement similaire réalisé en normoxie (Billaut et al., 2012; Brocherie et al., 2017).
Toutefois, l’organisation d’un camp d’entraînement en altitude et/ou l’utilisation d’appareils hypoxiques demande beaucoup de logistique et d’équipement, ce qui peut limiter la faisabilité et l’engouement pour cette modalité d’entraînement. Des chercheurs se sont alors intéressés à d’autres moyens d’induire un stress hypoxique, et l’entraînement en apnée est alors devenu très pertinent dans cette perspective. Lors d’exercices à intensité modérée où la respiration est retenue volontairement à un faible niveau pulmonaire, la saturation artérielle en oxygène (SpO2) est similaire à celle observée lors d’entraînements
2
Lorsque cette technique est appliquée aux sprints répétés (repeated-sprints, RS), l’hypoventilation volontaire à faible volume pulmonaire (VHL) permet également de diminuer la SpO2 à des moyennes de ~87%. Ceci induirait une plus grande désoxygénation
musculaire et une plus grande accumulation de lactate sanguin ([Lac-]) comparativement au même exercice avec une respiration normale (Woorons et al., 2017).
Récemment, le VHL a été appliqué lors d’un protocole d’entraînement en RS de quelques semaines et les résultats ont montré une amélioration de l’endurance de sprints à la course (Fornasier-Santos et al., 2018), à la nage (Trincat et al., 2017) et sur vélo (Woorons et al., 2019a). Toutefois, ces études n’ont mesuré que très peu de paramètres physiologiques permettant d’expliquer les gains de performance. Pour le moment, l’augmentation du [Lac
-] et de la consommation systémique d’oxygène (VO2) après l’entraînement est la seule
observation permettant de comprendre l’impact du VHL sur la RSA. De toute évidence, il y a un manque de connaissances sur les adaptations physiologiques à la suite d’un entraînement de RS en VHL et d’autres recherches sont nécessaires afin de mieux guider les pratiques sportives.
En premier lieu, ce mémoire vise à faire le point sur les connaissances actuelles concernant la RSA, les modalités d’entraînement en hypoxie et les techniques de mesure non invasives permettant d’apprécier des paramètres physiologiques clés reliés à la RSA. Par la suite, la problématique entourant le VHL sera exposée. Ensuite, le projet de recherche mis en place ainsi que les résultats seront détaillés. Pour finir, la discussion et la conclusion apporteront une contribution à l’état actuel des connaissances sur le VHL et son impact sur l’oxygénation musculaire, l’activité myoélectrique et la régulation métabolique. Ces informations permettront d’outiller les athlètes et entraîneurs afin d’incorporer cette technique dans leur planification d’entraînement.
3
Chapitre 1 Revue de littérature
1.1. Capacité à répéter des sprints
1.1.1. Définition
La capacité à répéter des sprints (RSA) est un déterminant majeur de la performance dans de nombreuses disciplines incluant une grande partie des sports collectifs (e.g. basketball, rugby, soccer) et les sports de raquettes (e.g. tennis, badminton). La RSA est donc aussi importante que l’endurance cardiovasculaire l’est pour un marathonien. C’est ce déterminant de la performance que la plupart des chercheurs et entraîneurs souhaitent développer et optimiser. Elle se définit par la capacité à répéter de brèves périodes d’effort à intensité maximale ou quasi maximale séparées par de courtes périodes de récupérations souvent passives ou à basse intensité (i.e. marche ou course). La durée des intervalles à haute intensité est souvent inférieure à 10 secondes et les périodes de récupération sont d’une durée inférieure à 60 secondes. Ces intervalles d’effort sont répétés sur des périodes allant d’une heure à quatre heures et exceptionnellement jusqu’à 10 heures lors de parties de tennis (Girard et al., 2011). De plus, les sprints répétés sont souvent combinés à de nombreux changements de direction et à un grand éventail de mouvements variés faits à haute intensité (e.g. sauts, déplacements latéraux).
L’enchaînement de sprints avec des récupérations incomplètes crée un état de fatigue qui mène à une diminution de la performance. Avant d’explorer les modalités d’entraînement, il est primordial de bien comprendre les facteurs qui mènent à l’apparition de la fatigue et comment elle impacte la performance motrice.
1.1.2. Test de capacité à répéter des sprints et déclin de la performance
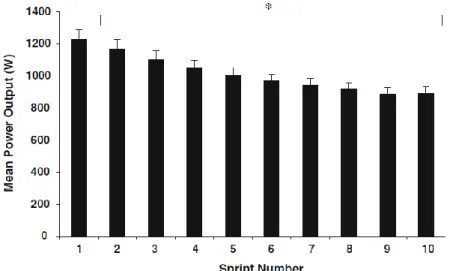
Lors de compétitions ou d’entraînements, il est difficile de quantifier précisément la charge et la fatigue associée aux RS. Ils existent de nombreuses stratégies (e.g. accéléromètres intégrés, GPS), mais le contexte de jeu, la variété des mouvements et des stratégies ne facilitent pas la tâche d’analyse de la RSA (Edwards et al., 2018). Pour pallier cette problématique, la RSA est étudiée sous forme de tests de terrain ou de laboratoire dans lesquels la détérioration de la performance est directement mesurée via la puissance4
mécanique développée. Il existe deux types de tests : à boucle fermée ou ouverte. Avant chaque test, une séance de familiarisation est nécessaire afin de déterminer la performance de référence. Pour simplifier la lecture, nous allons utiliser le sprint à la course comme modalité de test, donc ici la mesure de référence est une vitesse maximale (Vmax).
Toutefois, il est à noter que les tests de RSA peuvent se faire sous différentes modalités (e.g. ergocycle, double poussée de ski de fond, etc.) et que la mesure de référence variera en fonction de cette modalité. Peu importe le type de tests qui est utilisé, le premier sprint doit correspondre à au moins 95% de la vitesse de référence. La durée des sprints et des récupérations est standardisée et certains exemples sont présentés dans les Tableaux 1-1 et 1-2 (pages 18 et 27, respectivement). Ce qui différentie les deux types de tests est le critère d’arrêt du test. Pour le test à boucle ouverte, le participant doit compléter le maximum de sprints jusqu’à l’épuisement qui est prédéfini à un seuil habituel de 85% de la vitesse de référence. La performance correspond alors au nombre de sprints complétés avant l’épuisement, la performance de chacun des sprints (S) ou la performance moyenne. Le test à boucle fermée a un nombre fixe de sprints et le test prend fin lorsque le dernier sprint est complété (voir Figure 1-1). La performance peut cette fois être attribuée aux performances de sprints, leurs moyennes, à un indice de fatigue (FI) ou un score de détérioration de la performance (Sdec). L’avantage de la boucle fermée est de pouvoir comparer chacun des
sprints à l’ensemble du test de RSA. Voici les calculs utilisés où le symbole S correspond à la performance d’un sprint (Girard et al., 2011; Glaister et al., 2008).
𝐹𝐼 = 100 × 𝑆𝑀𝑒𝑖𝑙𝑙𝑒𝑢𝑟𝑒 − 𝑆𝑃𝑖𝑟𝑒 𝑆𝑚𝑒𝑖𝑙𝑙𝑒𝑢𝑟𝑒
𝑆𝑑𝑒𝑐 (%) = ( 𝑆1+ 𝑆2+ 𝑆3+ . . . +𝑆𝑓𝑖𝑛𝑎𝑙
𝑆𝑚𝑒𝑖𝑙𝑙𝑒𝑢𝑟𝑒 × 𝑛𝑜𝑚𝑏𝑟𝑒 𝑑𝑒 𝑠𝑝𝑟𝑖𝑛𝑡𝑠− 1) × 100
À noter pour le calcul de Sdec, si le temps de sprint correspond au paramètre S, la précédente
formule s’applique. Cependant, si le paramètre de performance est une vitesse ou puissance, l’équation suivante doit être appliquée.
𝑆𝑑𝑒𝑐 (%) = (1 − 𝑆1+ 𝑆2+ 𝑆3+ . . . +𝑆𝑓𝑖𝑛𝑎𝑙
5
Selon la littérature, le calcul de Sdec est considéré comme une méthode plus valide et
reproductible pour quantifier la fatigue comparativement à l’utilisation de FI (Glaister et al., 2008). En effet, le calcul de Sdec prend en compte l’ensemble des sprints alors que le FI
considère seulement les deux extrêmes. De plus, le premier sprint a un impact considérable sur le Sdec comme l’exprime la formule où la meilleure performance est habituellement le
premier sprint. Plusieurs auteurs ont en effet observé une corrélation positive entre la performance du premier sprint et la détérioration de performance des sprints subséquents (Bishop et al., 2003; Mendez-Villanueva et al., 2008; Yanagiya et al., 2003). En d’autres termes, plus la performance du premier sprint est élevée, plus les perturbations physiologiques qui s’ensuivent (et engendrant une fatigue neuromusculaire) sont importantes. C’est pour cette raison qu’un seuil de 95% de la vitesse de référence est utilisé lors du premier sprint.
Il y a plusieurs autres paramètres influençant les tests de RSA, parmi ceux-ci nous retrouvons par exemple le contexte environnemental et le moment de la journée auquel le test est exécuté, les différences sexuelles, l’âge, le niveau d’entraînement des participants. Le déclin de performance mesuré au cours d’un test de RSA permet donc d’évaluer et de quantifier l’apparition de la fatigue. La prochaine section détaillera les principaux facteurs physiologiques reliés à la performance en RS. Une meilleure compréhension de ces facteurs permettra d’influencer les interventions en vue d’optimiser la RSA.
Figure 1-1. Évolution des puissances moyennes au cours des 10 sprints du test RSA de type boucle fermée.
Le protocole de test était de 10 sprints de 6-s sur ergocycle avec 30-s de récupération passive entre les répétitions (Mendez-Villanueva, Hamer, et Bishop 2008).
6
1.1.3. Facteurs limitants de la capacité à répéter des sprints
Il existe un grand nombre de facteurs limitants de la performance lors d’un test de RSA. Ces facteurs limitants peuvent être localisés dans les structures périphériques (i.e. muscles squelettiques) ou avoir une origine plus centrale (i.e. système nerveux central, CNS). Il est cependant généralement difficile de dissocier complètement ces deux familles de facteurs, car ces milieux s’influencent mutuellement lors d’un exercice épuisant. Les sections suivantes mettent en lumière l’influence de la fatigue périphérique et centrale sur la RSA.
1.1.3.1. Facteurs limitants périphériques Excitabilité de la membrane musculaire
Les changements de concentrations ioniques de part et d’autre de la membrane permettent la propagation de l’influx nerveux, c’est l’excitabilité de la membrane musculaire. Au repos, il y a une distribution inégale des principaux ions, le sodium (Na+) et le potassium
(K+). Il y a une plus grande concentration de Na+ ([Na+]) dans le milieu extracellulaire et une plus grande concentration de K+ ([K+]) dans le milieu intracellulaire. Le maintien de ce gradient de concentration est possible grâce à l’activité de la pompe Na+/K+-adénosine
triphosphate (NKA). Lors d’une contraction musculaire, des canaux régulés situés sur le sarcolemme et les tubules-T permettent l’entrée rapide du Na+ et la sortie du K+ ce qui créer une dépolarisation. Cette dépolarisation se propage tout le long de la membrane. Ce potentiel d’action déclenche la libération des ions calcium (Ca++) au niveau des citernes
terminales du réticulum sarcoplasmique et permet la contraction musculaire. L’activité de la NKA permet de rétablir le gradient initial et ainsi préparer le sarcolemme à une prochaine dépolarisation (Christiansen, 2019).
Cependant, l’homéostasie de la membrane est compromise après des contractions musculaires dynamiques répétées (Fraser et al., 2002; Juel et al., 2000; Nielsen et al., 2004). L’activité de la NKA ne permet plus de maintenir le gradient adéquat et ceci entraîne une augmentation de la [K+] extracellulaire et une diminution de la [K+] intracellulaire. Le
phénomène inverse est observé pour les [Na+] (Christiansen, 2019; McKenna et al., 2008).
Cette perturbation de l’homéostasie membranaire provoque une réduction de l’amplitude du potentiel d’action et d’un ralentissement de l’influx nerveux vers les fibres musculaires
7
sollicitées (Christiansen, 2019; Girard et al., 2011). De plus, l’augmentation du K+ extracellulaire pourrait aussi causer une hausse de l’activation des groupes nerveux afférents III/IV ce qui diminuerait l’influx nerveux central vers les muscles sollicités (Amann et al., 2013). Ce bouleversement de l’homéostasie membranaire à de nombreux autres impacts sur la fonction de la cellule musculaire (Figure 1-2). Ainsi, la [K+] est un excellent marqueur de l’excitabilité de la membrane et son interprétation permet d’apprécier la contribution des facteurs limitants périphériques sur la performance.
Filières énergétiques et disponibilité des substrats
Plusieurs études utilisant la technique de biopsie musculaire ont analysé la contribution des diverses voies métaboliques à la fourniture d’énergie lors d’exercices de sprint (pour revue, voir Billaut & Bishop, 2009; Glaister et al., 2008). Lors du premier sprint d’un exercice de RS, la principale voie métabolique impliquée dans la production d’adénosine triphosphate
Figure 1-2. L’influence de l’élévation du K+ extracellulaire sur la dépolarisation de la membrane (1) et la
8 (ATP) est le métabolisme de la phosphocréatine (PCr), y contribuant pour 52% (Figure 1-3). Par la suite, la glycolyse anaérobie lactique contribue pour 40% et la glycolyse aérobieq pour 8%. Lors du dernier sprint, la contribution de la PCr est similaire à celle du premier sprint, soit environ 50%. Cependant, la contribution de la glycolyse anaérobie lactique et aérobie s’est inversée (9% vs 40%, respectivement) pour obtenir une forte majorité de l’énergie de la filière aérobie (Buchheit et al., 2009; Ratel et al., 2006).
La PCr représente la réserve d’énergie la plus immédiate lors d’exercices de RS étant donné la demande importante de re-phosphorylation de l’ATP qui est grandement utilisée pour ce type
d’effort (Hultman & Sjöholm, 1983). Cependant, les réserves en PCr peuvent diminuer de 35 à 55% après un sprint unique de 6 secondes (Gaitanos et al., 1993) et la récupération complète des réserves en PCr peut prendre 5 minutes (Bogdanis et al., 1995). La contribution de la PCr dépend en partie de sa quantité initiale dans le muscle squelettique, mais aussi de sa resynthèse lors des périodes de récupération entre les sprints. En effet, même si la resynthèse complète peut prendre 5 minutes, des périodes de 30 secondes seulement permettent une récupération suffisante afin de répéter plusieurs sprints de 6 secondes sans diminution de performance significative chez des athlètes modérément entraînés en sport collectif. Cette resynthèse rapide est également corrélée à la performance lors des 5 derniers sprints (r= 0,67) dans une série de 10 sprints, indiquant sa grande importance pour la RSA (Mendez-Villanueva et al., 2012). Cependant, la resynthèse de la PCr est hautement dépendante de la disponibilité en O2 (Girard et al., 2011; Haseler et al.,
1999). Ceci peut engendrer une dette en O2 qui est palliée par une augmentation de
l’activité aérobie en période de récupération, expliquant la contribution de plus en plus importante au cours d’un test de RS (9% à 40%). Par ailleurs, cette élévation de l’activité oxydative reflète aussi une augmentation de la contribution de la voie aérobie à la
Figure 1-3. Contribution des voies métaboliques lors d’une séance de sprints répétés du premier sprint (a) au dernier (b). (Girard et al., 2011)
9
phosphorylation de l’ATP. Ainsi, un autre facteur limitant la performance est la consommation maximale d’oxygène (VO2max) qui peut être atteinte lors des derniers sprints
(Dupont et al., 2005; McGawley & Bishop, 2015). Nous notons en effet une corrélation significative et négative (r= -0,45 à -0,75) entre le VO2max et les indices de fatigue lors d’un
test de RSA (FI ou Sdec) (Bishop et al., 2003; Brown et al., 2007; Rampinini et al., 2009).
Finalement, la filière anaérobie lactique perd rapidement en proportion avec l’enchaînement des sprints et ceci peut être dû à la cinétique de la filière oxydative aérobie qui prend un certain temps pour s’activer (Dupont et al., 2005; Girard et al., 2011). Nous avons observé une corrélation positive (r= 0,71 à 0,85) indiquant que plus la cinétique de VO2 est rapide, plus le Sdec tend à être petit (Dupont et al., 2005, 2010). De plus, la
contribution importante de la glycolyse anaérobie en début d’effort induit une acidité importante du milieu musculaire qui réduit l’efficacité des enzymes phosphofructokinase et glycogène phosphorylase ce qui altère le flux métabolique de la glycolyse et, par le fait même, diminue sa contribution (Spriet et al., 1989). Ainsi, la contribution de la voie anaérobie lactique serait alors très transitoire.
Toutefois, la mise en action très rapide et très intense des filières énergétiques n’est pas sans conséquence pour la contraction musculaire. Plusieurs résidus métaboliques issus de ces réactions chimiques s’accumulent au niveau musculaire et contribuent à la baisse de performance. Bien connaître les effets délétères de ces composés métaboliques permet également de réaliser des séances d’entraînement ciblant de façon spécifique des facteurs clés de la résistance à la fatigue dans les sports intermittents.
Accumulation de métabolites
Les métabolites sont des produits résultants des réactions chimiques permettant la production d’énergie via les différentes voies métaboliques vues précédemment. Dans une perspective d’amélioration de la RSA, deux résidus métaboliques ont été particulièrement étudiés et leur élimination corrélée à la performance : les ions hydrogène (H+) et le phosphate inorganique (Pi).
Les ions H+ proviennent essentiellement de la glycolyse anaérobie lactique. En effet, lors
de l’utilisation ce cette voie, il y a production d’acide lactique qui est immédiatement transformé en ion lactate (Lac-) et en un H+ par la réaction suivante :
10
𝐶3𝐻6𝑂3 → 𝐶3𝐻5𝑂3−+ 𝐻+
Lors de sprints, l’intensité est telle que la glycolyse fonctionne à plein régime et entraîne une accumulation forte de ces ions dans le muscle. De manière générale, l’accumulation importante d’H+ diminue le potentiel hydrogène (pH) et par le fait même la performance
(Bishop et al., 2003; Bishop & Edge, 2006). Plus précisément, l’augmentation du H+ pourrait impacter la fatigue en activant les groupes afférents III/IV, en diminuant la libération du Ca++ au niveau du réticulum sarcoplasmique via l’inhibition de la NKA, en
réduisant l’affinité des liaisons actine-myosine dans les fibres musculaires, en diminuant la production de l’ATP par la voie glycolytique et bien d’autres mécanismes (Cairns, 2006; Sundberg & Fitts, 2019).
Deux phénomènes permettent de limiter l’apparition de la fatigue en influençant l’accumulation de H+ musculaire. Premièrement, la capacité tampon permet de réguler le
pH en fixant le H+ à un ion bicarbonate (HCO3-) afin de l’éliminer. Plusieurs travaux ont
bien démontré que la capacité tampon est corrélée à une diminution du Sdec lors d’un test
de RSA, soit une meilleure résistance à la fatigue (Bishop et al., 2003, 2004; Bishop & Edge, 2006). D’autres auteurs ont également mis en évidence une relation de cause à effet entre l’ingestion de bicarbonate de sodium (NaHCO3) et la diminution de la manifestation
de la fatigue (Peart et al., 2012; Stecker et al., 2019), démontrant les effets bénéfiques d’une réduction de la concentration d’ions H+. Le NaHCO
3 agit comme agent alcalin réduisant
l’acidité musculaire, toutefois certains résultats sont contradictoires. Le système tampon offert par les
ions HCO3- n’est efficace qu’à
l’extérieur du muscle et, donc, les ions H+ produits par les fibres
musculaires doivent être « évacués » vers le plasma sanguin (voir Figure 1-4). Ainsi le transporteur monocarboxylate musculaire (MCT) joue un rôle crucial dans la sortie du Lac- et du
Figure 1-4. Régulation du H+ musculaire par le système tampon
11
H+ (Thomas et al., 2005). L’activité du MCT permet donc de réguler efficacement le pH intramusculaire et ainsi limiter l’apparition de la fatigue (Juel, 1998).
Le deuxième métabolite en jeu est le Pi, soit le produit de l’hydrolyse de la PCr pour
synthétiser des molécules d’ATP. Seul le Pi a un effet négatif sur la fatigue alors que le groupe créatine n’a pas d’influence majeure (Westerblad et al., 2002). L’accumulation importante du Pi à la suite d’efforts très intenses a un effet négatif sur la fonction contractile
du muscle en réduisant le nombre de ponts transversaux d’actine-myosine ce qui a pour effet direct d’inhiber la force et la puissance musculaire. Cette réduction du travail musculaire est due à une réduction à la fois de la quantité de Ca++ libéré par le réticulum sarcoplasmique et de la sensibilité des myofilaments au Ca++ (Sundberg & Fitts, 2019).
S’il est pertinent d’observer et de comprendre les effets individuels de chacun des métabolites, il est aussi important d’étudier l’effet synergique de la présence de ces deux métabolites sur la fatigue. En d’autres termes, cette synergie pourrait bien être plus importante que la simple addition des effets individuels de chaque métabolite (Karatzaferi et al., 2008; Nelson et al., 2014). Pour le moment, peu d’études ont étudié ce phénomène et les mécanismes sous-jacents de cette synergie ne sont pas encore élucidés (Sundberg & Fitts, 2019).
1.1.3.2. Facteurs limitants centraux Système et influx nerveux
Pour qu’un muscle squelettique se contracte, ses fibres musculaires doivent recevoir l’ordre de se contracter. Ainsi, le CNS envoie les commandes motrices efférentes adéquates vers les muscles par la propagation de l’influx nerveux. Le recrutement du nombre d’unités motrices et leur fréquence de stimulation sont des composantes de cette commande motrice et elles dépendent des centres de contrôle volontaire et réflexes (i.e. centres supra-spinaux et spinaux respectivement). Avec la répétition d’efforts, plusieurs facteurs peuvent altérer la commande motrice et ainsi contribuer à la diminution de la puissance, c’est ce que l’on nomme la fatigue centrale (Collins et al., 2018). Pour mesurer l’influx nerveux et la commande motrice, les chercheurs ont recours à la technique de stimulation transcutanée et à l’électromyographie de surface (EMG). Si la stimulation électrique ou magnétique
12
reste la technique étalon pour évaluer la fatigue centrale, elle est difficile à mettre en place hors du laboratoire puisqu’elle nécessite plusieurs équipements encombrants. On lui préfère alors l’EMG qui permet, malgré des limites méthodologiques connues, d’estimer le recrutement musculaire. Celle-ci est décrite en détail à la section 1.3.2. Somme toute, à l’aide de ces deux techniques combinées, les études scientifiques démontrent bien qu’au cours d’un exercice de sprints répétés, la fatigue neuromusculaire qui se développe lors des premiers sprints est principalement d’origine périphérique et s’accentue jusqu’à la fin de l’exercice. La fatigue centrale, quant à elle, semble apparaître dans la deuxième moitié de ce type d’exercice et se surimpose à la fatigue périphérique dans l’expression totale de la fatigue neuromusculaire (Hureau et al., 2016). La Figure 1-5 illustre bien la synergie entre ces deux types de fatigue au cours d’exercices de sprints répétés. C’est pourquoi la diminution de la force produite lors d’une contraction maximale volontaire est plus faible (-17%) que celle mesurée à l’aide d’une stimulation transcutanée du nerf moteur (-54%) immédiatement après un exercice de RS sur ergocycle (Billaut et al., 2013). La diminution de la force produite par une contraction maximale volontaire exprime la fatigue centrale qui est généralement plus faible que la fatigue périphérique lors de ce type d’efforts. Ceci est appuyé par une faible diminution du rapport d’activation centrale qui atteint 0,92 après l’effort (-3%) où un rapport de 1 exprime l’absence de fatigue centrale.
Figure 1-5. Le développement de la fatigue neuromusculaire durant un exercice de
13
Les mécanismes conduisant à une diminution de la commande motrice centrale ne sont pas encore complètement appréhendés. Cependant, les rétroactions sensorielles des muscles par différentes afférences (les organes tendineux de Golgi, les fuseaux musculaires, les terminaisons nerveuses des groupes III/IV) vers le CNS semblent jouer un rôle-clé. Ces informations sont intégrées par le cortex ou la moelle épinière qui peuvent alors ajuster la commande motrice. Par exemple, l’accumulation d’ions H+ ou K+ à l’extérieur de la membrane musculaire excite les groupes afférents III/IV qui remontent jusqu’au CNS et qui peuvent diminuer la prochaine commande motrice et ainsi la performance de l’exercice (Amann et al., 2011, 2013). Ensuite, au niveau de la moelle épinière, s’il y a une diminution des rétroactions sensorielles, il y a une diminution de la contraction volontaire (Macefield et al., 1993). Une diminution de l’afférence des interneurones-1α au niveau de la moelle épinière résultera en une diminution du recrutement de motoneurones-1α et ainsi du nombre d’unités motrices. Ce phénomène est mesurable à l’aide du réflexe d’Hoffman (H-réflex) où une stimulation électrique percutanée sur un nerf afférent permet une action motrice réflexe et peut être enregistrée à l’aide d’EMG (Zehr, 2002). Cette action réflexe a été mesurée récemment lors d’un protocole de 7 sprints répétés de 10-s avec 180-s de récupération. L’amplitude du H-réflex était diminuée après un seul sprint et ceci pouvait durer jusqu’à 10 minutes après l’exercice. Ainsi, il y avait une suppression de l’excitabilité réflexe qui pourrait entraîner une diminution de la performance (Pearcey et al., 2017). Finalement, un autre phénomène qui pourrait contribuer à la fatigue centrale est la stratégie de recrutement musculaire. Pour produire le maximum de force lors d’un sprint, les muscles doivent être pleinement activés lors de la phase de raccourcissement et relaxés lors de la phase d’étirement (Neptune & Kautz, 2001). Le synchronisme de la commande motrice est donc primordial pour l’exécution de la tâche et peut être altéré avec la fatigue. En effet, la puissance maximale développée lors de sprints de 6 s sur ergocycle a diminué de 11% entre le premier et le dixième sprint alors que les valeurs d’EMG initiales étaient maintenues (Billaut et al., 2005). L’analyse temporelle de la coordination musculaire indiquait une activation plus précoce des muscles agonistes par rapport au cycle de pédalage lors du dernier sprint de 92 millisecondes. Ainsi, dans la phase de pédalage, les muscles agonistes n’étaient pas pleinement recrutés au bon moment et le recrutement des muscles antagonistes était plus important. De plus, les hautes fréquences de stimulation
14
vers les fibres de type II majoritairement sollicitées lors de sprints, diminuent avec l’augmentation de la fatigue périphérique. En effet, ces fibres sont moins résistantes à la fatigue que les fibres de type I et celles-ci seront alors plus sollicitées avec des fréquences de stimulation plus basses résultant en une diminution de la puissance développée (Matsuura et al., 2006).
Cette section a mis l’accent sur les méthodes d’évaluation de la capacité à répéter des sprints maximaux, les manifestations de la fatigue neuromusculaire et les facteurs limitant la performance lors d’efforts intermittents. Les prochaines sections ont pour objectif de comprendre les effets chroniques de l’entraînement en RS sur la performance et les adaptations physiologiques.
1.2. L’entraînement de la capacité à répéter des sprints
Cette section met en valeur l’interaction et la complexité des différents paramètres à cibler dans une séance d’entraînement spécifique de la RSA et décrit les adaptations subséquentes à un protocole d’entraînement en condition normoxique et/ou normobare. De plus, ce mémoire s’intéressant aux effets potentiels de l’entraînement en apnée induisant une hypoxémie artérielle, il convient ensuite de décrire l’état des connaissances scientifiques au sujet de l’entraînement de sprints répétés en hypoxie.
1.2.1. L’entraînement des qualités neuromusculaires
Comme c’est le cas pour un grand nombre de qualités physiques, une analyse systématique des facteurs de performance de la RSA fait ressortir des composantes clés : la performance lors des sprints et la récupération entre les sprints (voir Figure 1-6). La performance lors d’un sprint dépend de deux paramètres, soit la longueur des enjambées et leurs fréquences. Pour améliorer la longueur des enjambées, il est recommandé de développer les qualités musculaires telles que la puissance, la mobilité, la capacité du muscle à restituer de l’énergie (i.e. force élastique) et la disponibilité en ATP. L’amélioration de la fréquence des enjambées passe essentiellement par un travail de la technique de course et de la coordination musculaire. Ainsi, pour améliorer la performance en sprints, l’entraîneur doit
15
élaborer des séances d’entraînement comprenant de la musculation en puissance, de la pliométrie et/ou des sprints. Pour prolonger le nombre de sprints de haute intensité, il serait primordial de cibler les déterminants énergétiques et métaboliques tels que la resynthèse de PCr, de la capacité tampon et du système aérobie. Il n’est donc pas surprenant d’observer ici les facteurs limitant la RSA vus précédemment.
Ainsi, pour développer la RSA, il est possible d’isoler chacun des paramètres en effectuant des séances variées durant une semaine d’entraînement. Toutefois, cette approche n’est pas la plus optimale si l’on considère toutes les qualités physiques, tactiques et techniques que les athlètes de sports collectifs doivent développer. L’entraînement en sprints répétés permet d’intégrer toutes ces composantes en un seul entraînement (Bishop et al., 2011). De plus, l'avantage de cette technique est de pouvoir être très spécifique au registre technique des athlètes en leur permettant d’incorporer des changements de direction (change of
direction, COD). L’ajout de COD lors d’entraînement en RS est de plus en plus populaire
et permettrait d’améliorer davantage la performance lors de tests de RSA comparativement à un entraînement avec des sprints en ligne droite (Beato et al., 2019). Le gain de performance provient du stress supplémentaire induit par la phase excentrique dynamique lors des COD (Chaabene et al., 2018) créant une augmentation des dommages musculaires (McHugh et al., 1999).
Pour mesurer l’effet de l’entraînement en RS sur la RSA, l’utilisation d’un devis expérimental avant-après avec groupe contrôle (CTL) est la plus fréquente. L’endurance de sprint est la principale variable étudiée au moyen des tests de RSA discutés
Figure 1-6. Résumé des composantes de la RSA et des paramètres clés d’entraînement, inspiré de Bishop et
16
précédemment. L’ajout de tests secondaires permet également de mesurer l’impact de l’intervention sur d’autres paramètres spécifiques de la RSA qui peuvent être jugés importants par l’entraîneur ou le chercheur. Par exemple, il est possible d’ajouter des sauts en contre-mouvement (CMJ) pour évaluer les qualités musculaires, des tests de sprints pour évaluer la performance lors de sprints et des tests aérobies et anaérobies pour évaluer la capacité de récupération des athlètes. Parmi ces tests cardiovasculaires, nous retrouvons le
Yo-Yo intermittent fitness test level 1 (YYIR1) et level 2 (YYIR2) et le 30-15 intermittent fitness test (30-15IFT) qui sont fréquemment utilisés dans les sports intermittents. À l’aide
de ces tests, il est possible de mettre en évidence la panoplie d’adaptations physiologiques induites par l’entraînement en RS. En effet, l’entraînement en RS sur une période allant de 2 à 6 semaines permet d’améliorer chacune des qualités neuromusculaires décrites plus haut (Taylor et al., 2015). Ce type d’entraînement est efficace et donc très répandu. C’est pour cette raison que les scientifiques du sport et les préparateurs physiques s’y intéressent et cherchent des stratégies afin d’augmenter le stress imposé aux athlètes lors des séances d’entraînement tel que les environnements extrêmes.
1.2.2. L’entraînement en hypoxie
L’hypoxie se définit comme une diminution de la disponibilité en O2, soit par une
diminution de la pression atmosphérique (e.g. en altitude) ou par une diminution de la fraction d’O2 dans le mélange respiré au profit de celle de l’azote (e.g. chambre ou masque
hypoxique). Cette diminution d’O2 affecte l’organisme en diminuant la pression partielle
de ce gaz dans le sang artériel et la SpO2. Il existe un grand éventail de méthodes
d’entraînement en hypoxie où l’exposition peut être de différentes durées (jours vs secondes), lors d’exercices de différentes intensités (sous-maximale vs maximale) ou appliqués de manière systémique ou locale. Toutefois, nous nous concentrerons seulement sur la modalité RSH (repeated sprints in hypoxia), soit lorsque l’exposition est continue et systémique lors de sprints répétés. Au cours d’un exercice de RSH, des modifications physiologiques surviennent au niveau du système cardio-pulmonaire, du métabolisme énergétique et de la fonction neuromusculaire. Parmi ces modifications nous retenons une augmentation de la désoxyhémoglobine musculaire, d’ions H+, K+ extracellulaires et du
17
musculaire et de la force produite. Une liste plus exhaustive des facteurs physiologiques modifiés par l’hypoxie est présentée dans la Figure 1-7. Ces modifications physiologiques impactent négativement la perception de l’effort et la qualité d’exécution du mouvement. Ainsi, l’ensemble de ces réponses physiologiques à l’hypoxie engendrent un déclin de la performance. Toutefois, ces réponses à l’hypoxie sont modulées par la nature de la tâche, la sévérité des conditions hypoxiques et l’expérience du sujet par rapport aux environnements hypoxiques (Girard et al., 2017).
Étant donné l’important stress imposé à l’exercice, l’hypoxie est ajoutée à des protocoles d’entraînement dans l’objectif de créer des adaptations supérieures à celles d’un entraînement en condition normale. Le Tableau 1-1 est un recensement des écrits à jour des adaptations à la suite d’un entraînement de RS en condition hypoxique (RSH) comparativement à un entraînement de RS en condition normoxique (RSN). Pour être incluses dans le tableau, les études devaient avoir un groupe en condition RSN et en RSH,
Figure 1-7 Principales réponses aiguës de la réduction de la disponibilité en oxygène (hypoxie) lors d’un
exercice de sprints répétés. O2 : oxygène, PCr : phosphocréatine, [H+] : concentration d’ions hydrogènes,
[Lac-] : concentration d’ions lactates et [K+] : concentration d’ions potassiums. Inspiré de (Girard et al.,
18
ne pas avoir d’exposition chronique à l’hypoxie lors de la phase d’entraînement (Live-High
and Train High), les tests devaient avoir lieu en condition de normoxie et les sujets devaient
être entraînés. Ces critères d’exclusion ont comme but d’isoler l’effet de l’hypoxie sur l’entraînement en RS.
19
Tableau 1-1. Effets comparés de l’entraînement de sprints répétés en hypoxie vs en normoxie sur les performances et les réponses
physiologiques
Études Sujets
(RSN, RSH)
Intervention
(Série x rép. x durées, repos intra, mode, repos inter-série)
Tests de RSA Performances Réponses physiologiques
Faiss et al., 2013 20, 20 cyclistes modérément entraînés 3 x 5 x 10-s, 20-s sur ergocycle, 5-min FiO2= 14,6%
8 séances sur 4 semaines
Boucle ouverte. 10-s 10-sprint, 20-10-s repo10-s. Ergocycle RSA: ↑ 4% Pmax ** et 42% du nombre de sprints**. Wingate: NS CLM 3-min: NS ↑ 18% [tHb]**,
11% [HHb] durant les sprints** et 7% [HHb] durant les récupérations* EMG, [Lac-] et VO 2max : NS Galvin et al., 2013 15, 15 joueurs de rugby hautement entraînés 1 x 10 x 6-s, 30-s sur tapis roulant, 5-min. FiO2= 13% 12 séances en 4 semaines 10 x 6-s, 30-s sur tapis roulant. RSA : NS Sprint 5, 10, 20-m : NS. YYIR1: ↑ 19% distance parcourue** ↑ 7% VO2 (p=0.06) VCO2, FC, VEet [Lac-] et [HHb] musculaire et cérébrale : NS Gatterer et al., 2014 5, 5 jeunes joueurs de soccer 3 x 5 x 10-s, 20-s en sprint navette, 5-min. FiO2= 14,8% 8 séances en 5 semaines. 6 x 40-m, 20-s en sprint navette. RSA : ↓ 47% régression linéaire de fatigue*. YYIR1: NS
[Lac-] lors du RSA et YYIR1 : NS
Faiss et al., 2015 8, 9 skieurs de fond hautement entraînés 4 x 5 x 10-s, 20-s en ski double poussée, 5-min. FiO2= 13,8%
6 séances sur 2 semaines
Boucle ouverte. 10-s sprint, 20-s repos. Double poussée. RSA : 55% du nombre de sprints**.
Pmoy et Pmax lors du test de
RSA : NS ↑ 142% [tHb]** et 252% [HHb]** EMG et [Lac-] : NS Brocherie et al., 2015 8,8 jeunes joueurs de soccer
5 x 4 x 5-s, 45-s sur tapis roulant ou en sprint navette, 5-min. FiO2= 14,3%
SpO2moy = 91-93%
10 séances en 5 semaines.
8 x 20-m, 20-s repos.
Sprint sur terrain.
RSASdec, moyen et
maximal : NS Temps sprint 10-m: ↓ 2% (ES 0,7; NS). CMJ, temps sprint 20 et 30-m : NS Kasai et al., 2015 16,16 joueuses de Lacross modérément entraînées 2 x 10 x 7-s, 30-s sur ergocycle, 10-min. FiO2= 14,5% SpO2 moy = 93% 8 séances en 4 semaines. 10 x 7-s, 30-s. Ergocycle. RSA : ↑ 4% Pmoy*et 4% Pmax* FI : NS VO2max et FC : NS
20 Goods et al., 2015 9,10 joueurs de football australien hautement entraînés 3 x 7 x 5-s, 15-35-s, sur ergocycle, 3-min. FiO2= 14,1% SpO2 moy = 89%
15 séances sur 5 semaines.
3 x 6 x 20-m, 25-s sur ergocycle et à la course, 3-min.
RSA ergocycle : NS RSA course : NS Test aérobique navette : NS [Lac-] et FC : NS Hamlin et al., 2017 9 joueurs de rugby bien entraînés 4 x 5 x s, 2s sur ergocyle, 5-min FiO2= 14,5%
6 séances sur 3 semaines.
8 x 20-m, 20-s. Sprint sur terrain.
RSA : ↓ 2% RSASdec 14
jours post entraînement (effet bénéfique possible (75%)). YYIR1: NS Montero & Lundby, 2017 15 cyclistes modérément entraînés. 4 x 5 x 10-s, 20-s sur ergocyle, 5-min. FiO2= 13,8% SpO2 moy = 94%
12 séances sur 3 semaines.
Boucle ouverte. 10-s 10-sprint, 20-10-s repo10-s. Ergocycle
RSA Pmoy. Pmax et nombre
de sprint : NS Test cardiovasculaire incrémental : NS
↑ 48% et 37% [tHb]* lors du test de RSA 5 jours post et 10 jours post. Brechbuhl et al., 2018 9, 9 joueurs de tennis bien entraînés. 4 x 5 6-s, 24-s sprint aller-retour à la course, 5-min. FiO2 = 14,5%
5 séances sur 2 semaines.
10 x 20-m sprint, 20-s. Sprint sur terrain. RSASdec : NS Test cardiovasculaire incrémental : ↑ 7%** temps avant épuisement.
Test cardiovasculaire
incrémental : ↑ 7% temps avant seuil lactate de 4mmol/L** et ↑ 12% temps avant le second seuil ventilatoire. VO2max, FC, VE et [Lac-] : NS Beard et al., 2019a 9, 10 joueurs de rugby hautement entraînés. 3 x 8 x 10-s, 20-s sur ergocycle, 2-min. FiO2 = 13,8%
4 séances sur 2 semaines.
6 x 10-s, 20-s. Sur ergocycle. RSA : ↑ 8% Pmax** et ↑ 5% Pmoy* RSASdec : NS Beard et al., 2019b 18, 18 joueurs de rugby hautement entraînés. 3 x 8 x 10-s, 20-s en ski double poussée, 2-min. FiO2 = 13,8%
4 séances sur 2 semaines.
6 x 10-s, 20-s. En double poussée.
RSA : ↑ 10% Pmax** et ↑
10% Pmoy**
RSASdec : NS
NS: changement non significatif, CLM: course contre la montre, RSA : capacité à répéter des sprints, RSASdec : score de détérioration de la performance lors d’un
test de RSA, EMG : électromyographie de surface, FC : fréquence cardiaque, FI : indice de fatigue, FiO2 : fraction d’O2 inspiré, [HHb]: concentration de
désoxyhémoglobine, [Lac-] : concentration de lactate, P
moy : puissance moyenne, Pmax : puissance maximale, rép. : répétition, [tHb]: concentration totale
d’hémoglobine, TOI : indice d’O2 du tissu, VE : ventilation, VCO2 : volume de dioxyde de carbone expiré, VO2max : volume de consommation maximale
d’oxygène, YYIR1 : Yo-Yo intermittent fitness test intermittent test level 1 et YYIR2: Yo-Yo intermittent fitness test intermittent test level 2. ** p< 0,01
21
Récemment, Brocherie et collaborateurs ont réalisé une méta-analyse à partir de 9 études concernant les effets du RSH sur les performances chez des athlètes de sports collectifs (Brocherie et al., 2017). D’après cette analyse combinée, le RSH, comparativement au groupe contrôle ayant subi le même entraînement de sprints, mais sans hypoxie ajoutée, aurait un effet bénéfique estimé de « faible à modéré » sur l’amélioration du temps moyen de sprints (taille d’effet (ES) : 0,46; 95% d’intervalle de confiance (IC) : -0,02 à 0,93) et du meilleur temps de sprint (ES : 0,31; IC : -0,27 à 0,89) lors des tests de RSA et du VO2max
(ES : 0,18; IC : -0,25 à 0,61) lors des tests incrémentaux aérobies. Cependant, ces résultats ne sont pas unanimes et certaines études ne rapportent aucune amélioration supplémentaire d’un entraînement RSH vs. en RSN sur la RSA (Goods et al., 2015; Montero & Lundby, 2017). En effets, ces dernières démontrent des gains de 10 à 17% pour la Pmoy et de 5 à
16% pour la Pmax en hypoxie comme en normoxie, et ces disparités pourraient être liées à
des différences méthodologiques ou d’analyse des données (Brocherie et al., 2017). De plus, l’entraînement RSH ne semble pas non plus apporter de bénéfices significatifs sur les performances aux tests aérobies comme le YYIR1 (Gatterer et al., 2014; Goods et al., 2015; Hamlin et al., 2017). Il semble donc au regard de l’ensemble de la littérature disponible que l’entraînement RSH soit pertinent dans un programme d’amélioration de la RSA, mais pas si l’on vise des adaptations de type aérobie.
Ces gains de performance pourraient être expliqués, entre autres, par une augmentation de la consommation d’O2 musculaire. En effet, les valeurs de [HHb] et de [tHb] au niveau du
muscle peuvent être deux à trois fois supérieures en RSH comparativement au RSN, exprimant une extraction et une perfusion sanguine améliorées (Faiss et al., 2015). D’autres études démontrent des adaptations similaires, mais de moins grandes amplitudes (Faiss et al., 2013; Montero & Lundby, 2017). Ces adaptations pourraient être possibles grâce à une vasodilatation compensatoire vers les muscles sollicités (Casey & Joyner, 2012). Ceci permettrait en un premier temps d’augmenter la perfusion sanguine et la disponibilité en O2 aux fibres de type II. Certains auteurs proposent une modification comportementale des
fibres de type II où ces fibres rapides agiraient plus comme des fibres de type I, soit des fibres moins fatigables (Cleland et al., 2012; Faiss et al., 2013, 2015). Ainsi, les fibres rapides seraient plus dépendantes du métabolisme oxydatif, surtout lors des périodes de récupération pour augmenter la disponibilité en O2 aux cellules musculaires et ainsi
22
bonifier la resynthèse de PCr. Lors des sprints, les voies anaérobies seraient alors davantage utilisées pour la production de l’énergie.
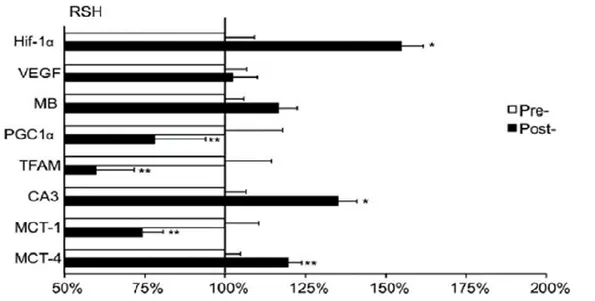
Cette interprétation est appuyée par l’analyse de biopsies musculaires prises avant et après 4 semaines d’entraînement en RSH (voir Figure 1-8) (Faiss et al., 2013). En effet, les auteurs ont mesuré une augmentation de l’expression des gènes responsables de la régulation du pH, soit l’enzyme anhydrase carbonique III (CA3) et du transporteur monocarboxylate musculaire IV (MCT4) ainsi qu’une diminution de l’expression des gènes responsables de la biogenèse mitochondriale dont le facteur de transcription mitochondriale A (TFAM) et le coactivateur 1α du récepteur gamma activé par des proliférateurs de peroxysomes (PGC-1α). De plus, l’étude démontre que l’activité de la citrate synthase est maintenue indiquant que la glycolyse aérobie n’a pas changé. Les auteurs interprètent ces résultats de biopsies par une augmentation de la contribution de la glycolyse anaérobie lors des sprints supportés par la glycolyse aérobie pour resynthétiser les substrats énergétiques lors des récupérations.
Si l’ajout d’un stress environnemental hypoxique à des entraînements de RS semble bénéfique à la performance chez des athlètes, une problématique importante persiste. En effet, l’entraînement RSH peut être compliqué à intégrer pour la plupart des équipes en raison des coûts financiers associés à l’équipement et de la logistique de mise en place. Ainsi, une approche plus accessible et pratique pour induire l’hypoxie à l’entraînement est nécessaire.
Figure 1-8 Changements dans l’expression des acides ribonucléiques messagers (mRNA) de certains