Étude originale
Application de marqueurs moléculaires
pour l’analyse de la diversité génétique
chez l’amandier (Prunus dulcis Mill.)
Karim Kadri1
Hager Snoussi1
Bennaceur M’Barek1
Abdallah Ben Abdallah1,2
1Laboratoire de biotechnologie et physiologie
végétale,
Institut national de la recherche agronomique de Tunisie (Inrat),
29, rue Hédi Karray, Ariana 2080, Tunisie
<[email protected]> <[email protected]> <[email protected]>
2UTF/NAM/004/NAM – FAO Project Date
Production Support Programme, P.O. Box 8697, Bachbrecht, Windhoek, Namibie <[email protected]>
Résumé
Plus de 100 cultivars d’amandier plantés en Tunisie pourraient constituer une source de variation génétique adaptée au milieu méditerranéen, à condition de réaliser une classification appropriée, identifiant les différences et similitudes entre les cultivars ainsi que les synonymies. Des essais reposant sur l’emploi de caractères moléculaires pour-raient complémenter les classifications traditionnelles fondées sur la morphologie. La technique RAPD (Random amplified polymorphic DNA) a été adoptée pour l’analyse de la diversité génétique des variétés tunisiennes d’amandier et de leurs relations avec d’autres variétés étrangères. À partir de 50 amorces testées, dix ont été choisies pour leur reproductibilité et leur polymorphisme élevé. Sur les 137 bandes détectées, 85 (soit 62 % de la totalité des bandes) se sont révélées polymorphes. Les corrélations entre les cultivars ont été évaluées à travers des analyses multivariées (analyses en composantes principales, ACP ; analyse factorielle des correspondances, AFC) utilisant les données de la RAPD. Les résultats obtenus indiquent une forte proximité génétique entre les cultivars, malgré des origines géographiques parfois très différentes. La technique RAPD s’est avérée être un outil efficace pour l’évaluation de l’organisation du germoplasme dans un objectif d’amélioration génétique.
Mots clés:polymorphime génétique ; RAPD ; Prunus dulcis. Thèmes:productions végétales ; métabolisme ; méthodes et outils.
Abstract
Using RAPD markers to assess the genetic diversity of almond trees (Prunus dulcis Mill.)
More than 100 almond cultivars grown in Tunisia could be a source of genetic variation adapted to the Mediterranean environment, provided that a proper classification identi-fying similar characteristics and synonymous cultivars is made. DNA-based assays could integrate traditional classifications based on morphology. RAPD markers (Randomly Amplified Polymorphic DNA) were used to analyse the genetic diversity of Tunisian almond cultivars and their relationships with foreign cultivars. Out of 50 primers tested, 10 were selected for their reproducibility and high polymorphism. Eighty-five out of 137 polymerase chain reaction fragments (62% of the total bands) were scored to be polymorphic. Cultivar relationships were estimated using multivaried analyses (FCA, PCA) based on RAPD data. The results indicate a strong genetic affinity among the cultivars, in spite of different geographic origins. RAPD seemed to be an effective tool to assess germplasm organization for further breeding programs.
Key words:genetic polymorphism; RAPD; Prunus dulcis. Subjects:vegetal productions; metabolism; tools and methods.
L’
amandier (Prunus dulcis Mill.) originaire des montagnes irano-afghanes, est un arbre d’une très grande importance économique et nutritionnelle. Cette espèce occupe, à l’échelle internationale, la troisième place après les agrumes et les pommes [1]. À l’heure actuelle, on assiste irrévocable-ment à un passage d’un agrosystème forestier caractérisé par une dynamique perpétuelle et une grande diversité phé-notypique à un système industriel repo-sant essentiellement sur une production monovariétale. Cela crée divers problè-mes qui ont un impact néfaste sur l’équili-bre environnemental. Par conséquent, la caractérisation des cultivars d’amandiers, l’évaluation du potentiel tunisien, ainsi que l’analyse de la diversité génétique des différents cultivars revêt un intérêt considérable pour les actions visant à la sauvegarde et à la conservation des res-sources génétiques de cette espèce. Dans cette optique, le présent travail a pour objet de caractériser les cultivars d’une collection nationale tunisienne d’amandiers en vue d’analyser la diversité génétique existante, en utilisant comme outil d’analyse du polymorphisme les marqueurs moléculaires de type RAPD (Random amplified polymorphic DNA) qui est une variante de la technique PCR (Polymerase chain reaction). Cette tech-nique, qui a été proposée simultanément par Welsh et McClelland [2] et Williams et al. [3], repose sur l’amplification parPCR de segment d’ADN en utilisant une amorce de petite taille (généralement
10 nucléotides), avec une séquence
nucléotidique arbitraire. L’avantage de cette technique par rapport aux autres est qu’elle n’utilise que quelques nanogram-mes d’ADN et n’emploie pas la radioacti-vité ; de plus, elle ne nécessite pas la connaissance préalable de la séquence de l’ADN matrice à amplifier. L’objectif à long terme est la conservation et l’amélio-ration du germoplasme des variétés tuni-siennes d’amandier.
Matériel et méthode
Matériel végétal
L’échantillonnage a été fait sur deux col-lections d’amandiers, l’une située au Cen-tre de Ettawos (Institut de l’olivier de Sfax, sud de la Tunisie) et l’autre apparte-nant au Groupement obligatoire des
viti-culteurs et producteurs de fruits
(GOVPF), situé à Grombalia au nord de la Tunisie. Les deux collections comportent des variétés locales et des variétés étran-gères. Treize variétés originaires des dif-férentes régions de Tunisie et réputées pour leur excellente qualité, leur produc-tivité élevée et leur résistance aux mala-dies cryptogamiques, ont été choisies pour cette étude (tableau 1).
Extraction de l’ADN total
L’ADN a été extrait à partir des feuilles. Nous avons opté pour le protocole décrit par Aitchitt et al. [4], qui est une combi-naison de trois méthodes d’extractions, faisant intervenir le CTAB (Cetyltrimethyl-ammonium bromide, bromure d’alkyltri-methylammonium). Ce même protocole a été adopté sur des cultivars tunisiens de palmier dattier pour une caractérisation au moyen de la technique AFLP (Ampli-fied fragment length polymorphism) [5].
Obtention des profils RAPD
Toutes les réactions RAPD ont été effec-tuées dans un milieu réactionnel de 25 lL
contenant 30 ng d’ADN génomique,
2,5 lL du tampon de l’enzyme de poly-mérisation 10X concentré (0,1M Tris HCl,
pH 9 ; 1,5 mm MgCl2; 0,5 M KCl),
20 pmol d’amorce, 2,5 lL de dNTPs (200 lM), 2 lL de MgCl2 (2,5 mM) et
1,5 U de Taq polymérase à 5 U/lL (Pro-mega, États-Unis). Les milieux réaction-nels contenus dans des tubes PCR ont
été placés dans un thermocycleur
(Biometra Uno II), programmé pour
effectuer une prédénaturation à 94 °C pendant 5 min, suivie de 45 cycles com-portant chacun une étape de dénatura-tion à 94 °C pendant 1 min, une étape
Tableau 1
.Les différentes variétés d’amandier sélectionnées pour cette étude.
Table 1. The different almond varieties selected for this study.Variétés Relations parentales Sources Origine
’Blanco’ Greffage GOVPFa Tunisie
’Heuch ben smail’ Greffage GOVPFa Tunisie
’Khouki’ Greffage GOVPFa Tunisie
’Achaak’ Semi Station Ettawsb Tunisie
’Ksontni’ Semi Station Ettawsb Tunisie
’Zaaf’ Semi Station Ettawsb Tunisie
’Nonpareil’ Semi Station Ettawsb États-Unis
(Californie)
’Mazzetto’ Greffage GOVPFa Lybie
’Ferragnes’ ’Cristomorto’ x ’Ai’ Station Ettawsb France (Inra, 1960)
’Ferraduel’ ’Cristomorto’ x ’Ai’ GOVPFa France (Inra, 1960)
’Lourane’ ’Ferragnès’ x ’Tuono’ Station Ettawsb France (Inra, 1978)
’Fakhfak’ Semi Station Ettawsb Tunisie
’Abiod Ras Jbel’ Greffage GOVPFa Tunisie
aGroupement obligatoire des viticulteurs et producteurs de fruits à Grombalia, Tunisie. b
d’hybridation des amorces à 34 °C pen-dant 1 min et une étape d’élongation à 72 °C pendant 2 minutes. La dernière étape de la réaction d’amplification consiste en une élongation finale à 72 °C pendant 10 min. La présence d’un éven-tuel polymorphisme a été détectée par migration des produits de PCR sur gel d’agarose à 1,8 % (migration 3 heures à 70 mA). Les gels ont été colorés au bro-mure d’éthidium et photographiés sous lumière UV. La taille des bandes obtenues a été estimée grâce au marqueur de poids moléculaire 1Kb DNA Ladder (Promega, États-Unis).
Méthodes numériques
et statistiques d’analyses
des résultats
Les différents profils RAPD obtenus ont été comparés entre eux. La lecture visuelle des gels a permis de tracer une matrice binaire 0/1 traduisant respective-ment l’absence/présence de bandes à chacun des allèles, pour chaque individu. À partir des matrices binaires obtenues, et en appliquant la formule de Nei et Li [6] relative au coefficient de Dice, des matri-ces de similarités génétiques entre les différentes variétés d’amandier ont été établies au moyen du programme SIM-QUAL (Similarity for Qualitative Data Program) dans le logiciel NTSYS-pc (The Numerical Taxonomy and multivariate Analysis System for Personnel Computer), version 2.0. Les analyses multivariées que nous avons utilisées étaient : d’une part, l’analyse en composantes principales (ACP) qui est fondée sur les corrélations pour transformer les variables de départ, dans notre cas les marqueurs RAPD, en variables synthétiques ou axes. Cette ana-lyse permet ainsi de définir les marqueurs qui contribuent le mieux à la description de la structuration de la variabilité et d’obtenir une représentation graphique de la projection des individus ou popula-tions selon leur ressemblance dans le plan des axes de l’ACP ; et d’autre part, l’analyse factorielle des correspondances (AFC) qui a été décrite comme étant une approche d’analyse appropriée dans le cas de données qualitatives binaires, per-mettant ainsi l’étude descriptive des don-nées [7]. L’AFC repose sur le calcul de la distance de v2à partir d’un tableau
dis-jonctif complet qui comporte l’ensemble des données portant sur des individus [8]. À partir des projections d’une AFC, il est
possible de déterminer quelles sont les variables qui interviennent dans la struc-turation des données et qui caractérisent les populations grâce à la superposition de la représentation graphique des varia-bles à celle des populations.
Ces analyses AFC ont été effectuées à l’aide du programme Digital Equipment Corporation – Open VMS AXP (version 6.2), contenu dans le logiciel SAS « Statis-tical Analysis System » [9]. L’ensemble de ces analyses a été réalisé au Centre inter-régional d’informatique et d’automatisme (Ciria) de la faculté des sciences de Tunis.
Résultats
Analyse numérique
des profils RAPD
Les différentes variétés d’amandier ont été analysées avec un total de 50 amor-ces, parmi lesquelles 11 n’ont donné aucun amplimère, quel que soit le culti-var analysé et 16 amorces ont généré des amplifications variables avec seulement
quelques-uns des cultivars. Sur les
13 amorces ayant produit des bandes reproductibles et intenses, 10 ont permis de générer un très grand nombre de bandes polymorphes (figure 1) et ont donc été utilisées dans le reste de l’étude (tableau 2).
Un total de 137 bandes reproductibles a été obtenu lors des amplifications utili-sant les 10 amorces retenues ; 85 bandes seulement étaient polymorphes (soit 62 % de la totalité des bandes), la moyenne des bandes polymorphes par génotype étant de 4,76 %. Environ 80 % des bandes présentaient une taille com-prise entre 0,5 et 3 Kb ; 12 % avaient une taille inférieure à 0,5 Kb et 8 % une taille supérieure à 3 Kb.
La moyenne des bandes reproductibles par amorce était de 13,7 bandes, avec 8,5 bandes reproductibles en moyenne
pour les bandes polymorphes. La
moyenne totale des bandes polymorphes par amorce était de 4 %.
Similarités génétiques
Les coefficients de similarités génétiques varient entre 0,52 et 0,81 avec une moyenne de 0,68. Les similarités les plus élevées sont observées entre les
combi-naisons variétales suivantes
(Nonpareil [N] – Achaak [A] : 0,8020), (Nonpareil [N] – Zaaf [Za] : 0,8085) et (Khouki [K] – Lourane [L] : 0,8135). Les
similarités les plus faibles ont été obte-nues avec les combinaisons (Mazetto [Mz] - Heuch Ben Samil [HB] : 0,5274) et (Mazetto [Mz] – Ferragnes [Fg] : 0,5454).
Analyse
en composantes principales
La matrice des données (matrice des simi-larités génétiques) a été traitée par une
analyse en composantes principales
(ACP) grâce au logiciel SAS [9]. Il en ressort que les trois composantes absor-bent 55,1 % de l’inertie totale, ce qui montre que les marqueurs de type RAPD peuvent être efficaces dans la description de la variabilité chez ces variétés. Les proportions ainsi que les variables qui définissent chaque axe sont représentées dans le tableau 3. La projection des varié-tés dans le plan défini par les axes 1 et 2 (figure 2), aboutit à des résultats qui sont plus ou moins similaires. Une aggloméra-tion de la presque totalité des variétés
forme un nuage à partir duquel
Blanco [B], Heuch ben smail [HB] et Mazetto [Mz], se différencient significati-vement des autres. En effet, les deux premières variétés corrèlent positivement
avec l’axe 1, alors que la variété
Mazetto [Mz] corrèle positivement avec l’axe 2. En tenant compte de la compo-sante 1, il est à noter que cette
représenta-tion permet d’opposer les variétés
Blanco [B], Houch ben smail [HB] aux variétés Ferragnes [Fg] et Ferraduelle [Fe], reflétant ainsi les faibles coefficients de similarité génétique obtenus entre ces variétés.
La représentation graphique de la projec-tion de l’ensemble des variétés dans les plans définis par l’axe 1 et l’axe 3 de l’ACP montre que la majorité des variétés impliquées ne se distinguent pas signifi-cativement les unes des autres et forment une agglomération qui se définit par la partie négative des deux axes. Cependant
les variétés Blanco [B], Heuch ben
smail [HB] et Mazetto [Mz], semblent se détacher de l’ensemble des variétés, par-ticulièrement par rapport à l’axe 1 qui absorbe le maximum de variabilité. La projection des variétés dans le plan défini par les axes 2-3 de cette analyse en composantes principales a montré que, dans la majorité des cas, les variétés sont regroupées entre elles avec un détache-ment assez significatif pour la variété Mazetto [Mz], qui corrèle positivement avec l’axe 2 et les variétés Ksontini [Ks] et Fakhfak [Fa] du côté positif de l’axe 3.
Analyse factorielle
des correspondances
Ce type d’analyse permet de caractériser des populations ou des groupes de popu-lation à l’aide des fréquences alléliques. C’est une technique qui consiste à faire une projection sur les mêmes axes des populations et des marqueurs RAPD et à déceler les principales bandes associées à la structure révélée. Dans ce travail, les 13 variétés constituent une population dans laquelle chaque variété correspond à un individu.
Le traitement des données par l’AFC utili-sant le programme SAS [9] a montré que les trois premières dimensions absorbent 60,6 % de l’inertie totale, ce qui repré-sente un taux d’absorption élevée et tra-duit l’efficacité de ces marqueurs dans la description de la variabilité génétique chez les variétés d’amandier étudiées.
Tableau 2
.Amorces RAPD sélectionnées, nombre de fragments
amplifiés et polymorphes obtenus pour chaque amorce.
Tableau 2. RAPD primer sequences used for the screening of the 13 accessions, scorable and polymorphic fragments obtained for each primer.
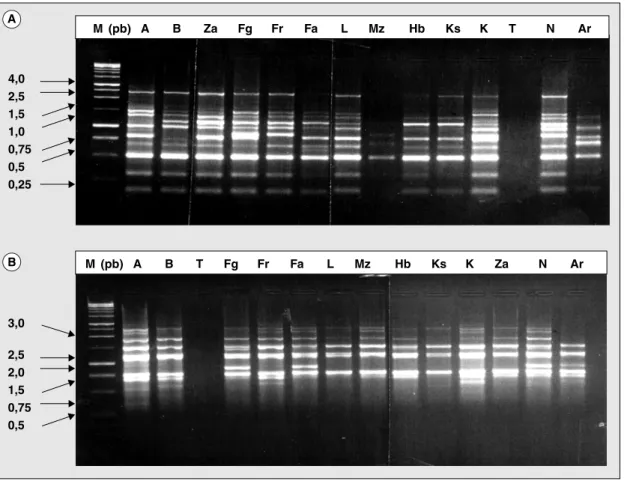
Amorces Séquence 5’ ---3’ Nombre de fragments amplifiés Nombre de fragments polymorphes OPA -03 AGTCAGCCAC 15 9 OPA -08 GTGACGTAGG 11 8 OPE - 06 AAGACCCCTC 14 9 OPE -07 AGATGCAGCC 15 10 OPP-08 ACATCGCCCA 15 9 OPB -01 GTTTCGCTCC 9 6 OPL -11 ACGATGAGCC 16 9 OPG –04 AGCGTGTCTG 14 8 OPI -18 TGCCCAGCCT 13 8 OPH - 04 CTGCATCGTG 15 9 Total 137 85 4,0 2,5 1,5 1,0 0,75 0,5 0,25 3,0 2,5 2,0 1,5 0,75 0,5 M (pb) A A B B Za Fg Fr Fa L Mz Hb Ks K T N Ar M (pb) A B T Fg Fr Fa L Mz Hb Ks K Za N Ar
Figure 1.Profil RAPD des variétés d’amandier obtenu avec l’amorce OPP08 (A) et OPE07 (B), (Operon Technologie Inc). Figure 1. RAPD pattern of Almond varieties obtained with primer OPP08 (A) and OPE07 (B), (Operon Technologie Inc).
Les lettres situées au-dessus de chaque bande correspondent aux variétés utilisées. A : Achaak ; B : Blanco ; Za : Zaaf ; Fg : Ferragnes ; Fr : Ferraduelle ; Fa : Fakhfak ; L : Lourane ; Mz : Mazetto ; Hb : Heuch ben smail ; Ks : Ksontini ; K : Koukhi ; N : Nompareil ; Ar : Abiod Ras Jbel.
Tableau 3
.Définition des axes et absorption de l’inertie de l’analyse en composantes
principales basée sur les marqueurs RAPD sur l’ensemble des variétés tunisiennes
d’amandier.
Table 3. Definition of the axes and absorption of the Principal Component Analysis inertia based on the RAPD markers for the whole set of the Tunisian almond-tree varieties.
Composante 1 2 3
% Inertie 22,84 17,43 14,83
% Cumulé 22,84 40,28 55,12
Définition des axes OPI18-06 (+ 0,1637) OPE07-14 (+ 0,2076) OPP08-01 (+ 0,2222) par les variables OPI18-07 (+ 0,1601) OPL11-01 (+ 0,2078) OPP08-04 (+ 0,2237) OPI18-11 (+ 0,1658) OPL11-02 (+ 0,2085) OPP08-06 (+ 0,2233) OPG04-01 (+ 0,1626) OPL11-05 (+ 0,2046) OPP08-08 (+ 0,2225) OPG04-03 (+ 0,1650) OPL11-09 (+ 0,2042) OPP08-09 (+ 0,2170) OPG04-05 (+ 0,1687) OPL11-14 (+ 0,2030) OPP08-13 (+ 0,2166) OPG04-06 (+ 0,1658) OPL11-15 (+ 0,2070) OPP08-14 (+ 0,2184) OPG04-09 (+ 0,1673) OPL11-16 (+ 0,2060)
OPG04-11 (+ 0,1667) OPG04-14 (+ 0,1684)
Les chiffres entre parenthèses représentent la contribution de chaque variable dans la définition de l’axe.
0 + 5 + 10 + 15 - 2,5 - 5,0 17,5 15,0 12,5 10,0 7,5 5,0 2,5 0 - 10 - 5 • K • Za • Fa • A • N • Ks • Ar • L • Fe • Fg •Mz • HB • B Axe 2 (17,43)%) Axe 1 (22,84)%)
Figure 2.Représentation graphique de la distribution spatiale de 13 variétés d’amandier dans les plans 1-2 d’une analyse en composantes principales (ACP) (progiciel SAS, SAS Institute Inc., États-Unis), réalisée à partir des marqueurs RAPD.
Figure 2. Graphic representation of the spatial distribution of 13 almond varieties in plans 1-2 resulting from a Principal Component Analysis (SAS package, SAS Institute Inc., USA) based on RAPD markers.
A : Achaak ; B : Blanco ; Za : Zaaf ; Fg : Ferragnes ; Fr : Ferraduelle ; Fa : Fakhfak ; L : Lourane ; Mz : Mazetto ; HB : Heuch ben smail ; Ks : Ksontini ; K : Koukhi ; N : Nompareil ; Ar : Abiod Ras Jbel.
La définition et l’absorption de la variabi-lité par les dimensions (variables synthéti-ques) de l’AFC sont résumées dans le tableau 4.
Discussion
Plusieurs facteurs peuvent affecter le nombre de bandes obtenues à l’issue des amplifications, à savoir : la variété étu-diée, le nombre de génotypes comparés, les séquences des amorces ainsi que des variations mineures dans les protocoles d’amplification. Ce dernier facteur affecte généralement la reproductibilité. Par comparaison avec d’autres travaux por-tant sur l’étude du polymorphisme molé-culaire par le biais des marqueurs RAPD, la moyenne des bandes polymorphes par amorce était de 4 % chez 17 cultivars d’amandier italiens [10], 4,7 % chez 15 cultivars d’amandier portugais [11], 5 % chez 25 variétés de pommiers [12].
Nos résultats, qui révèlent un niveau de polymorphisme du même ordre que celui obtenu au sein d’autres variétés d’aman-dier dans d’autres pays (62 % chez les 13 cultivars tunisiens d’amandier), suggè-rent que les marqueurs de type RAPD sont potentiellement utiles pour la carac-térisation des variétés d’amandier en Tunisie.
Les coefficients de similarités génétiques présentent un intervalle qui traduit une forte convergence génétique et montre que les variétés étudiées constituent des groupes étroitement liés. En fait, ces résultats sont presque similaires à ceux de Martins et al. [11] qui obtenaient un inter-valle de similarités génétiques variant entre 0,65 et 0,97 (moyenne 0,73) chez 15 cultivars d’amandier d’origine portu-gaise, en utilisant le même coefficient de similarité de Dice (SD). De même, les
17 cultivars d’amandier d’origine ita-lienne analysés par Resta et al. [10] pré-sentaient des similarités génétiques variant entre 0,64 et 0,94. Bartolozzi et al. [13] ont également obtenu des similarités
variant entre 0,62 et 1,0 chez 20 cultivars d’amandiers d’origine américaine. Tous ces résultats militent en faveur du fait que les cultivars d’amandier sont génétiquement proches, étant donné que les indices de similarités génétiques sont pour leur majorité proches de 1.
Le résultat trouvé chez les variétés qui présentent les similarités les plus élevées suggère que ces dernières constituent des groupes étroitement liés. Cette similarité pourrait être due au mode de multiplica-tion par semis suivi par les agriculteurs de la région (Centre Ettawos, Sfax), qui favo-rise l’homogénéité génétique de l’ensem-ble de ces variétés (autochtones et étran-gères), en dépit de leur origine. Le résultat trouvé chez les variétés qui pré-sentent les similarités les plus faibles tra-duit une certaine divergence entre ces variétés. Cette divergence peut être expli-quée par le mode de multiplication par greffage adopté par les agriculteurs de la région du nord (GOPVF de Grombalia).
Tableau 4
.Définition et absorption de la variabilité par les dimensions (variables synthétiques)
de l’analyse factorielle des correspondances à partir de la matrice des données.
Table 4. Definition and absorption of the variability by the measurements (synthetic variables) of the Factorial Correspondance Analysis from the data matrix.
Dimension de l’AFC 1 2 3
% Inertie 23,24 19,79 17,61
% Cumulé 23,24 43,04 60,64
Variétés contribuant à la définition
des dimensions de l’AFC (variables synthétiques)
Achaak (+ 0,251) Mazetto (- 1,429) Ksontini (- 1,135) Blanco (- 1,091) Abiod.de Ras.Jbel (+ 1,307) Fakhfak (+ 0,212)
Ferraduelle (+ 0,090) Ferragnes (+ 0,614) Houch Ben Smail (- 1,078)
Khouki (+ 0,195) Lourane (+ 0,300) Nonpareil (+ 0,228) Zaaf (+ 0,177) Marqueurs RAPD contribuant à la définition des dimensions de l’AFC (variables synthétiques)
OPI18-01 (- 1,240) OPP08 -04 (+ 1,602) OPP08-06 (+ 1,228)
OPI18-06 (- 1,160) OPL11-11 (- 1,379) OPP08-08 (+ 1,245)
OPI18-13 (- 1,219) OPL11-12 (- 1,180) OPB01-01 (+ 1,379)
OPG04-01 (- 1,315) OPP08-14 (+ 1,336) OPB01-02 (+ 1,282)
OPG04-05 (- 1,501) OPL11-03 (- 1,293) OPB01-03 (+ 1,829)
OPG04-07 (- 1,209) OPL11-05 (- 1,305) OPB01-04 (+ 1,534)
OPG04-08 (- 1,140) OPL11-10 (- 1,616) OPB01-05 (+ 1,710)
OPG04-14 (- 1,335) OPL11-14 (- 1,861) OPP08-13 (- 1,258)
OPG04-02 (- 1,174) OPL11-16 (- 2,117) OPE06-07 (+ 1,240)
Analyse multivariée
Les résultats montrés par l’ACP suggèrent que l’axe 1 est la composante qui permet de distinguer significativement les varié-tés Blanco [B] et Heuch ben smail [HB]. Ces variétés semblent être caractérisées par les marqueurs RAPD corrélant avec la composante 1. En revanche, la variété
Mazetto [Mz] serait caractérisée par les marqueurs qui définissent la compo-sante 2. L’axe 3 a permis la discrimination significative des variétés Ksontini [Ks] et Fakhfak [Fa] dans sa partie positive. Notons que la variété Mazetto [Mz], en se détachant légèrement des autres selon toutes les projections analysées, semble
être caractérisée par les marqueurs RAPD qui définissent les trois axes de cette ACP. L’analyse en composantes principales, réalisée à partir des fréquences de la présence des fragments RAPD chez diffé-rentes variétés, montre une distinction qui est plus ou moins importante entre ces variétés. Elle témoigne notamment de
Axe 2 (19,79 %) 2 1,5 1 0,5 0 - 0,5 - 1,0 - 1,5 - 1,5 - 1,0 - 0,5 0 + 0,5 + 1,0 + 1,5 - 2 Axe 1 (23,24 %) ♦H5 ♦H14 ♦H2 •F g ♦H1 ♦H12 ♦E 3 ♦E 13 ♦E 1 ♦E 3 ♦H8 ♦E 7 ♦H6 •H B ♦B 1 ♦E 12 ♦E 5 ♦H3 ♦ B 7 ♦E 2 • Z a ♦E 6 ♦E 8 ♦E 9 ♦D6 ♦D14 ♦D1 ♦D3 •K s ♦D5 ♦D7 ♦D13 ♦D2 ♦D8 ♦D6 ♦D15 ♦F1 ♦F6 ♦F9 ♦F7 •F a ♦F2 ♦J7 ♦J5♦J2 ♦F8 ♦J1•A r ♦F4 ♦C8 ♦C1 ♦J12 ♦J7 ♦A 1 ♦C15 •A ♦J6 ♦A 5 ♦I 3 ♦I 7 ♦C2 ♦A 2 •K ♦C1 ♦J10 ♦J3 ♦I 2 •N ♦A 15 ♦B 8 ♦B 9 ♦B 13 ♦A 12 ♦B 11 •B ♦I 1 ♦B 6 ♦B 2 ♦I 8 ♦I 5 ♦A 5 ♦A 1 •F e ♦B 3 ♦C6 ♦G6 ♦B 2 ♦G9 •L ♦ G13 ♦G8 ♦G12 •M z ♦G16 ♦G10 ♦ G7 ♦G16
Figure 3. Représentation graphique de la projection des marqueurs RAPD (♦) et des variétés d’amandier (•) dans le plan défini par les axes 1 et 2 d’une analyse factorielle des correspondances (AFC), (progiciel SAS, SAS Institute Inc., États-Unis).
Figure 3. Graphic representation of the RAPD markers projection (♦) and that of the Tunisian almond varieties (•) in the plan defined by the axes 1 and 2 resulting from a Factorial Correspondances Analysis (SAS package, SAS Institute Inc., USA) based on RAPD markers.
A : Achaak ; B : Blanco ; Za : Zaaf ; Fg : Ferragnes ; Fr : Ferraduelle ; Fa : Fakhfak ; L : Lourane ; Mz : Mazetto ; HB : Heuch ben smail ; Ks : Ksontini ; K : Koukhi ; N : Nompareil ; Ar : Abiod Ras Jbel.
convergences génotypiques importantes et traduit l’existence de flux de gènes élevés entre les variétés Achaak [A] et Zaaf [Za]. La variété Mazetto [Mz] fait exception : elle paraît relativement éloi-gnée dans les trois plans et montre une divergence génétique assez importante. Ces résultats confirment l’hypothèse émise par Resta et al. [10] concernant l’absence de l’effet d’éloignement géogra-phique dans la structuration de la variabi-lité. En effet, la majorité des variétés se regroupent pour former un agglomérat en dépit de leur éloignement géographique. La représentation graphique de la projec-tion des variétés et des marqueurs dans les plans définis par les dimensions 1, 2 et 3 prises deux à deux, montre des agglomérats de l’ensemble des données (variétés et marqueurs) de part et d’autre des axes qui définissent chaque plan de l’AFC.
Par ailleurs, la projection des individus selon le plan de l’AFC défini par les axes 1 et 2 (figure 3), montre la séparation des
variétés Ferragnes [Fg], Houch ben
smail [HB] et Zaaf [Za] des autres variétés selon la partie négative de l’axe 1. Cepen-dant, des superpositions des marqueurs avec ces variétés sont observées. C’est le cas des marqueurs OPA08-01, OPA08-07, OPG04-08, OPG04-06 et OPG04-03 qui caractérisent les variétés Heuch ben smail [HB], d’une part, et des marqueurs OPI18-12, OPI18-2, OPI18-05, OPI18-06 et OPI18-09 qui définissent la variété Zaaf [Za], d’autre part. Les variables
OPG04-01, OPG04-02, OPG04-14,
OPG04-12 et OPI18-03 corrèlent négati-vement avec la variété Ferragnes [Fg]. La structuration observée avec l’AFC confirme celle obtenue avec l’ACP. Notons que des résultats similaires ont été obtenus par Martins et al. [11], qui ont analysé avec des marqueurs RAPD la diversité génétique de cultivars d’aman-dier issus de différentes origines géogra-phiques. Dans leurs travaux, ces auteurs
ont pu démontrer que les cultivars testés constituent des groupes d’affinités généti-ques indépendamment de leur origine géographique et de la qualité de leurs fruits.
Conclusion
Au cours de cette étude nous avons utilisé des outils moléculaires pour caractériser des variétés tunisiennes d’amandier. L’analyse en composantes principales (ACP) réalisée sur les fréquences d’appa-rition des bandes RAPD chez les variétés étudiées, a montré que la majorité des variétés paraissent relativement proches. Ces résultats seraient en faveur de l’hypo-thèse stipulant l’existence d’un flux géni-que entre ces variétés et reflètent les ressemblances morphologiques.
La structuration de la variabilité génétique a également été étudiée par l’analyse fac-torielle des correspondances (AFC). Cette analyse a abouti à des résultats similaires à ceux obtenus par l’ACP. Elle a montré que la majorité des variétés sont chevau-chantes à quelques exceptions près. L’extension de l’étude à d’autres variétés ou autres génotypes et l’utilisation d’un plus grand nombre d’amorces permet-traient de confirmer et de valider la struc-turation obtenue.
L’ensemble des résultats issus de cette analyse montre que la technique d’ampli-fication par RAPD constitue une appro-che informative de la diversité génétique chez les variétés tunisiennes d’amandier. Les résultats obtenus, bien que prélimi-naires, apportent des éléments d’informa-tions ouvrant des pistes pour une explo-ration plus approfondie de la diversité génétique et des relations entre les culti-vars tunisiens d’amandier. Ces données pourraient permettre d’explorer les rela-tions phylogénétiques entre les variétés sans pour autant refléter les processus évolutifs chez cette espèce.■
Références
1. Grasselly C, Duval H. L’amandier. Paris : Centre technique interprofessionnel des fruits et legumes (CTIFL), 1997.
2. Welsh J, Mc Celland M. Fingerprinting genomes using PCR with arbitrary primers.
Nucleic Acids Res 1990 ; 18 : 7213-8.
3. Williams JGK, Kubelik AR, Livac KJ, Rafalski JA, Tinge SV. DNA polymorphisms amplifies by arbitrary primers are useful as genetic markers. Nucleic Acids Res 1990 ; 22 : 6531-5.
4. Aichitt M, Aisworth CC, Thangavelu M. A rapid and efficient method for the extraction of total DNA from mature leaves of the data palm (Phoenix dactylifera L.). Plant Mol Biol Rep 1993 ; 11 : 317-9.
5. Snoussi H. Génotypage d’individus
apomic-tiques chez le palmier datier (Phoenix
dactyli-fera L.) au moyen de la technique AFLP. DEA, faculté des sciences agronomiques de Gem-bloux, Belgique, 2000
6. Nei M, Li WH. Mathematical model for stu-dying genetic variation in terms of restriction endonucleases. Proc Natl Acad Sci USA 1979 ; 76 : 5269-73.
7. Boursiquot JM, Faber MP, Blachier O, Truel P. Utilisation et traitement statistique d’un fichier ampélographique. Agro 1987 ; 7 : 13-20. 8. Jumbo M. Exploration informatique de
don-nées. Paris : Ed Dunod, 1989.
9. SAS Institute. SAS user’s guide. Version 6, Fourth Edition. Cary (North Carolina) : SAS Institute Inc., 1990.
10. Resta P, Ferrara G, Fanizza G, Palasciano A, Godini A. Random amplified DNA polymor-phism of almond (Amygdalus communis L.) cultivars in Apulia. Proc : “X GREMPA semi-nar”, Meknès, Morocco, 14-17 /10/1996. Cahier
Opt Med 1998 ; 33 : 11-8.
11. Martins M, Tenreiro R, Oliveira MM. Gene-tic relatedness of Portuguese almond cultivars assessed by RAPD and ISSR markers. Plant
Cell Rep 2003 ; 22 : 71-8.
12. Landry B, Li R, Cheung W, Granger R. Phy-logeny analysis of 25 apple rootstocks using RAPD markers and tactical gene tagging.
Theor Appl Genet 1994 ; 89 : 847-52.
13. Bartolozzi N. Random amplified DNA poly-morphisms in Amygdalus communis L. Second International Symposium on Pista-chios and Almond, August 24-291997, Davis, USA. Acta Hortic 1998 ; (470) : 82–90.