HAL Id: hal-02810771
https://hal.inrae.fr/hal-02810771
Submitted on 6 Jun 2020HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Accroissement annuel des arbres en épaisseur : effet de
la méthode de mesure.
Rémi Blazin
To cite this version:
Rémi Blazin. Accroissement annuel des arbres en épaisseur : effet de la méthode de mesure.. [Stage] IUT d’Avignon. Université d’Avignon et des Pays de Vaucluse (UAPV), Avignon, FRA. 2011, 34 p. �hal-02810771�
RAPPORT DE STAGE
Accroissement annuel des arbres en épaisseur :
effet de la méthode de mesure.
Rémy BLAZIN
IUT d'Avignon – DUT Génie Biologique option Agronomie
Tuteur : François MESLEARDMaître de stage : François COURBET 26 Avril – 1 Juillet 2011
INRA PACA Domaine Saint-Paul Site Agroparc 84914 Avignon Cedex 9 +33 (0)4 32 72 20 00 IUT d'Avignon Université d'Avignon et des Pays du Vaucluse
377 chemin des Meinajaries, BP 1207, 84911 Avignon
cedex 09 +33 (0)4 90 84 14 00
2
Remerciements
Je voudrais tout d’abord remercier François LEFEVRE, directeur de l’Unité de Recherche « Ecologie des Forêts Méditerranéennes » au sein de laquelle j’effectue mon stage.
Je remercie particulièrement François COURBET sans qui je n’aurais pas pu faire ce stage. Je le remercie pour toute l’énergie qu’il a consacrée pour le bon déroulement de mon séjour à l’INRA. Mais aussi pour avoir pris autant sur son temps pour éclaircir mes zones d’ombres et de m’avoir aidé pour la rédaction de ce rapport.
Je remercie également Joannès GUILLEMOT, diplômé de la FIF et désormais étudiant en Master II, pour toutes les connaissances qu’il m’a apportées sur le domaine forestier.
Un merci tout particulier à Marianne CORREARD, pour le soutient et ses précieux conseils apportés lors de la rédaction du rapport, mais aussi pour ses indications sur le monde forestier et ses formations.
Merci à Philippe DREYFUS, Thomas BOIVIN, Aurore BONTEMPS, Marie-Joe KARAM, Marion GILLMANN, William BRUNETTO, Hadrien LALAGÜE, et Nicolas MARIOTTE, pour leur soutien et leurs encouragements, mais aussi pour leurs conseils et leurs remarques pour préparer ma soutenance de stage.
Enfin je remercie toute l’équipe EFDC et l’URFM en général pour leur accueil, leur sourire et leur bonne humeur.
3
Sommaire
1. Introduction : ... 1
1.1 Contexte et cadre de l'étude. ... 1
1.2 Objectif de l'étude. ... 2
2. Matériels et Méthodes ... 3
2.1. Le site d'étude. ... 3
2.1.1. Le dispositif expérimental de « l'Arigier ». ... 3
2.1.2. La parcelle étudiée. ... 4
2.2. Matériel biologique : le cèdre de l'Atlas. ... 4
2.3. Le protocole expérimental. ... 4
2.3.1. Mesures sur le terrain. ... 4
2.3.2. Mesures en laboratoire. ... 5
2.4. Mise en forme et prétraitement des données. ... 7
2.4.1. Vérification de la chronologie : inter-datation. ... 7
2.4.2. Les modèles d'épaisseur d'écorce et de retrait. ... 7
2.4.3. Calcul des accroissements en surface terrière. ... 9
3. Résultats ... 11
3.1. L'inter-datation. ... 11
3.1.1. Comparaison des largeurs de cerne entre arbres. ... 11
3.1.2. Comparaison des accroissements en surface terrière laboratoire et terrain. ... 12
3.2. Comparaison individuelle entre les deux méthodes de mesure. ... 13
3.2.1. La régression. ... 13
3.2.2. Tests sur les résidus du modèle : accstt= f (accstlab). ... 14
3.3. Comparaison annuelle entre les deux méthodes de mesure. ... 15
3.3.1. Observations graphique des données. ... 15
3.3.2. Test sur les résidus du modèle accstt= f (accstlab) en fonction de l'année... 17
3.3.3 Coefficient de synchronisation (SR) ... 18
4. Discussion ... 19
4.1.Synthèse sur l'analyse des résultats. ... 19
4.1.1.L'inter-datation. ... 19
4.1.2. Analyse individuelle. ... 20
4.1.3. Analyse annuelle. ... 21
4
4.2.Les points critiques de la méthode ... 22
4.2.1 Les mesures terrains ... 22
4.2.2 Les mesures en laboratoire ... 23
4.2.3 Les modèles ... 23 4.3.Perspectives d'améliorations ... 24 4.3.1. Mesures terrains ... 24 4.3.2. Mesures en laboratoires ... 24 Conclusion : ... 25 BIBLIOGRAPHIE ... 26 ANNEXES ... 28
1
1. Introduction :
1.1 Contexte et cadre de l'étude.
Qui de nos jours n'a pas entendu parler du réchauffement climatique, de ses causes et de ses impacts multiples? C'est un sujet d'actualité qui revient régulièrement et de plus en plus fréquemment. Des scientifiques tentent de le caractériser notamment en mesurant et en surveillant l'évolution des températures annuelles par exemple. Des premières observations ont pu alors être faites en particulier sur le déplacement des isothermes vers le nord : on remarque par exemple que la température moyenne annuelle à Valence est déjà celle qu'on observait en Avignon il y a 25 ans (Séguin 2011 communication personnelle). La région méditerranéenne est déjà un bon observatoire qui préfigure les futures conditions climatiques des régions plus septentrionales.
Cependant des effets résultant de ce changement climatique sont déjà perceptibles sur l'agriculture et la forêt en France, pour exemple la floraison précoce des arbres
fruitiers et des vignes (Bréda et al. 2011). Ces premières conséquences amènent à se demander quels sont les risques
pour la forêt face aux modifications climatiques. Comment vont réagir les essences végétales présentes ? Vont-elles s'adapter, migrer ou sont-elles appelées à disparaître ? Autant de questions qui laissent les forestiers dans l'incertitude quant aux essences à implanter et aux techniques sylvicoles à appliquer.
C'est en faisant ce constat qu'est né il y a deux ans, le réseau mixte technologique AFORCE. Ce réseau, comprenant 12 partenaires qui appartiennent aux domaines de la recherche, du développement, de la gestion et de la formation, a pour objectif la coordination des actions visant à adapter les forêts au changement climatique pour fournir aux gestionnaires forestiers des outils d'aide à la décision. Résistant à la sécheresse, peu sujet aux gelées printanières et pourvu d'une bonne croissance et d'une bonne valeur économique, le cèdre de l'Atlas s'est présenté comme le candidat logique pour parer au réchauffement climatique (Riou-Nivert 2007). C'est donc autour de cet arbre que de nombreux projets de recherches sont concentrés.
L'Unité de Recherches des Forêts Méditerranéennes de l'INRA d'Avignon travaille sur l'un des projets du RMT. Ce dernier vise à rassembler les connaissances disponibles pour aider les gestionnaires dans leur décision d'installer ou non du cèdre et de choisir pour les peuplements existants et à venir, les itinéraires techniques les plus adaptés pour minimiser le risque face au changement climatique. Toutes ces informations seront au final rassemblées sur une brochure à la disposition de tous, combinées avec un bilan économique des sylvicultures préconisées.
Ce projet se décompose en deux volets. Un volet développement, chargé de rassembler les connaissances disponibles sur l'espèce et sa conduite en plantation,
2 afin d'amener des solutions efficaces, et viable économiquement pour les gestionnaires de cédraie. Le volet recherche, est celui dans lequel s'inscrit ce rapport de stage. Ce volet prévoit l'étude des effets du climat et de la sylviculture sur la croissance des arbres, en faisant la part respective des différents facteurs ; densité du peuplement, climat, et de leur interaction précisant ainsi l'effet de la compétition sur la relation entre la croissance et le climat. Différentes situations de concurrences contrastées, obtenues par des éclaircies plus ou moins fortes, ont été testées dans un dispositif expérimental sur le versant sud du mont Ventoux dans le Vaucluse (84). Ce travail fait l'objet d'un stage de master II mené parallèlement à notre travail.
1.2 Objectif de l'étude.
Ce stage de master prévoit d'analyser des mesures individuelles de croissance, obtenues par arbre suite aux inventaires des circonférences, réalisés tous les ans dans le dispositif. Or l'étude de la croissance annuelle se pratique habituellement en dendrochronologie* ou en dendroclimatologie sur des carottes ou des rondelles prélevées sur les arbres et sur lesquelles on mesure la largeur des cernes annuels successifs (Pardé 1988).
L'objectif de la présente étude est de préciser si les accroissements annuels mesurés sur le terrain sont bien représentatifs de ceux mesurés a posteriori sur rondelles. Quelles relations existent entre ces deux types de mesures? L'objectif de ce stage est donc méthodologique. Il n'existe peu voir pas d'étude, ou d'ouvrage qui ait abordé ce type de travaux. Cela est en parti dû au fait qu'il n'est pas fréquent d'utiliser deux techniques différentes pour réaliser un même travail. Nous avons profité d'une éclaircie récente dans le dispositif expérimental pour comparer les résultats de mesures effectuées avec les deux techniques sur un échantillon d'arbres abattus. Les résultats peuvent avoir une valeur générique évidente pour d'autres espèces que le cèdre, et permettront de valider les mesures analyser par Joannès Guillemot étudiant en Master II.
*Dendrochronologie : La dendrochronologie est une science qui repose sur la mesure des largeurs de cernes annuels de croissance des arbres et sur leur datation précise
3
2. Matériel et Méthodes
2.1. Le site d'étude.
2.1.1. Le dispositif expérimental de « l'Arigier ».
Le cadre géographique de cette étude se situe en région Provence-Alpes-Côte-d'Azur, dans le département du Vaucluse sur la commune de Sault. La zone mise à disposition pour les expérimentations est sur le versant sud du mont Ventoux dans la forêt domaniale du Ventouret au lieu-dit l'Arigier. La pente est d'environ 6-8°, pour une altitude moyenne de 1170 mètres. Le dispositif a été installé en 1991 dans une plantation équienne de 27 ans réalisée à 2700 tiges par hectare. Il comprend 5 placettes qui se distinguent les unes des autres par leur traitement sylvicole : intensité et précocité des éclaircies (cf figure 1).
Figure 1 : Photo aérienne du dispositif expérimental de l'Arigier. Forêt domaniale du Ventouret.
Sault
Mt Ventoux Sault
4 2.1.2. La parcelle étudiée.
Les arbres sur lesquels ont été réalisées les mesures faites durant le stage proviennent de la placette 1. Cette placette a subi un traitement qualifié de faible: une première éclaircie a eu lieu en 1991 au profit de 250 arbres d'avenir par ha, en enlevant les 4 ou 5 voisins les plus gênants, ramenant la densité à 1800 tiges ha. Puis en 1998 une seconde éclaircie a réduit la densité à 1200 tiges par ha. C'est lors de l'éclaircie de 2008, qui a ramené la densité à 700 tiges par ha, que les arbres de cette étude ont été abattus. Ils étaient donc âgés de de 44 ans depuis la graine.
2.2. Matériel biologique : le cèdre de l'Atlas.
Cedrus atlantica est un arbre pouvant atteindre 40 mètres de hauteur. Le
genre Cedrus appartient à la famille des Pinacées. C'est un arbre d'une très grande longévité, pouvant aller jusqu'à 800 ans. Il affectionne les climats lumineux aux étés secs. Dans sa jeunesse c'est une espèce de demi-ombre très sensible à la compétition avec la strate herbacée. Il supporte les fortes sécheresses estivales et tolère les hivers froids tant que la température ne descend pas en dessous de -20°C. Il est donc parfaitement adapté à la région méditerranéenne au-dessus toutefois de 400 m d'altitude. Le système racinaire du cèdre est pourvu d'une puissante racine pivot, qui lui permet de se développer dans des sols superficiels calcaires si la roche est fissurée. Il donne aussi d'excellents résultats dans des sols meubles et profonds. En revanche il redoute les sols argileux limoneux compacts ou présentant des traces d'hydromorphie. Son bois présente des qualités mécaniques élevées. Son duramen odorant est de couleur rouge. Imputrescible, on l'utilise donc comme bois de charpente, en ébénisterie, mais aussi en menuiserie intérieure et extérieure.
L'aire naturelle du cèdre de l'Atlas se situe en Afrique du nord où il vit généralement dans les zones montagneuses à une altitude comprise entre 1500 et 2500 mètres. On le retrouve aujourd'hui dans d'autres régions du monde tel que le sud de la France. Son introduction sur les pentes dénudées du Mont Ventoux et celles du petit Luberon remonte à 1860 suite aux lois de Restauration des Terrains en Montagne. La réussite spectaculaire de ces reboisements procure au département du Vaucluse le privilège de posséder deux des plus belles cédraies de France (Girerd 1991). De plus l’extension naturelle de cette essence est telle que l'on peut actuellement rencontrer des cèdres un peu partout dans les massifs montagneux vauclusiens.
2.3. Le protocole expérimental. 2.3.1. Mesures sur le terrain.
L'étude a porté sur un échantillon de 21 arbres répartis dans toutes les classes de circonférence de la placette.
5 circonférence des arbres au mm près à l'aide d'un ruban souple tendu sans excès autour de la section (Rondeux 1993). L'outil employé est un « ruban métrique », gradué en centimètres et en millimètres, à trame métallique, afin d'éviter toute déformation. (Forêts d'Auvergne fiche technique n°38). La mesure se fait à hauteur fixe dite à « hauteur d'homme », proche de 1,30 m et repérée à la peinture en 1989 en s'éloignant du niveau d'insertion des branches pour éviter les déformations. Jusqu'en 2002 inclus, une double mesure a été faite sur les arbres (2 inventaires successifs). Lorsque la deuxième mesure différait de 2 mm ou moins de la première, la moyenne des deux mesures a été utilisée. Dans le cas contraire une troisième mesure a été effectuée et utilisée. A partir de 2003, pour des raisons économiques, il n'y a eu plus qu'un seul inventaire annuel complet effectué. Cet inventaire était ensuite vérifié au bureau pour détecter d'éventuelles erreurs de mesures au moyen de l'établissement de graphiques et de relations établies entre l'accroissement annuel et :
- l'accroissement annuel de l'année précédente - la circonférence de l'arbre de l'année précédente
Les arbres s'écartant trop du nuage de points ou des relations établies, ont fait l'objet d'une remesure sur le terrain qui a été utilisée ensuite.
Les mesures ont toujours été faites en arrêt de végétation d'octobre à mars. En effet il est nécessaire que les arbres ne soient pas dans une période de production de bois pour effectuer les mesures.
2.3.2. Mesures en laboratoire.
Les mesures en laboratoire ont été effectuées sur les rondelles des 21 arbres issus de la placette 1 mesurés sur le terrain. Dans le but de travailler avec le même matériel que sur le terrain, ces rondelles ont toutes été prélevées au niveau du trait de mesure. Ce trait, situé au Nord, a permis d'orienter les rondelles.
Suite à leur prélèvement, les rondelles ont été poncées sur les deux faces, afin de rendre leurs surfaces planes et de faciliter la vision des cernes et donc leur mesure. Après cette préparation les rondelles sont stockées en chambre froide pour limiter leur dessèchement et leur fendillement. Malgré tout, les rondelles subissent quand même une variation de dimension, c’est le retrait du bois.
Leur circonférence a donc été remesurée avant scannage. La technique en laboratoire consiste à mesurer plusieurs rayons, allant du centre de la moelle au bord extérieur de l'écorce. Sur chaque rondelle ont été mesurés 8 rayons, numérotés de 1 à 8 dans le sens des aiguilles d'une montre à partir du Nord, sur la face supérieure (cf figure 2).
6 Figure 2: Schéma descriptif de l'orientation des 8 rayons sur les rondelles.
Un cerne correspond à la croissance annuelle de l'arbre. Il est composé de deux parties :
-le bois de printemps ou bois initial constitué, chez les conifères, de grosses cellules à paroi fine appelées trachéides. Peu dense il est de couleur claire, il se forme du début du mois de juin à la fin du mois de juillet. Néanmoins cette période est variable selon le climat.
-le bois d'été ou bois final est constitué de petites trachéides aux parois épaisses. Plus dense, lisse et compact, il est de couleur plus foncée.
Il est donc possible de compter et de mesurer la largeur des cernes grâce à la différence de couleur entre le bois final de l'année N et le bois initial de l'année N+1. Cette séparation correspond à l'arrêt de croissance dès la fin de la saison de végétation et durant la période hivernale.
Les rondelles sont scannées à l’aide du logiciel Mac Dendro V3.2. L’image du scan est ensuite affichée à l’écran. A l'aide de la souris, l'opérateur clique au centre de la moelle et le logiciel Mac Dendro V3.2 repère automatiquement les limites des cernes le long des rayons grâce à la différence de contraste. A l'aide de la souris, l'opérateur peut annuler une limite de cerne mal placée et la placer manuellement au bon endroit, Le logiciel calcule automatiquement en millimètre la largeur de chaque cerne avec une précision qui dépend de la résolution du scannage (0,03 mm dans notre cas). Par la suite il sera donc possible de déduire la longueur totale du rayon, et la valeur d'un rayon à une année donnée en cumulant les largeurs des cernes formés au cours des années antérieures.
Cependant pour les rondelles de faible diamètre les derniers cernes ne sont pas toujours visibles sur l'écran de l'ordinateur du fait d'une résolution insuffisante. On utilise alors dans ce cas une loupe binoculaire pour observer la rondelle à plus fort grossissement, après l'avoir rafraîchie avec un scalpel, et ainsi observer et placer correctement les cernes les plus fins sur l'écran de l'ordinateur.
Il arrive aussi que sur les arbres très dominés, on ne puisse faire la distinction entre le bois final de l'année N et le bois initial de l'année N+1 sur certains rayons. C'est à dire que sur ce rayon à l'année N+1 l'arbre n'a pas poussé. On a alors ce que
5
Nord
1
Ouest
2
4
6
7
8
Sud
Est
3
7 l'on appelle un cerne manquant, un cerne de valeur nulle. Il est toutefois important de comptabiliser ces cernes particuliers pour pouvoir réaliser l'inter-datation entre rayons d'une même rondelle et entre rondelles.
L'épaisseur d'écorce au bout de chaque rayon a été également mesurée selon la même technique
2.4. Mise en forme et prétraitement des données.
2.4.1. Vérification de la chronologie : inter-datation.
C'est une étape nécessaire faisant suite aux mesures afin de vérifier leur cohérence et la justesse de ces dernières. Pour vérifier l'effet de la méthode de mesure, les données terrain de l'année N doivent être comparées avec les données laboratoire de la même année. Il est donc important de vérifier sur chaque rondelle si le nombre de cernes trouvé est correct. Le nombre de cernes doit être le même pour l'ensemble des rayons d'une rondelle mais peut varier d'une rondelle à l'autre, En effet, même si les arbres ont le même âge, ils n’ont pas mis le même nombre d'années pour arriver à « hauteur d'homme ».
Les arbres réagissent sensiblement de la même façon aux variations inter annuelles du climat. Ce fait est à la base des études de dendrochronolgie et dendroclimatologie. L'inter-datation est l'outil permettant de contrôler s'il existe un comportement de croissance similaire pour l'ensemble des individus selon les années. Cela consiste à tracer pour chaque arbre, la largeur moyenne de chaque cerne, en fonction de l'année. Cette valeur moyenne étant calculée à partir des largeurs mesurées sur les 8 rayons. Il suffit ensuite de superposer toutes les courbes obtenues. La superposition des courbes permet la mise en évidence des années remarquables, c'est à dire les années où tous les arbres ont eu une bonne croissance ou au contraire les années défavorables à leur développement. Par extension elle indique si sur les rondelles le bon nombre de cernes a été trouvé. Lorsque les valeurs maximum d'un arbre sont décalées par rapport aux autres on peut alors imaginer qu'un cerne ou plusieurs ont été oubliés lors des mesures. Grâce à cela on peut éviter les erreurs dans les données en refaisant les mesures sur un arbre qui est en décalage par rapport aux autres.
Cependant, il arrive qu’il y ait, sur des arbres dominés de faible diamètre, des cernes manquants. Ce sont des cernes que l'on trouve sur certains rayons mais plus sur d'autres. Il peut arriver aussi qu'un cerne soit manquant sur tout le tour de la rondelle. Dans ce cas il est impossible de le repérer pendant les mesures mais il est révélé lors de l'analyse de données. Dans notre étude les cernes manquants n’ont pas pu être datés. Les rondelles concernées ont été exclu de l’analyse de données.
2.4.2. Les modèles d'épaisseur d'écorce et de retrait.
Les mesures terrain sont des circonférences sur écorce à l'état frais alors que les mesures laboratoire sont des mesures de rayons réalisées sous écorce à l'état sec.
8 Avant d'analyser les données, il faut les rendre comparables et corriger les données laboratoire en passant des données sous écorce à l'état sec aux données sur écorce à l'état frais. Pour cela deux modèles ont été mis en point,
Au laboratoire, nous mesurons l'épaisseur d'écorce de 2008 uniquement. Alors que sur le terrain cette épaisseur d'écorce est prise en compte annuellement et augmente avec l'âge et surtout le diamètre des arbres. Un modèle d’épaisseur d'écorce a alors été mis au point pour avoir une valeur approximative de l'écorce aux années antérieures à 2008, et donc avoir une estimation du rayon sur écorce ces mêmes années.
Il faut aussi tenir compte du retrait du à la perte d'eau entre l'abattage et la mesure au laboratoire Ce retrait radial et transversal du bois aboutit à une diminution de la circonférence des rondelles par rapport à leur circonférence sur pied. Le retrait résultant de ces deux composantes a été modélisé.
Les abréviations utilisées dans le texte suivant, sont définis dans la figure 3.
La première étape dans la constitution du modèle d'écorce est le calcul de la valeur moyenne d'écorce de 2008 et la valeur moyenne du dernier rayon de la même année. Une fois cela fait pour tous les arbres, on trace le nuage de points (wecm)=f(rdmlast) on obtient alors un point par arbre. De là est tracée une courbe de tendance sur ce nuage de points. Les paramètres a et b de cette courbe de tendance, de la forme : epg = a × rb sont alors sauvegardés. Ce modèle est général pour tous les arbres. Il semble plus intéressant de faire un modèle individuel pour chaque arbre, pour gagner en précision. Pour cela on fixe le paramètre b mais on modifie le paramètre a, en forçant la courbe de l'arbre à passer par la dernière valeur qui est connue : celle de 2008 (cf annexe1). La variable « ai » est alors définie par
ai = wecm / rdmlastb. Cette variable est différente pour chacun des arbres « i ».
On obtient ainsi un modèle par arbre, que l'on écrit de la manière suivante :
ep = a
i× r
bGrâce à cette formule, avec un paramètre ai différent pour chaque arbre et un
paramètre b commun à tous les arbres, on peut alors estimer une épaisseur d'écorce en fonction de la valeur du rayon de chaque année.
Pour l'élaboration du modèle de retrait, la première étape est la mesure de la circonférence des rondelles « sèches » (CS). En soustrayant cette valeur à celle de la dernière mesure sur terrain (CF), nous obtenons un retrait en millimètre (R). Pour avoir ce retrait en pourcentage (rcf) on divise alors la valeur de retrait par la valeur de la dernière mesure terrain. Comme avec le modèle d'écorce on trace alors le nuage de points : rcf= f (CS), avec sa courbe de tendance associée, sous la forme :
rcfg = p × CS + q
9 retrait prédites en fonction de la circonférence sèche. Comme avec le modèle d’épaisseur d'écorce, dans un souci de précision, on réalise par la suite un modèle de retrait par arbre. Pour cela, il suffit de modifier l'un des paramètres de la courbe de tendance. Ici on gardera la pente de la droite p mais en changeant le paramètre q, on forcera la droite du modèle individuelle à passer par la valeur connue en 2008, celle présente sur le nuage de point (cf annexe 2). On obtient alors qi = rcf2008 – CS2008 × p.
A partir de là le modèle individuelle utilisé dans cette étude s'écrit.
rcf = p × CS + q
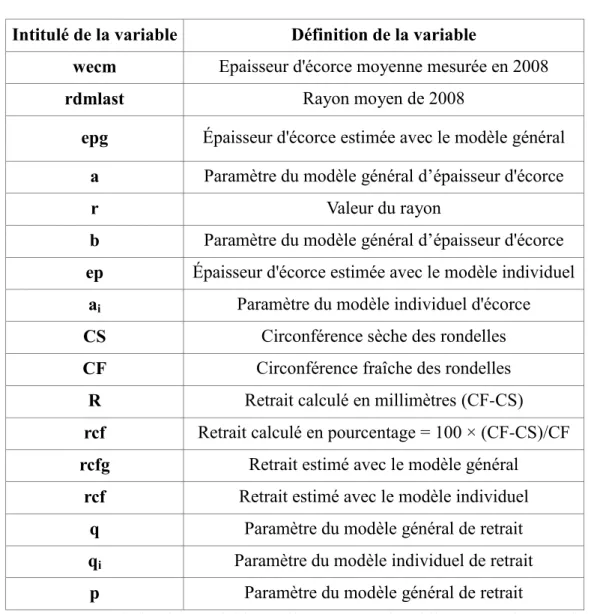
iIntitulé de la variable Définition de la variable
wecm Epaisseur d'écorce moyenne mesurée en 2008
rdmlast Rayon moyen de 2008
epg Épaisseur d'écorce estimée avec le modèle général a Paramètre du modèle général d’épaisseur d'écorce
r Valeur du rayon
b Paramètre du modèle général d’épaisseur d'écorce ep Épaisseur d'écorce estimée avec le modèle individuel
ai Paramètre du modèle individuel d'écorce
CS Circonférence sèche des rondelles
CF Circonférence fraîche des rondelles R Retrait calculé en millimètres (CF-CS)
rcf Retrait calculé en pourcentage = 100 × (CF-CS)/CF rcfg Retrait estimé avec le modèle général
rcf Retrait estimé avec le modèle individuel q Paramètre du modèle général de retrait qi Paramètre du modèle individuel de retrait
p Paramètre du modèle général de retrait
Figure 3 : Descriptif des variables utilisées pour l’établissement des modèles d’épaisseur d’écorce et de retrait.
2.4.3. Calcul des accroissements en surface terrière.
La surface terrière* d'un arbre notée g est la surface de la section transversale à hauteur d'homme de cet arbre. Le stage de master a étudié les accroissements en surface terrière qui est plus proche de l'accroissement en matière (biomasse) que l'accroissement en diamètre ou en circonférence,
*Surface terrière : Cet indice correspond pour un arbre donné : à la surface de sa section mesurée à 1.30 m du sol.
10 A partir des données terrain, la meilleure estimation de la surface terrière à partir de la mesure de la circonférence est donnée par la formule dans laquelle on suppose que la section de l'arbre est circulaire, ce qui n'est jamais rigoureusement exact ( Rondeux 1993):
Pour comparer les deux méthodes, il nous faut, à partir des rayons mesurés au laboratoire, estimer une surface terrière. Après application des deux modèles sur la valeur des 8 rayons, on en déduit la moyenne arithmétique, pour appliquer la formule mathématique suivante :
Les accroissements en surface terrière terrain et labo, qui sont les variables sur lesquelles porteront les analyses, s'en déduisent aisément par la différence entre la surface terrière de deux années successives
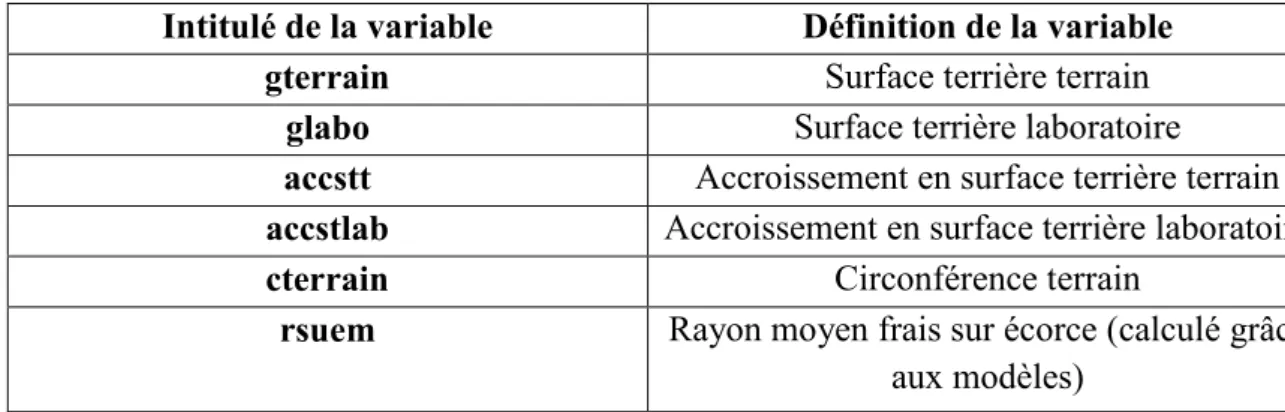
Intitulé de la variable Définition de la variable
gterrain Surface terrière terrain
glabo Surface terrière laboratoire
accstt Accroissement en surface terrière terrain accstlab Accroissement en surface terrière laboratoire
cterrain Circonférence terrain
rsuem Rayon moyen frais sur écorce (calculé grâce aux modèles)
Figure 4 : Descriptif des variables utilisées pour le calcul des surfaces terrières.
glabo=Pi × rsuem²
gterrain=cterrain²/4 × Pi
11
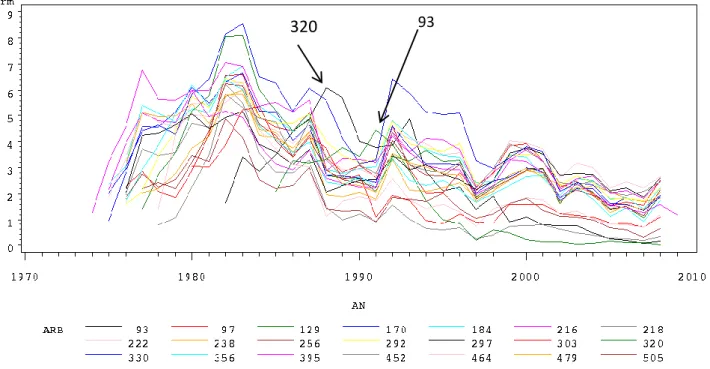
320 93
3. Résultats
3.1. L'inter-datation.
3.1.1. Comparaison des largeurs de cernes entre arbres.
Figure 5 : Évolution de la largeur moyenne des cernes à l’état sec en mm (rm), en fonction de l'année pour tous les arbres.
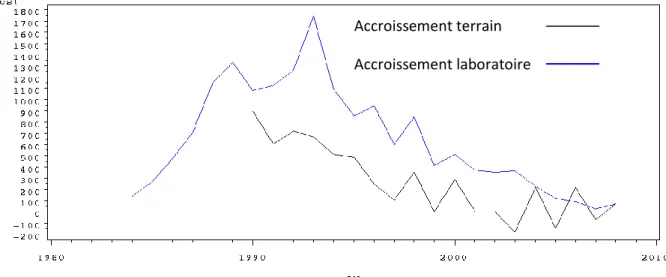
Figure 6 : Evolution des accroissements en surface terrière terrain et laboratoire en mm² de l’arbre 256.
Accroissement terrain Accroissement laboratoire Accroissement terrain Accroissement laboratoire
12 Accroissement terrain
Accroissement laboratoire
Figure 7 : Évolution des accroissements en surface terrière terrain et laboratoire en mm2, de l'arbre 93.
La figure 5 illustre l'évolution de la largeur moyenne des cernes en fonction des années pour tous les arbres mesurés. Elle montre que globalement les arbres présentent le même type d'évolution. Lorsqu’une année est favorable à la croissance, le cerne correspondant à cette année est plus important que ceux des années voisines moins favorables, cela se traduit par un pic sur la courbe. Et inversement pour les années défavorables qui se traduisent par un creux de la courbe. Il est alors possible de noter les années remarquables : 1992 et 1999 sont de bonnes années puisque elles font suites aux deux éclaircies, qui réduisent temporairement la compétition entre les arbres. Au contraire 1991 1997 et 2002 ont été des années défavorables pour les arbres ; 1991 et 1997 sont les années qui précédaient les éclaircies quand la compétition entre arbres était très importante. La mauvaise année 2002 est quant à elle certainement due à de mauvaises conditions climatiques.
Cependant deux arbres sont différents des autres le n°93 et le n°320 ont leurs années remarquables décalées.
3.1.2. Comparaison des accroissements en surface terrière laboratoire et terrain.
La figure 6 illustre l'évolution des deux types d'accroissements en surface terrière de l'arbre 256. L'arbre 256 a un comportement représentatif des arbres étudiés. Les deux courbes d'accroissement sont relativement en phase au fil des années avec des valeurs comparables : il n'y a pas de suspicion de cerne manquant.
Sur la figure 7, on observe un important décalage entre les courbes des différents types d'accroissements, pour l'arbre 93. Le même type de graphique est observable pour l'arbre 320. Ces écarts importants d'accroissement, ne peuvent être dus qu'à un oubli de cernes ou à des cernes manquants. Les vérifications à la loupe binoculaire ont permis d'établir qu'il y avait des cernes manquants sur toute la périphérie des rondelles. Il n’a pas été possible de les dater à cause des faibles largeurs des cernes voisins et donc de l’absence d’année remarquable repérable. Ces
13 Y=1.03742*X
R2=0.9931
deux arbres ont donc été écartés de l'analyse des données.
3.2. Comparaison individuelle entre les deux méthodes de mesure.
Durant l'analyse des résultats on teste l'hypothèse que l'accroissement en surface terrière terrain (accstt) est égal à l'accroissement en surface terrière estimé en laboratoire (accstlab). Donc que la droite du modèle accstt en fonction de accstlab est une droite d'équation : accstt = a × accstlab avec a=1 (cf figure 4). L'analyse portera donc sur ce modèle et ses résidus. Les résidus étant les différences entre valeurs estimées par le modèle et valeurs observées.
3.2.1. La régression.
Arbre R2 Estimation du paramètre (a) 97 0.9812 1.03991 129 0.9866 1.02900 170 0.9724 1.01299 184 0.9902 1.03475 216 0.9891 1.03630 218 0.8865 1.05068 222 0.9931 1.03742 238 0.9879 1.05061 256 0.9781 1.04673 292 0.9918 1.03275 297 0.9870 1.03106 303 0.9832 1.07670 330 0.9811 1.02366 356 0.9852 1.06184 395 0.9867 1.02136 452 0.9751 1.01732 464 0.9726 1.03098 479 0.9776 1.05301 505 0.9678 1.02471
Figure 8 : Résultats des régressions accstt = a × accstlab par arbre.
14 Le tableau de la figure 8 montre pour tous les arbres, une forte corrélation entre les accroissements terrain et les accroissements laboratoire, avec des R2 supérieurs à 0.96 hormis le 218 qui a un R2 de 0.89. Nos deux mesures sont fortement liées. Le tableau indique aussi un coefficient de pente, de la droite de régression toujours supérieur à 1 montrant que les accroissements terrain sont toujours supérieurs aux accroissements laboratoire.
3.2.2. Tests sur les résidus du modèle : accstt = f (accstlab).
Afin de confirmer statistiquement les observations graphiques. Un test de comparaison de moyenne fut réalisé afin de vérifier si la moyenne des résidus du modèle par arbre est significativement différente de zéro ou pas. Ceci dans le but de savoir si nos deux méthodes aboutissent à des résultats significativement différents ou non.
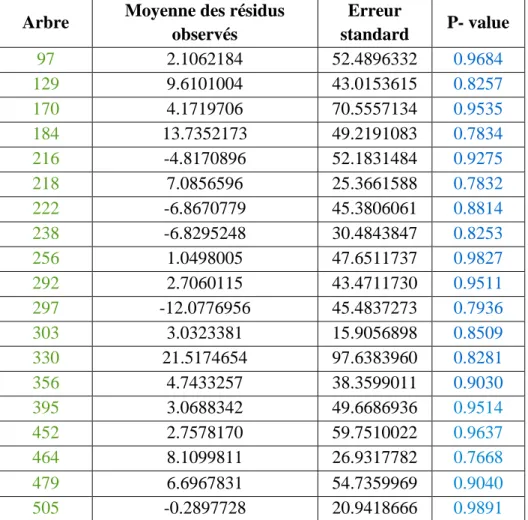
Figure 10 : Résultats sur les tests de comparaison de moyenne par arbre. Ho : la moyenne des résidus est égale à 0.
Arbre Moyenne des résidus observés Erreur standard P- value 97 2.1062184 52.4896332 0.9684 129 9.6101004 43.0153615 0.8257 170 4.1719706 70.5557134 0.9535 184 13.7352173 49.2191083 0.7834 216 -4.8170896 52.1831484 0.9275 218 7.0856596 25.3661588 0.7832 222 -6.8670779 45.3806061 0.8814 238 -6.8295248 30.4843847 0.8253 256 1.0498005 47.6511737 0.9827 292 2.7060115 43.4711730 0.9511 297 -12.0776956 45.4837273 0.7936 303 3.0323381 15.9056898 0.8509 330 21.5174654 97.6383960 0.8281 356 4.7433257 38.3599011 0.9030 395 3.0688342 49.6686936 0.9514 452 2.7578170 59.7510022 0.9637 464 8.1099811 26.9317782 0.7668 479 6.6967831 54.7359969 0.9040 505 -0.2897728 20.9418666 0.9891
15 Accroissement terrain
Accroissement laboratoire
3.3. Comparaison annuelle entre les deux méthodes de mesure.
3.3.1. Observations graphique des données.
Figure 11 : Évolution des deux types d'accroissements en surface terrière en fonction de l'année, pour l'arbre 452.
L'arbre 452 est représentatif de l'ensemble des observations faites sur les autres arbres. Les courbes d'accroissements obtenues par les deux mesures sont assez similaires jusqu'en 2002, après cette année-là, en revanche les courbes ne sont plus du tout en phase. Il est alors possible d'imaginer un effet lié à l'arrêt de la double mesure.
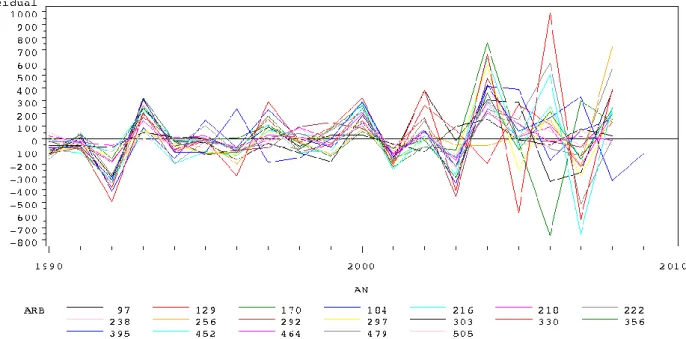
Figure 12 : Evolution des résidus du modèle accstt=f (accstlab) en fonction de l'année par arbre.
16 Figure 13 : Évolution de la moyenne des résidus du modèle accstt= f (accstlab) en fonction des années.
Figure 14: Évolution de la variance des résidus du modèle accstt= f (accstlab) en fonction des années.
La figure 12 semble indiquer une différence notable entre les résidus de chaque arbre après 2002. Durant les années avec la double mesure les arbres évoluent de manière semblable et sont plutôt en phase, alors qu'après 2002 les courbes des résidus sont plus anarchiques et n'évoluent plus de manière synchrone. L'évolution de la moyenne des résidus au cours des années présentées sur la figure 13 ne révèle pas d'effet de la double mesure, elle reste centrée sur 0. Au contraire de la
17 figure 14 où la brusque augmentation de la variance après 2002 est clairement notable. Il semble donc bien qu'il existe un effet lié à la double mesure.
3.3.2. Test sur les résidus du modèle accstt = f (accstlab) en fonction de l'année.
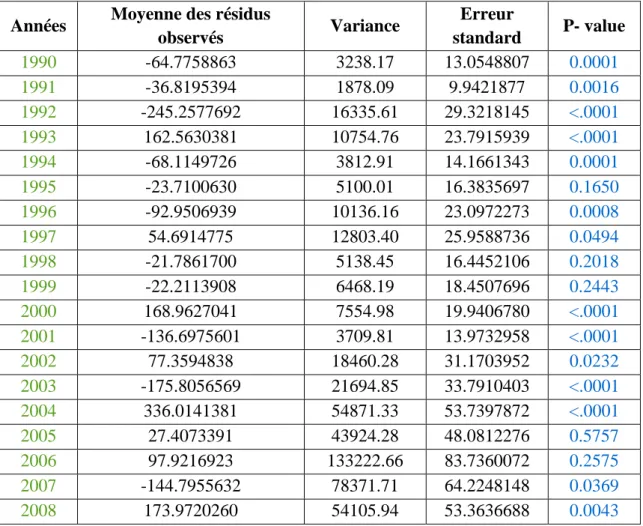
Années Moyenne des résidus
observés Variance Erreur standard P- value 1990 -64.7758863 3238.17 13.0548807 0.0001 1991 -36.8195394 1878.09 9.9421877 0.0016 1992 -245.2577692 16335.61 29.3218145 <.0001 1993 162.5630381 10754.76 23.7915939 <.0001 1994 -68.1149726 3812.91 14.1661343 0.0001 1995 -23.7100630 5100.01 16.3835697 0.1650 1996 -92.9506939 10136.16 23.0972273 0.0008 1997 54.6914775 12803.40 25.9588736 0.0494 1998 -21.7861700 5138.45 16.4452106 0.2018 1999 -22.2113908 6468.19 18.4507696 0.2443 2000 168.9627041 7554.98 19.9406780 <.0001 2001 -136.6975601 3709.81 13.9732958 <.0001 2002 77.3594838 18460.28 31.1703952 0.0232 2003 -175.8056569 21694.85 33.7910403 <.0001 2004 336.0141381 54871.33 53.7397872 <.0001 2005 27.4073391 43924.28 48.0812276 0.5757 2006 97.9216923 133222.66 83.7360072 0.2575 2007 -144.7955632 78371.71 64.2248148 0.0369 2008 173.9720260 54105.94 53.3636688 0.0043
Figure 15 : Résultats sur les tests de comparaison de moyenne par années. Ho : la moyenne des résidus du modèle est égale à 0.
Le tableau ci-dessus révèle des différences significatives entre les deux méthodes de mesure à certaines années mais pas à d'autre. Certaines années la moyenne des résidus du modèle est donc significativement différente de zéro, indiquant des résultats significativement différents selon la technique utilisée. Cependant on remarque des p-value inférieurs à 0.05 en amont et en aval de 2002, il semble donc qu'il y est bien un effet annuel, mais que celui si n'est pas uniquement lié à l'arrêt de la double mesure.
18 3.3.3 Coefficient de synchronisation (SR)
Le calcul de ce coefficient a pour objectif de vérifier mathématiquement les observations graphiques faites sur la figure 12 et de confirmer à l’aide d’indices reconnus dans le domaine de la dendrochronologie, l’effet de la double mesure. Le coefficient de synchronisation correspond au plus ou moins bon synchronisme de l’évolution des résidus du modèle accstt = f (accstlab). Les valeurs les plus proches de 1 expriment un synchronisme maximum : les résidus évoluent de manière similaire pour une année donnée. Les valeurs proches de 0 expriment en revanche un asynchronisme total de l’évolution des résidus. Ce coefficient est le rapport de la sensibilité moyenne (SMm), calculé avec la moyenne des résidus d’une année N, sur
la moyenne des sensibilités moyennes calculées pour chaque arbre (SMi)
(Lebourgeois, 2010). Notons qu’habituellement les sensibilités se calculent avec les largeurs de cernes, mais dans notre cas nous les calculerons avec les résidus du modèle accstt = f (accstlab) augmenté de 4000 pour travailler avec des valeurs positives.
SR= SMm/ SMi
Période de calcul 1990_2008 1990_2002 2002_2008
SR 0.78 0.86 0.72
Figure 16 : Résultat des calculs des coefficients de synchronisation pour différentes périodes.
La figure 16, confirme les observations graphiques et révèle une synchronisation des résidus après 2002 inférieur à celle avant 2002. L’effet de la double mesure est ainsi mis en avant de manière mathématique.
19
4. Discussion
4.1. Synthèse sur l'analyse des résultats. 4.1.1. L'inter-datation.
L'inter-datation a permis de mettre en relief des erreurs obtenues lors de l'acquisition des données. Tout d'abord avec la mise en évidence des années remarquables, deux arbres avaient un comportement différent des autres. Puis avec la juxtaposition des évolutions des deux types d'accroissements en surface terrière en fonction de l'année, l'existence de cernes manquants a pu être notée pour les arbres 93 et 320. Notons que ce sont les arbres avec les deux plus petites circonférences.
Ces deux arbres ont, durant toute leur existence, été dominés par les autres. On dit qu'ils étaient d'un faible statut social. Au début de leur croissance la compétition entre les arbres étant quasi inexistante, ces arbres ont pu se développer normalement. Cependant croissance et compétition sont indissolublement liées. La compétition résulte de la croissance des arbres, et en retour, elle impose des contraintes et des limites à cette croissance (Dreyfus 1988). La compétition pour l'eau la lumière et les nutriments s'est alors installée progressivement. Les arbres 93 et 320 ont souffert de cette compétition, malgré les éclaircies qui sont censées la diminuer. Cette domination a abouti à des problèmes de croissance ou du moins à une croissance moins importante que les autres arbres. C'est dans les dernières années de leur vie que ces arbres ont le plus souffert de la compétition, les arbres les entourant étant plus imposants et avec des besoins largement supérieurs. C'est donc durant cette période de leur vie, que les arbres 93 et 320 ont produit très peu, voire pas de bois. Le cerne est alors manquant. Cependant il arrive que ce cerne ne soit pas manquant sur tout le tour de la rondelle. Lorsque cela arrive, avec la loupe binoculaire il est possible de suivre le cerne sur tout le tour de la rondelle, pour repérer les endroits où il est présent et ceux où il disparaît, et ainsi le placer correctement avec le Mac Dendro. Cependant lorsqu’un cerne est manquant sur tout le tour, son année ne peut être déterminé avec exactitude, comme cela a été le cas pour les rondelles des arbres 93 et 320.
La figure 17 illustre bien ce phénomène de croissance nulle qui aboutit à un cerne manquant, on observe bien le bois final des années successives qui s'accolent les uns aux autres sans pouvoir distinguer une année d'une autre. Les comparaisons de mesures devant se faire précisément entre une année terrain N et l'année laboratoire N, il est logique que les arbres 93 et 320 aient été éliminés pour la suite des analyses.
20 Figure 17 : Différents types de croissance observables chez les arbres (Motta 1996).
4.1.2. Analyse individuelle.
L'objectif ici était de comparer les deux méthodes de mesure arbre par arbre pour comparer les deux techniques employées. Il ressort tout d'abord graphiquement qu'il existe une forte relation entre les valeurs terrain et les valeurs laboratoire. Par la suite les R2 élevés mettent en évidence une forte corrélation entre ces deux variables. Enfin les tests statistiques mettent en avant le fait qu'il n'existe aucune différence significative entre les deux méthodes de mesure pour un individu donné, au seuil de 5%. Il semblerait, donc que la méthode de mesure n'ai pas d'effet significatif sur l'estimation de l'accroissement en surface terrière.
Néanmoins il est possible de faire quelques remarques. Le tracé des droites de régressions sur les nuages de points accstt= f (accstlab), révèle pour tous les arbres un coefficient directeur supérieur à 1. Il existe donc une tendance à surestimer l'accroissement en surface terrière sur le terrain par rapport à l'accroissement en surface terrière laboratoire. Cette tendance à la supériorité s'explique du fait de la technique de mesure utilisée sur le terrain. Premièrement il est impossible de placer parfaitement le ruban perpendiculairement à l'axe de l'arbre, ceci provoque une surestimation de la circonférence, qui est peu plausible lors de mesure radiale. L'écorce n'étant pas forcément lisse sur toute la circonférence, surtout lorsque l'arbre avance dans l'âge, les rugosités du tronc peuvent elles aussi biaiser le placement du ruban et conduire à une surestimation de la mesure. La tension appliquée sur le ruban est également une source d'erreur pour la mesure de la circonférence, selon la tension appliquée la circonférence sera surestimée ou sous-estimée. Enfin, intervient aussi le problème d'exactitude et de netteté des graduations qui peuvent elles aussi mener à des erreurs de mesures (Massenet 2009). Le placement du ruban et les rugosités du tronc restent les responsables majoritaires dans l'erreur de mesure, d'où la surestimation constatée.
Notons aussi les valeurs des p-value, des tests de comparaison de moyenne, qui sont largement non significatifs. Le p-value le plus faible est de 0.7668 le plus
21 élevé de 0.9891. Grâce à ces résultats il est alors possible de dire que pour un individu donné son accroissement moyen en épaisseur n'est pas significativement différent, que l'on estime cet accroissement à partir de mesure annuelle de circonférence sur pied, ou bien avec des mesures radiales sur rondelle.
Après observation de l'effet de la méthode de mesure, sur l'estimation de la croissance en surface terrière au niveau de l'individu, il semble intéressant de regarder si on obtient des résultats similaires en prenant en compte tous les arbres en même temps mais par année. Ce type d'analyse va nous servir de la même manière que l'inter-datation. En regroupant les valeurs de plusieurs arbres nous pourrons mettre ou non en évidence un éventuel effet « année » sur la méthode de mesure. Un effet annuel qui ne pouvait être mis en évidence en analysant les résultats des individus de manière indépendante.
4.1.3. Analyse annuelle.
L'analyse annuelle portait sur les 19 arbres gardés pour l'analyse de données. Les premières conclusions sont graphiques. Les courbes d'accroissement en surface terrière des deux méthodes de mesure sont relativement synchrones jusqu'en 2002, à partir de 2003 les deux courbes ne sont plus en phase. La courbe d'évolution de l'accroissement terrain évolue avec de plus grande amplitude que celle des données laboratoires après 2002. De plus la figure 12 présente une évolution des résidus du modèle, moins synchrone et plus anarchique entre les arbres après 2002. Il semble donc bien qu'il existe un effet « double mesure ». Les figures 13 et 14 sont là pour valider cette hypothèse. La moyenne des résidus du modèle sur la figure 13 n'évolue pas de manière différente avant et après 2002. Elle reste centrée sur 0 tout au long des années. Au contraire la figure 14 est marquée par une nette augmentation de la variance des résidus après 2002, sachant que la variance est la somme des résidus du modèle au carré, les résidus du modèle après 2002 sont supérieurs à ceux avant 2003. Autrement dit les différences entre les deux mesures sont plus importantes après 2002. La figure 16 vérifie mathématiquement cette hypothèse à l’aide d’indices reconnus dans le domaine de la dendrochronologie. L'effet « double mesure » est mis en évidence.
La figure 15 montre que pour certaines années, il existe une différence significative entre les deux techniques de mesure. Mais aussi que dans ces années, certaines sont antérieur et d’autre sont postérieur, à 2002. Les p-value inférieurs à 0.0001 marquent clairement cette différence significative entre les deux méthodes en 1992, 2000, 2003, et 2004. Si l'effet « double mesure » existe bien ; ici est mis en évidence un autre effet annuel. Un effet annuel qui n'intervient pas de manière régulière. Or la mesure de la circonférence d'un arbre avec un ruban, est influencée par les conditions climatiques du jour de la mesure (communication personnelle Courbet 2011). Un taux d'humidité important dans l'air va faire gonfler l'écorce des arbres. Les jours de mesure où l'air était chargé en humidité la circonférence des arbres mesurés à donc été surestimée. Ceci rajouté à l'erreur de placement du ruban,
22 explique pourquoi pour certaines années les deux méthodes de mesure présentent une différence significative.
4.1.4. Lien entre les deux mesures
Au niveau arbre, il existe incontestablement un lien très fort entre les deux mesures. Au niveau individuel, les mesures sont fortement liées et aboutissent à des résultats similaires. Cela permet de confirmer la validité des études dendroécologiques faites par comparaison d'inventaires annuels et non de rayons sur rondelles, comme il est habituel de faire en dendroclimatologie et dendrochronologie. Les mesures de terrain sont toujours supérieures aux mesures de laboratoire montrant l'existence d'un biais qui s'interprète par le fait que le ruban est souvent mal placé ou pas assez tendu. On ne peut aussi complètement écarter le fait que de petits morceaux d'écorce ont pu être perdus entre le terrain et le laboratoire. Il existe toutefois un effet annuel sur cette relation entre les deux types de mesures. En effet les mesures sur le terrain sont dépendantes des conditions climatiques du jour de la mesure.
De plus la vérification de tous les arbres par une double mesure, semble bien avoir un impact sur la précision de la mesure.
4.2 Les points critiques de la méthode 4.2.1 Les mesures terrain
La mesure de la circonférence des arbres sur pied comporte quelques points faibles. Elle est dépendante de nombreux facteurs. Premièrement il est impossible de placer parfaitement le ruban parallèlement à 1,30 m, à la section de l'arbre. Ce fait rend une mesure exacte impossible et biaise la mesure en la surestimant.
La mesure terrain est également très dépendante des conditions climatiques environnantes : température, taux d'humidité. L'une des explications est la suivante : L’écorce étant de nature plus spongieuse que le bois, elle gonfle facilement par temps de pluie ou quand le taux d'humidité augmente. Cela menant à une surestimation de la circonférence sur pied de l'arbre. Les dimensions du bois varient aussi en fonction du taux d'humidité mais dans une moindre mesure.
Le gel peut être une autre explication. L'eau gelée occupant un volume supérieur par rapport à l'eau liquide pour une même quantité, lorsque les mesures sont faites à des températures où l'eau a gelé dans le bois, là aussi la circonférence de l'arbre est surestimée.
Pour finir, l'aspect physique de l'écorce peut être un problème pour la mesure. Lors de ses premières années de vie le cèdre a une écorce lisse, sa mesure au ruban ne pose alors pas de problème de ce côté-là. En revanche plus il grandit plus son écorce épaissit et devient rugueuse, avec des creux et des bosses, rendant le placement du ruban plus difficile, et l'erreur de mesure plus risquée.
23 protocole. Cette méthode permettait de limiter les erreurs de mesure au minimum sur l'ensemble des arbres.
4.2.2 Les mesures en laboratoire
Le fait que l'on ne puisse pas mettre en évidence l'existence des cernes manquants est le défaut principal des mesures en laboratoire. En effet nous l'avons vu précédemment, il arrive qu'un cerne soit manquant sur tout le tour de la rondelle. Dans notre cas cela est arrivé sur des arbres très dominés et dont les cernes visibles étaient déjà très minces. Dans ce cas la synchronisation avec les autres arbres s'est révélée impossible. Seule la confrontation avec les mesures réalisées sur le terrain a permis de détecter ces cernes manquants mais sans pouvoir les dater. La résolution parfois limitée du scannage et la mauvaise résolution de l'image sur l'écran sont aussi un problème. Lorsqu'une définition d'image n'est pas assez importante pour localiser un cerne fin, le cerne est mesuré à la binoculaire. Même si l'erreur qui fait suite à cette manipulation est très infime, elle existe quand même.
Ceci soulève donc un problème important, en effet la majorité des mesures de largeur de cernes se font sur carotte. Comment arriver à savoir s’il existe un cerne manquant sur ce rayon mesuré ? Comment alors déterminer avec exactitude l’âge de l’arbre carotté ? Sans compter la difficulté de repérer certains accroissements qui peuvent se traduire par la présence de seulement quelques cellules. Tout cela amène à s’interroger sur les mesures de largeurs de cernes effectuées sur carottes par l’INRA. Les mesures sur rondelles restent plus fiables mais sont destructives puisque elles obligent l’abattage de l’arbre.
4.2.3 Les modèles
Les modèles d'épaisseur d'écorce et de retrait permettent une estimation de ces deux valeurs mais ne représentent pas la réalité. Dans un souci de rendre comparable les deux types de mesures, (passage du sous écorce à l'état sec au sur écorce à l'état frais), ont été établis des modèles généraux, qui ont été par la suite adaptés à chaque arbre. Même si cette technique employée est la plus juste, elle ne peut donner avec précision les véritables valeurs de retrait et d'épaisseur d'écorce pour chaque année, mais seulement une approximation. Ces modèles sont cependant indissociables des mesures laboratoire, sans eux la comparaison avec les mesures terrain aurait été impossible.
Même si cette technique n'est pas complètement satisfaisante, la construction de modèles est la technique indispensable mais aussi la plus juste dans le cadre de notre étude. En revanche lors de travaux similaires futurs sur des espèces autres que le cèdre de l'atlas, ces deux modèles devront être modifiés en fonction des caractéristiques de l'espèce étudiée.
24 4.3. Perspectives d'améliorations
4.3.1. Mesures terrains
En ce qui concerne le placement du ruban, aucune amélioration ne pourra être faite en dehors de précautions renforcées au moment de la mesure. Mais même dans ce cas la mesure au ruban tend toujours à surestimer la circonférence de l'arbre, pour les raisons vues précédemment.
En revanche pour limiter l'effet annuel lié aux conditions climatiques du jour, il serait peut être intéressant de fixer certains critères météo le jour des mesures pour mesurer toujours dans des conditions semblables. Cependant cet objectif est très dur à mettre en place ; tout d'abord comment déterminer les critères météo ? De plus les jours de sortie sur le terrain sont fixés à l'avance, il est alors impossible de savoir à l'avance si les critères météo seront respectés le jour j. Sans compter que les jours précédents le jour de la mesure peuvent avoir un impact sur celle-ci. S'il a plu la veille des mesures, l'écorce sera gonflée entraînant une mesure faussée. Cette solution serait idéale mais difficilement réalisable.
La réalisation du double échantillonnage, est une pratique, à remettre en place dans les mesures terrain. Comme cela a été démontré plus haut, ses effets sont positifs et non négligeables pour une acquisition de données les plus justes possible.
4.3.2. Mesures en laboratoires
Pour parer à la non détection des cernes manquants en laboratoire, il n'existe qu'une seule méthode. Celle-ci est facile à mettre en place, mais est très coûteuse en temps et n'aurait pas pu être mise en place lors de ce stage. Cette technique consiste à prélever sur les arbres, d'autres rondelles à des hauteurs différentes jusqu'au sommet. Il est en effet rare qu'un cerne soit absent tout le long de la tige, il est en général visible dans la partie la plus vigoureuse du houppier En procédant ainsi, il est alors possible de suivre l'évolution et le devenir de chaque cerne au sein de l'arbre entier, en partant de la cime jusqu'à 1.30 m de hauteur. Ce suivi individuel permet de détecter la zone dans laquelle un cerne disparaît et ainsi déterminer son année. Cependant sur chaque arbre une seule rondelle a été prélevée à 1.30 m de hauteur, et même si d'autres rondelles avaient été prélevées à des hauteurs différentes, la trop courte durée du stage n'aurait pas permis une analyse approfondie de toutes les rondelles d'un ou plusieurs arbres.
25
Conclusion :
En conclusion de cette étude, il semble que les deux méthodes de mesure d’accroissement des arbres ne soit pas très différentes selon le point de vue où l’on se place. En se plaçant du point de vue de l’individu les techniques ne semblent pas conduire à des résultats significativement différents. En revanche sur le plan annuel la similitude des deux mesures reste dépendantes de nombreux facteurs extérieurs. Même si elles sont fortement liées, cette liaison reste fragile et met en jeu plusieurs éléments. Mais pour retenir l’essentiel il semble que des mesures annuelles en circonférence sont utilisables pour réaliser une étude de dendrochronologie. Cependant des mesures sur un grand nombre d’année sont nécessaire pour réaliser ce type d’étude, or cette technique de mesure n’est pas fréquentes, car difficile à mettre en place, d’où l’utilisation plus courante des mesures sur carotte.
Cette étude a également soulevé le problème des cernes manquants, des cernes manquants qui peuvent fausser les acquisitions de données de largeurs de cernes faites sur carotte. C’est cependant la technique la plus utilisée car facile à mettre en place et surtout non destructive. La mesure sur rondelle permet d’avoir une meilleure précision, mais elle oblige l’abattage de l’arbre, et les mesures sont plus longues à effectuer. D’où des mesures sur carottes généralisées dans le monde de la dendrochronologie.
Le jeu de données acquis et utilisé lors de cette étude aurait pu servir à une autre problématique, ayant pour objectif de comparer les mesures entre carottes, et rondelles. Il aurait été intéressant de comparer les résultats d’estimation de surfaces terrières et d’accroissements en surface terrières obtenus avec une carotte, simulée par un rayon, et avec une rondelle, réunissant huit rayons. Quels résultats avec deux, trois ou quatre carottes ? Etc.... Mais la durée du stage trop courte, ne nous a pas donné l’occasion de traiter ce sujet.
26
Bibliographie
Bréda, N., Badeau, V., Bosc, A., 2011. Climator, simulations d'impacts du changement climatique. Forêt entreprise. N°196 p.37
Dreyfus, P., 1988. Compétition et croissance dans de jeunes peuplements résineux aspects biologiques, écologique et sylvicoles. Thèse de Doctorant. p.1.
Duplat, P., Perrotte, G., 1981. Inventaire et estimation de l’accroissement des peuplements forestier.
§ 6.2.1. Le sondage à la tarière et l’accroissement courant en diamètre sur écorce. 101-102.
§ 6.5 Discussion sur les méthodes d’estimation de l’accroissement à l’aide de sondage à la tarrière. 115-123.
Annexe X : sondage à la tarière et estimation sans biais de l’accroissement radial d’un arbre. 263-269.
AnnexeXI : variabilité des accroissements radiaux et dissymétrie de la croissance des arbres.271-279.
ONF Section Technique.
Girerd, B., 1991. La flore du département du Vaucluse. p.116.
Hennon, J-M., Mourisset, D., 2006. Comment mesurer la grosseur d'un arbre ? Centre Régional de la Propriété Forestière d'Auvergne. Bulletin semestriel. Fiche technique n°38.
Khanfouci, M S., 2005. Contribution à l'étude de la fructification de la régénération du Cèdre de l'Atlas (Cedrus atlantica M.) dans le massif de Belezma. Université hadj lakhdar-batna. Mémoire pour l'obtention du diplôme, magister en science forestière. p.7.
Lebourgeois, F., 2010. Principes et méthodes de la dendrochronologie. Equipe
Ecologie Forestière ENGREF. p.22.
Motta, R., 1996. Metodi e problemi nella determinazione dell’età di alberi viventi in studi ecologici o dinamica forestale. Linea ecologica, n°1, p.2-9.
Massenet, J-Y., 2005. Chapitre I : Grosseur des arbres. Cour de lycée forestier. p.7. Pardé, J., Bouchon, J., 1988. Dendrométrie. 2éme édition. § 457 et 458 252-255.
Riou-Nivert Ph., 2007. Climat propice pour le Cèdre. Forêt entreprise. N°174 p.12-13.
27 ANNEXES
Annexe 1 : Modèle individuel de l’estimation de l’épaisseur d’écorce.
28 Annexe 3 : Coupe transversale du bois de Cedrus atlantica et cernes annuels de croissance.
Petites trachéides Grosses
trachéides
Bois initial Bois final
29
Résumé :
Dans le cadre du changement climatique le lien entre climat et accroissement des arbres fait l'objet de nombreuses études, et cela dans le but de préparer nos forêts à cette perturbation du climat. Deux techniques sont utilisées pour mesurer l'accroissement des arbres : La première consiste à mesurer la circonférence des arbres chaque année et la seconde à mesurer les largeurs de cernes sur carottes ou rondelles. L'objectif de ce stage est méthodologique. Il s'agit de s'assurer que quel que soit la technique de mesure nous arrivons à des résultats semblables et vérifier s’il existe un biais entre les deux techniques. Cette étude porte sur 21 cèdres de l’Atlas prélevés dans la forêt domaniale du Ventouret, sur un versant Sud du Mont Ventoux.
Nous nous sommes dans un premier temps intéressés à des comparaisons de mesures individuelles, c'est à dire arbre par arbre. L’analyse a montré qu’il n’existe aucune différence significative entre les deux techniques. En revanche il existe des différences significatives entre les deux méthodes de mesure selon les années. De nombreux facteurs, tel que les conditions climatiques du jour des mesures sur le terrain, mais aussi les choix fait dans le protocole de mesure expliquent ces différences.
De plus a été mis en évidence une faille dans le protocole de mesure de largeur de cernes sur carottes ou rondelles pouvant tromper les études de dendrochronologie.
Les résultats présentés dans ce rapport ayant une valeur générique valable pour toutes les espèces d’arbres, ces défaillances peuvent amener l’INRA à se poser des questions sur la justesse de leurs techniques de mesure d’accroissement des arbres.
Mot clés : cèdre / cerne / technique de mesure / surface terrière / dendrochronologie
Abstract
In the context of climate change the link between climate and tree growth has been investigated in a number of studies, specifically with a view to promoting forest adaptation to climate perturbation. Two techniques are available to measure tree growth: the first is measuring the circumferences of the trees every year, and the second is measuring ring width on tree cores or slices. The objective of this study is methodological; to discover whether the same results are achieved with both tree growth measurement techniques and whether there is bias in either of the techniques.
30 For this study we used 21 Atlas cedars (Cedrus atlantica) in the forest of Ventouret located on the southern slope of Mont Ventoux.
We compared the results of the two tree growth measurement techniques for each tree. Overall the analyses do not reveal any significant difference between the two techniques. However there are differences between the results from the two techniques at the scale of individual years. Several factors, including the climatic conditions on the day of measurement and the measurement protocol used may explain these differences.
In addition we suggest that there are potential failings within the wood core ring measurement protocol itself. This failings which can falsify dendrochronologie studies.
We suggest that the results presented here are relevant for a wide variety, tree species. Therefore we suggest that INRA take into account these potential pitfalls in application of tree ring measurement techniques.
Key words: cedar / ring / technique of measure / basal area / dendrochronology